











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Glioblastomas, the deadliest type of brain tumor in adults, attract “turncoats.”
These are macrophages, a type of immune cell, which promotes tumor progression and mask tumors from the immune system’s scrutiny.
To better understand the cells that brain tumors recruit, scientists led by Dolores Hambardzumyan, PhD, developed advanced imaging techniques to visualize macrophages found in the brain tumors of living mice.
The results were published Monday, June 24 in PNAS.
The researchers demonstrated that depending on where these macrophages originated, their appearance and behavior vary significantly.
Hambardzumyan and her colleagues say their results provide support for a strategy that would block macrophage infiltration into brain tumors as part of cancer treatment.
“This research provides the scientific community with a novel mouse model to study the populations of glioblastoma-associated macrophages both separately and together,” Hambardzumyan says.
“Our results also demonstrate the efficacy of blocking macrophage infiltration and calls for the use of combination therapies to enhance patient survival.”
Hambardzumyan is an associate professor of pediatrics at Winship Cancer Institute of Emory University and Aflac Cancer and Blood Disorders Center at Emory and Children’s Healthcare of Atlanta.
Tumor-associated macrophages come from two distinct sources.
They can come from the peripheral blood supply, or they can be microglia: cells that are already resident within brain tissue.
The Hambardzumyan lab has previously demonstrated macrophages coming from the blood are much more prevalent in number and aggressive in behavior, compared to macrophages coming from the brain.
“In order to develop more effective immunotherapies to treat glioblastoma, we must first understand the properties of each of these macrophage populations individually rather than treating them as one homogenous entity,” says Zhihong (“Z”) Chen, PhD, a postdoctoral instructor and the co-first author of the paper.
The researchers developed a mouse model which allows them to observe infiltrating macrophages in real time inside the tumor.
The mice are genetically modified (also, some received bone marrow transplants) so that only particular groups of macrophage cells produce fluorescent proteins, making peripheral blood-derived macrophages or brain microglia visible under a microscope.
The team installed a clear imaging window made of thin glass over the brain tumor and utilized advanced techniques including two-photon imaging to observe macrophages over time as they move through and interact with the tumor.
They were able to image cells as deep as 500 micrometers in the tumor and up to several hours at a time.
The researchers found clear morphological and behavioral differences between macrophages coming from the blood and brain resident macrophages.
They showed blood-derived macrophages are smaller, more motile cells that zip around the tumor along blood vessels.
On the contrary, they found brain-resident macrophages to be large, highly branched, and immobile cells that have minimal migratory behavior.
To test the efficacy of blocking the infiltration of the blood-derived macrophages, they treated mice with a drug (anti-VEGF antibody or bevacizumab) that normalizes the irregular vasculature of glioblastomas.
This drug has been used in the clinic to treat glioblastoma and improve quality of life in patients.
The research team observed fewer macrophages in the tumors that were treated, and they observed an increase in survival in these mice.
They also observed a morphological switch in the macrophages, indicating that not only can the infiltration of these cells be inhibited but their state of differentiation may also be affected.

Brain tumor-associated microglia (left, green) and blood-derived macrophages (right, cyan) appear different when visualized by multiphoton microscopy. The microglia appear to be bushy and seaweed-like, exhibiting minimal migratory behavior, while the blood-derived macrophages are smaller and rounder, and zip around the tumor along blood vessels, shown in red. The image is credited to Zhihong Chen.
Despite aggressive therapeutic strategies, the average survival for patients with glioblastoma is only 12 months.
One reason for this dismal outcome is that current therapeutic strategies fail to effectively target properties of other cell types within the tumor, such as tumor-associated macrophages.
The Hambardzumyan lab is now actively seeking to find new therapeutic approaches, such as blocking macrophage infiltration in combination with immune checkpoint inhibitors to treat these deadly brain tumors.
Myeloid cells form a vast and heterogeneous group of cells that play a major role in shaping the tumor microenvironment (TME).
Tumor associated macrophages (TAM) represent the most abundant myeloid subset across multiple cancer types, and they generally correlate with poor outcomes. Dendritic cells (DC) in tumors represent a less abundant subset, and contradictory results surround their association with tumor prognosis. DC are classified into subpopulations exhibiting different specificity for priming T-cells (1, 2).
Macrophages and DC subsets display a strong overlap of phenotypic markers, adding a high level of complexity to accurately identify them. So far, TAM have been considered to arise primarily from monocyte cells. Recent discoveries regarding tissue macrophage ontogeny challenge this assumption (3).
