










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Researchers in Japan have discovered that the Plasmodium parasites responsible for malaria rely on a human liver cell protein for their development into a form capable of infecting red blood cells and causing disease.
The study, which will be published June 12 in the Journal of Experimental Medicine, suggests that targeting this human protein, known as CXCR4, could be a way to block the parasite’s life cycle and prevent the development of malaria.

According to the World Health Organization, there were an estimated 219 million cases of malaria in 2017, resulting in the deaths of approximately 435,000 people.
Infected mosquitoes transmit Plasmodium parasites to humans in the form of rod-shaped sporozoites that travel to the liver and invade liver cells (hepatocytes).
Once inside these cells, the Plasmodium sporozoites develop into spherical exoerythrocytic forms (EEFs) that eventually give rise to thousands of merozoites capable of spreading into red blood cells and causing malaria.
“It seems likely that the transformation of Plasmodium sporozoites into EEFs is tightly controlled so that it only occurs in hepatocytes and not at earlier stages of the parasite’s life cycle,” says Masahiro Yamamoto, a professor at the Research Institute for Microbial Diseases of Osaka University.
“However, we know very little about the host factors that regulate the differentiation of sporozoites in infected hepatocytes.”
In the new study, Yamamoto and colleagues discovered that a hepatocyte protein called CXCR4 helps Plasmodium sporozoites transform into EEFs.
Depleting this protein from human liver cells reduced the ability of sporozoites to develop into EEFs.
Moreover, mice pretreated with a drug that inhibits CXCR4 were resistant to malaria, showing reduced levels of parasites in the blood and significantly higher survival rates following Plasmodium infection.
CXCR4 structure
The CXCR4 structure consists of 352 amino acid residues comprising an N-terminal domain, seven TM domains, three extra-cellular loops (ECL), three intra-cellular loops (ICL) and a C-terminal domain [11]. Crump et al proposed a two-site theory for the binding of the natural ligand, SDF-1 with CXCR4 [12].
First, the RFFESH loop (site 1) of SDF-1 interacts with the N-terminal domain of CXCR4; then the N-terminal region (site 2) of SDF-1 binds to the receptor groove comprising the TM helices and the extra-cellular loops.
There have been several crystal structures of the CXCR4 protein published. PDB identification codes 3ODU and 3OE0 describe the crystal structures of the TM regions of CXCR4 co-crystallized with a small-molecule inhibitor IT1t and a cyclic peptide inhibitor CVX15, respectively (Figure 2) [11].
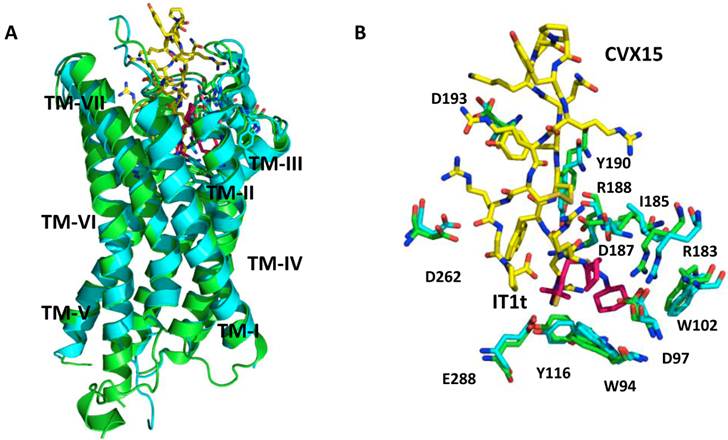
Both structures are CXCR4 homodimers, with IT1t or CVX15 located in the ligand-binding cavity that comprises the N-terminal, ECL2, ECL3 and TM domains.
It is important to note that the binding cavity of CXCR4 is larger and closer to the extra-cellular surface compared to other GPCRs. IT1t binds in only a portion of the cavity, interacts with TMs I, II, III and VII, while, peptide CVX15 occupies the complete binding cavity and makes contact with all TMs.
In addition, CVX15 binding causes conformational changes in the binding cavity, especially at the N-terminus and to some extent the extracellular portions of TMs V, VI and VII [11], whereas IT1t induces no significant conformational changes (Figure 2B).