Different tissues of origin are likely to contribute in part to how TAM heterogeneity arises (4–6).
Flow cytometry allows qualitative and quantitative characterization of these cells but does not preserve their in situ localization to study native cell-cell interactions. In contrast, intra-vital imaging at cellular-scale resolution offers the ability to study cell migration and interactions in living tissue in real-time.
It is tempting to consider “truth” what is visible to the eye, and thus direct visualization of cell interactions tends to provide more confidence in the interpretation of a biological process.
A main hurdle of this approach rests on the accurate tracking of these cells since the number of available markers are more limited than for flow cytometry and many markers overlap between monocytes, macrophages, DC and even neutrophils, potentially leading to misinterpretations.
Moreover, one must keep in mind that imaging experiments usually focus on specific cell subsets, avoiding the potential contribution of the “unseen.” Herein, we review how fluorescent imaging, and more specifically in situ live imaging, has contributed to the characterization of TAM and tumor-DC.
We discuss limitations of the most common models used for the discrimination and tracking of these different subsets, and we present some perspectives derived from the combination of different fluorescent reporter mouse strains used to unveil microanatomical niches of myeloid subsets in tumors.
Functional Imaging of Tumor-Associated Myeloid Cells
Microscopy studies represent a necessary approach to truly comprehend the relationship between cells in their physiological environment (7).
Beyond the simple identification of cell distribution across the tissue provided by histological analysis, the development of live imaging in situ has generated fundamental insights in cellular functions and is termed “functional imaging.”
Here, we mention studies based on monocyte and TAM imaging to highlight how this approach has contributed to our knowledge of their function within tumors.
Functional Imaging of Tumor-Associated Myeloid Cell Dynamics and Interactions With Tumor Cells
Intra-vital imaging of TAM has helped to identify their role in tumor invasiveness and metastasis (8, 9).
Direct visualization of fluorescent macrophages and tumor cell lines has revealed CSF1 and EGF-dependent chemotaxis, respectively, (10) and has led to the elaboration of a tumor cell/macrophage cross-talk model (7, 11).
In vitro imaging is an important complementary approach to study the molecular pathways involved in this model.
Beyond paracrine loops, the combination of in vitro and in vivo imaging has provided evidence that physical contacts between macrophages and tumor cells correlate with invadopodium formation through the induction of RhoA activity on tumor cells (12).
The strength of real time imaging is elegantly illustrated by the work of Harney et al showing that the role of Tie2+ perivascular macrophages in this intravasation process is transient and mainly occurs in highly defined microanatomical niches termed “Tumor Microenvironment of Metastasis” (TMEM) (13).
Another study has found that macrophages orchestrating early dissemination in breast cancer are CD206Hi and Tie2+ and migrate toward tumor cells through CCL2 production by the latter (14).
Macrophages have also been involved in the “streaming cell movement” of tumor cells, defined as the migration of multiple cells in a single file pattern (15).
Directional streaming toward the endothelium results from CXCR4 upregulation on TAM and CXCL12 secretion by peripheral fibroblasts (16).
Cocultures in 3D-matrices have provided the subcellular resolution to identify a macrophage/tumor cell communication mechanism involving the formation of tunneling nanotubes between the two cell types that is required to induce this directional cell streaming (17).
This heterotypic interaction might favor the switch from a mesenchymal migration mode of tumor cells toward an MMP-independent ameboid-like migration as observed in spheroid culture (18).
Cytoplasmic exchange between macrophages and tumor cells has been confirmed in vivo in zebrafish (19).
In vivo visualization of migratory activity of TAM, tumor-DC and neutrophils has been observed using differentially ingested dextran particles or differential staining by intravascular injection of fluorescent antibodies in MMTV-PyMT/cfms-EGFP+ mice. Sessile cells exhibited strong endocytosis and MMP activity, however TAM and tumor-DC could not be disciminated based on the tested labeling combination (20).
Similar labeling approaches have unveiled that migratory capacities of myeloid cells in mammary cancer were less sensitive to hypoxia than regulatory T-cells (21).
Considering macrophage ontogeny and tissue specification has raised the question of their differential function in pathological contexts, particularly in cancer development. Although microglial cells have been considered as the primary TAM subset in brain tumors, it is commonly held that the majority of TAM among many other tumors are monocyte-derived (MoD-TAM) (22).
Evidence is recently accumulating that tissue-resident macrophages represent a distinct functional subset from MoD-TAM in other cancer types (16, 23, 24).
While resident macrophages were associated with ECM production, recruited macrophages were more involved in the modulation of the adaptive immune response (24), in addition to matrix remodeling and tumor cell clearance following chemotherapeutic treatment (16).
So far, very little information on the role of tissue-resident macrophages in solid tumors is available from imaging studies.
The reporter model used in our recent study has been an interesting option for simultaneous tracking of macrophages of different origins in lung tumors (16).
MoD-TAM and monocytes tended to accumulate in the periphery of advanced lung tumor nodules and displayed higher displacements than their resident counterparts (16).
Their increased migratory behavior also fits with the observation of streaming TAM recruited in a CCR2-dependent manner (25).
Accordingly, CCR2-dependent recruited TAM in lung tumors have been associated with remodeling activity and higher tumor cell dissemination (16).
So far, modulating the CCL2/CCR2 axis appears useful in identifying the monocytic origin of TAM. Nevertheless, while the accumulation of tissue resident macrophages has been shown to be CCR2-independent in lung tumors (16), this subset binds CCL2, suggesting that they might respond to a local CCL2 gradient.
One should consider that targeting the CCR2 axis may directly or indirectly affect recruited as well as resident TAM.
Resident TAM do not necessarly have an embryonic origin but could also arise from local proliferation of MoD-resident macrophages that have progressively colonized the tissue at steady state as observed in several tissues (26).
Fate mapping models to track embryonic-derived macrophages by imaging are necessary to determine whether resident TAM are of embryonic origin.
Functional Imaging of TAM Role in Metastatic Seeding
With the opportunity to track single cells in real time, live imaging has greatly improved our knowledge on the early events of metastatic seeding, in particular through the development of in vivo lung imaging (27).
Patrolling monocytes have been reported to rapidly engulf tumor material in lung capillaries reducing metastasis development (28).
This patrolling activity has also been efficiently monitored using a peritoneal window in colorectal tumors treated with anti-VEGFR2 therapy, highlighting a protumoral activity through neutrophil recruitment (29).
Patrolling monocytes do not appear to be the only myeloid cells involved in this process. Rather, a series of sequential waves involving different myeloid subsets are able to uptake tumor material in the lung (30).
CCL2-dependent monocyte recruitment has been strongly implicated in metastatic seeding by experiments utilizing CCL2 blockade or global macrophage depletion (31, 32).
The relative roles of interstitial lung macrophages and monocyte-derived cells on this early process remain unclear.
Functional Imaging of TAM and Tumor-DC Interactions With Lymphocytes
Live imaging has also contributed to identifying direct interactions of myeloid cells with T-cells in the TME. Trapping of antigen specific T-cells by myeloid cells in sustained and non-productive interactions has been proposed to favor immunosuppression (33, 34).
Macrophage depletion has been associated with increased CD8 T-cell infiltration and improved response to anti-PD-1 “checkpoint” immunotherapy (35).
Macrophage/Treg interactions after radiotherapy have also been visualized in a model of head and neck cancer. TNF-mediated cross talk between the two subsets is a proposed mechanism responsible for how an immunosuppressive environment dampens therapeutic efficacy (36).
While the vast majority of tumor-infiltrating T-cells seem to be in contact with TAM correlating with poor ability to induce effector functions, Broz et al. have identified a sparse subset of tumor-DC with strong immunostimulatory capacities (2).
Recruitment of this subset via NK cell crosstalk mediated by FLT3 ligand and resulting physical interactions defines a positive prognostic factor for anti-PD-1 therapy in melanoma patients (37). Overall, this supports the idea that TAM are usually associated with immune suppressive activity while tumor-DC are more immunostimulatory (38).
Overall, monitoring myeloid cell dynamics, morphology, local distribution in specific TMEM, and interactions with other partners of the TME has unveiled many of their key biological mechanisms. However, the capacity to accurately identify specific myeloid subsets by imaging can be limiting.
Tracking Myeloid Cells in Tumors
Specific identification of myeloid cells by imaging is challenging because of their heterogeneity, plasticity, and overlapping markers.
In vivo antibody injection represents an interesting alternative for cell identification, but there are multiple limitations of this approach.
Efficient cell staining is limited by tissue penetration of antibodies, and the persistence of the staining is low due to degradation and recycling activities in living tissues.
Finally, the impact of multiple in vivo antibody staining on cell dynamics and function cannot be neglected, and findings regarding cell behavior should be interpreted with caution.