CXCR4 and CCR5 are the two major co-receptors for HIV entry into its target cells in the human immune system and play important physiological roles in viral infection (Figure 3) [13, 14].
In a multi-step process, HIV enters the target cells by binding to the host surface receptor CD4 and a co-receptor, either CCR5 or CXCR4 [13].
As the initiation step, viral glycoprotein gp120 interacts with CD4, which in turn triggers the binding of gp120’s V3 loop to the N-terminus, ECL2, ECL3 and the ligand binding cavity of CXCR4 [11].
These interactions lead to a conformational change in the viral TM protein gp41, causing a pH-dependent fusion of the viral and the host cell membranes and the delivery of the viral payload [15–18].
In early stages of HIV infection, HIV predominantly uses the CCR5 co-receptor, whereas during the disease progression HIV uses either CXCR4 alone or in combination with CCR5 in about 50% of the infected individuals [18, 19].
Use of CXCR4 as a co-receptor is associated with a marked drop in CD4+ T-cell counts [19]. Unfortunately, individuals infected by CXCR4 utilizing strains experience a faster rate of disease progression [20, 21].
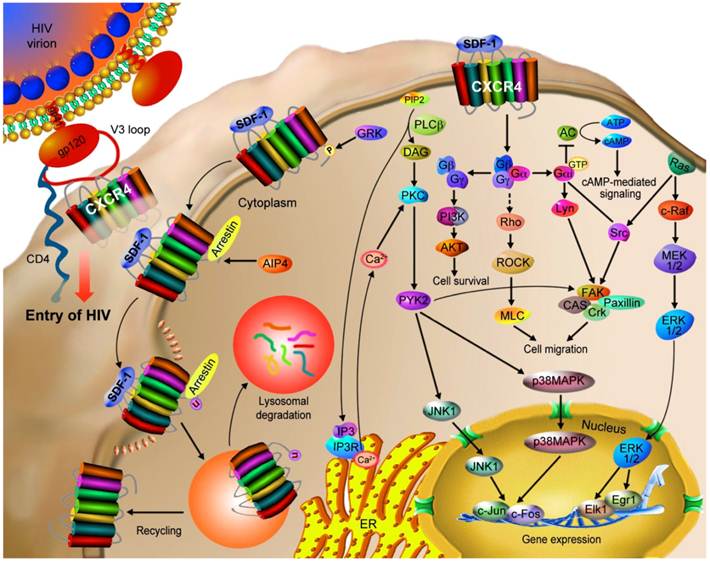
CXCR4 mediates HIV infection and cancer progression. CXCR4 is a co-receptor used along with CD4 by HIV-1 strains for infecting T cells. The binding of gp120 to CD4 induces a conformational change of gp120, allowing it to interact with CXCR4’s N-terminal, ECL2 and ECL3 domains as well as the ligand binding cavity through the V3 loop of gp120. These interactions trigger a conformational change in gp41, causing a pH-dependent fusion of the viral and the host cell membranes and thus the delivery of the viral payload. CXCR4 is also involved in the progression of tumor (hematopoietic and solid) via the interaction with SDF-1. The SDF-1/CXCR4 interaction stimulates tyrosine phosphorylation of CXCR4, followed by the activation of multiple G protein-dependent signaling pathways, which may be different among cell types. The major potential pathways and their cellular outcomes are described in this figure. Activated CXCR4 recruits arrestin 2/3, which mediate receptor internalization. CXCR4 may then be recycled to the cell surface or ubiquitinated by AIP4 that results in the lysosomal degradation of CXCR4.
Yamamoto and colleagues also identified a cell signaling pathway that causes hepatocytes to produce more CXCR4 in response to Plasmodium infection and determined that the protein aids the parasite’s development by raising the levels of calcium inside the cells.
“Our study reveals that CXCR4 blockade inhibits Plasmodium sporozoite transformation in hepatocytes,” Yamamoto says.

“Most anti-malaria drugs targeting Plasmodium-derived molecules eventually lead to drug resistance in these parasites.
However, inhibitors targeting human proteins such as CXCR4 might avoid this problem and could be used prophylactically to prevent the development of malaria.
Moreover, the CXCR4 inhibitor used in this study is already widely used in humans undergoing treatment for blood cancers, which could accelerate its repurposing as a new way of combating malaria.”
More information: Bando et al. 2019. J. Exp. Med. jem.rupress.org/cgi/doi/10.1084/jem.20182227
Journal information: Journal of Experimental Medicine



{kind=link}
[…] Malaria Plasmodium parasites can be stop targeting CXCR4 protein […]