Fluorescent reporter mice are, thus far, the best option to overcome these limitations. However, the lack of cell-specifc labeling ability still presents a challenge. Promoter-driven fluorescent protein (FP) production is never restricted to a specific subset. Moreover, it is not recommended to associate reporter expression with endogenous protein expression.
Therefore, a careful phenotypic characterization of each model using flow cytometry is required to adequately define the imaged cell populations.
Many transgenic mice (listed below) have been developed with various fluorescent reporters to attempt to discriminate specific myeloid populations.
The development of a Csf1r-EGFP transgene (MacGreen) has confirmed that this receptor is expressed in monocytes, tissue-resident macrophages and some populations of DC, such as the Langerhans cells; yet is also present in trophoblasts and granulocytes (39, 40).
The deletion of a conserved distal element from the Csf1r promoter on the ΔCSF1R-ECFP reporter (MacBlue) mouse ablated expression in trophoblasts and reduced expression in granulocytes (41).
Reporter gene expression is maintained in alveolar macrophages, microglia, and Langerhans cells, however it is ablated in most resident macrophage populations including osteoclasts (42), Kupffer cells (43), and lung interstital macrophages (44).
Hawley et al. created a Csf1r-mApple mouse (MacApple) with the same pattern of expression as MacGreen mice (45). Crossing MacApple with MacBlue mice results in specific patterns of fluorescent expression among monocytes and macrophages as observed in the lung and the brain.
The authors propose that ECFP expression may be present in cells relying more on IL-34 or CSF2 while ECFP− mApple+ macrophages would depend more on CSF1 for their homeostasis (45, 46). The regulation of CSF1R expression requires further investigation.
The Cx3cr1EGFP reporter mouse (47) is commonly used to monitor patrolling monocytes (29, 48–50) and tissue macrophages (51), but this reporter is also expressed by subsets of NK cells and dendritic cells as well as epidermal T-cells harboring a dendritic-like morphology.
EGFP upregulation on subsets of T-cells has been also reported during viral infection (52). Whether tumor-infiltrating T-cells upregulate CX3CR1 must be investigated when using this strain as they can represent an important confounding subset when imaging the TME.
We have developed an additional dimension of resolution using the combination of MacBlue x Cx3cr1EGFP x MacApple reporter mice.
This strain provides an improved display of the myeloid compartement heterogeneity in lung tumors, allowing the visualization of recruited, resident interstitial, and alveolar macrophages as well as neutrophils based on differential expression of the fluorescent reporters (Figure 1A).
This further highlights microanatomical niches with specific myeloid subset distributions (Figure 1B).
Although EGFP expression is lower in classical compared to non-classical monocytes (and has thus been used to track the latter), the discrimination between both subsets by imaging is imprecise.
The high expression of ECFP in the MacBlue mouse improves the detection of both subsets, but their discrimination is still not possible (53, 54).
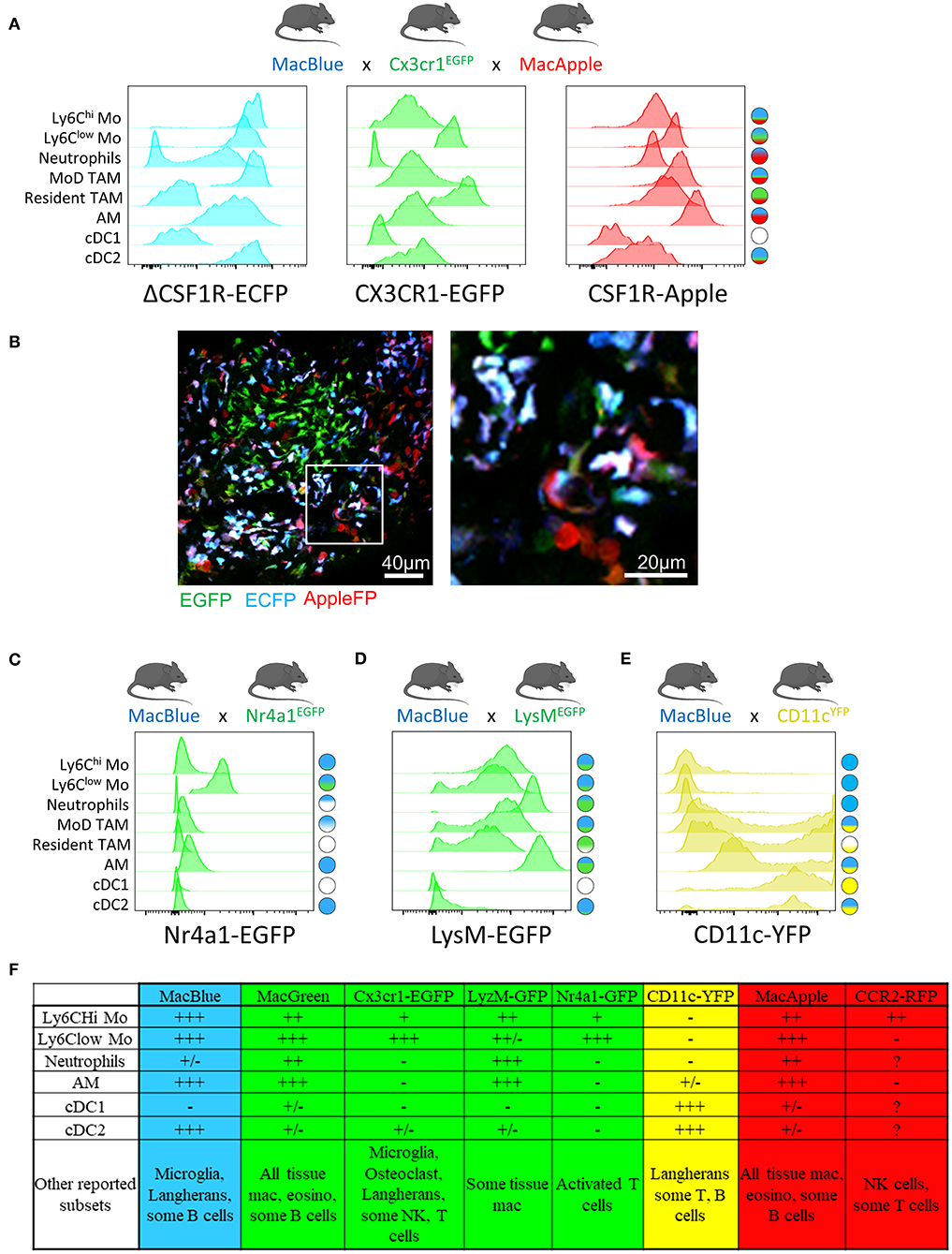
The Nr4a1gfp fluorescent reporter mouse provides a good marker to monitor non-classical monocytes in the lungs (28). Combination between MacBlue and Nr4a1gfp might offer an opportunity to simultaneously track both subsets (Figure 1C).
FP expression guided by the Ccr2 promoter would be expected to preferentially label classical monocytes, but this fluorescent reporter is also highly expressed on NK cells [(55) and personal observation].
NK cells are often abundant in the TME and can lead to misinterpretation of imaging studies using this reporter. Combination with other reporters may therefore improve specificity.
For instance, combining Ccr2RFP and Cx3cr1EGFP reporters allows tracking of the relative accumulation of CCR2hiCX3CR1low and CCR2lowCX3CR1hi cells in glioblastoma, arguing for distinct origins of TAM in this model (56)
As NK subsets also express EGFP in the Cx3cr1EGFP, the risk of NK contamination when imaging and identifyinng myeloid cells in the TME using this mouse strain must be considered.
LysMEGFP reporter mice display bright expression of GFP based on the lysozyme M locus and are widely used to visualize monocytes and macrophages. However, this marker is also strongly expressed in neutrophils (50, 57).
Using this reporter for live imaging is challenging as monocytes, macrophages, and neutrophils are closely related in the TME and the discrimination of these populations requires additional markers.
The combination of LysMEGFP with MacBlue might be considered, but the strong overlap of expression of these two reporters between granulocytes, monocytes and macrophages limits their accurate identification by imaging (Figure 1D).
Mouse strains expressing FP driven by the Itgax promoter (CD11c) typically provide very bright fluorescent signal and are available in different colors (58).
Although Itgax-based reporters are routinely associated with DC, it is clear that numerous TAM will express the FP and thus prevent the exclusive visualization of DC using this unique reporter (Figure 1E).
The combination of CD11cRFP with Cx3cr1EGFP in the study by Broz et al has provided an additional dimension to better discriminate DC and TAM in breast tumors (2).
The combination of CD11RFP and Xcr1venus reporters provides also an alternative to more accurately identify DC by imaging (59).
Altogether, these transgenic models have demonstrated utility in providing new insights on the dynamics of different myeloid populations (Figure 1F).
Furthermore, the combination of different fluorescent reporters appears to be a valid and worthwhile approach to target the cells more accurately. We have already demonstrated that the relative expression of the fluorescent reporter in MacBlue x Cx3cr1EGFP mice identifies TAM subsets of distinct origins with specific anatomic distribution (16).
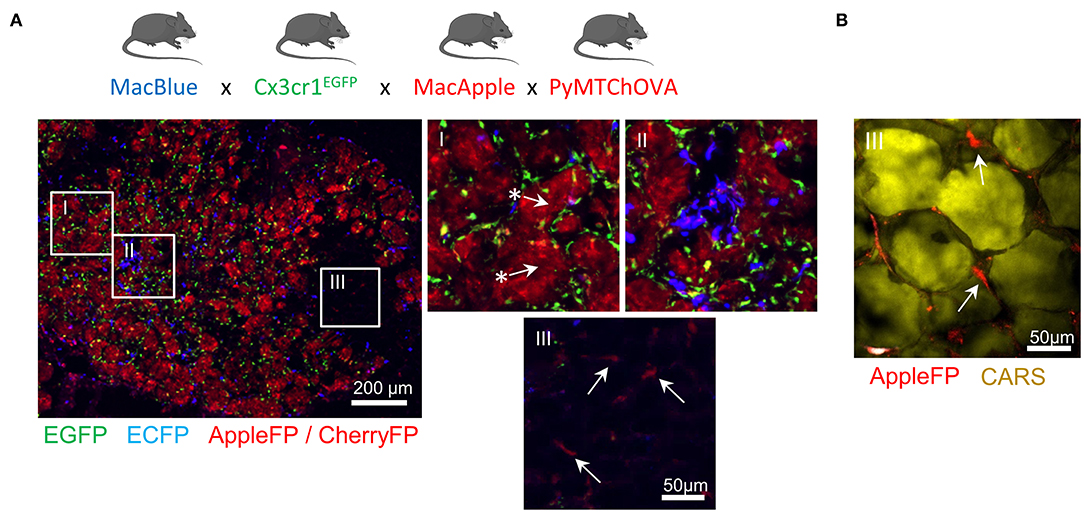
TAM microanatomical niches are even more marked in the spontaneous mammary tumor model PyMT-ChOVA combined with the MacBlue x Cx3cr1EGFP x MacApple reporters. Subsets with relative dominant expression of the three FP have been identified (Figure 2A). EGFP+ cells are mainly localized to the neoplasic mammary epithelium basal membrane and ECFP+ are more clustered in the stroma.
In addition to genetic fluorescent reporters, two-photon imaging can be used to generate fluorescence from specific cellular structures without the need of an exogenous fluorescent probe. Coherent anti-Stokes Raman scattering (CARS) imaging, for example, allows imaging of lipid deposits showing that a MacApple+ subset is enriched in the adipose tissue of the PyMT tumors (Figure 2B) and favoring the notion of spatial diversity of TAM (60).
Whether or not these subsets originate from resident macrophages of the mammary epithelium and surrounding adipose tissue needs further investigation. Second harmonic generation (SHG) is another label-free approach based on the intrinsic optical properties of extracellular structures that has been used to highlight T-cell trafficking in the collagen matrix of the TME (61, 62).
Tracking the evolution of collagen density according to tumor stage can be correlated with the functional characterization of TAM, as they are major actors in ECM remodeling. Szulczewski et al. have reported a label-free metabolic imaging protocol allowing for the visualization of NADH and FAD based on their autofluorescent properties.
This technique has identified that macrophages express high levels of FAD and are mainly glycolytic, enabling their discrimination from tumor cells without adding any exogenous staining molecule (63).
Label-free sensing of biomolecules typically does not result in photobleaching and reflects physiological content and distribution when compared with exogenous fluorescent probes. This label-free imaging also provides an opportunity to obtain information from human samples. As these methods lack specificity, complementary markers are necessary to study myeloid function.
Funding: The research was supported by the National Cancer Institute (F31CA232531) and the National Institute of Neurological Disorders and Stroke. (R21NS106554-01).
Source:
Emory Health Sciences
Media Contacts:
Quinn Eastman – Emory Health Sciences
Image Source:
The image is credited to Zhihong Chen.
Original Research: Open access
“Intravital 2-photon imaging reveals distinct morphology and infiltrative properties of glioblastoma-associated macrophages”. Zhihong Chen, James L. Ross, and Dolores Hambardzumyan.
PNAS. doi:10.1073/pnas.1902366116



{kind=link}