











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Scientists at the University of Warwick have discovered a new process that sets the fastest molecular motor on its marathon-like runs through our neurons.
The findings, now published in Nature Communications, paves the way towards new treatments for certain neurological disorders.
The research focuses on KIF1C: a tiny protein-based molecular motor that moves along microscopic tubular tracks (called microtubules) within neurons. The motor converts chemical energy into mechanical energy used to transport various cargoes along microtubule tracks, which is necessary for maintaining proper neurological function.
Neurons are cells that form the basis of our nervous system, conducting the vital function of transferring signals between the brain, the spinal cord and the rest of the body. They consist of a soma, dendrites, and an axon, a long projection from the cell that transports signals to other neurons.
Molecular motors need to be inactive and park until their cargo is loaded onto them. Neurons are an unusually long (up to 3 feet) type of nerve cell, and because of this marathon distance, these tiny molecular motors need to keep going until their cargo is delivered at the end.
Insufficient cargo transport is a crucial cause for some debilitating neurological disorders. Faulty KIF1C molecular motors cause hereditary spastic paraplegia, which affects an estimated 135,000 people worldwide. Other studies have also found links between defective molecular motors and neurological disorders such as Alzheimer’s disease and dementia.

The research shows how, when not loaded with cargo, KIF1C prevents itself from attaching to microtubule tracks by folding on to itself.
The scientists also identified two proteins: PTNPN21 and Hook3, which can attach to the KIF1C molecular motor. These proteins unfold KIF1C, activating it and allowing the motor to attach and run along the microtubule tracks—like firing the starting pistol for the marathon race.
The newly identified activators of KIF1C may stimulate cargo transport within the defective nerve cells of patients with hereditary spastic paraplegia, a possibility the team is currently exploring.
Commenting on the future impact of this research, Dr. Anne Straube from Warwick Medical School said: “If we understand how motors are shut off and on, we may be able to design cellular transport machines with altered properties.
These could potentially be transferred into patients with defect cellular transport to compensate for the defects.
Alternatively they can be used for nanotechnology to build new materials by exploiting their ability to concentrate enzymes or chemical reagents. We are also studying the properties of the motors with patient mutations to understand why they function less well.
“We still know very little about how motors are regulated.
There are 45 kinesins expressed in human cells, but we only have an idea how the motors are activated for less than a handful of them.
KIF1C is the fastest motor in neurons and the motor that is the most versatile—it delivers cargoes efficiently to all processes in a neuron, not just the axon.”
The molecular motor KIF1C is a fast plus end-directed kinesin which is important for cargo transport in several cell types, delivering dense core vesicles to the dendrites of neurons and trafficking integrins to the actin-rich adhesions of macrophages and smooth muscle cells.
It has been suggested that like other kinesin-3 motors KIF1C may cooperate with dynein to enable bidirectional cargo transport, but how this mechanism is regulated is unclear.
Most kinesin-3 motors are inactive, diffusive monomers which dimerise upon cargo binding, activating their processive movement.
This mechanism of autoregulation is distinct to kinesin-1 and kinesin-7 motors, which are regulated through direct interactions between the tail domains and the motor and neck regions, inactivating them until cargo binding relieves inhibition.
Key Findings:
In this paper Siddiqui et al. show that unlike other Kinesin-3 motors, purified KIF1C is intrinsically dimeric.
In high salt concentrations, KIF1C adopts a more elongated conformation than in physiological conditions which suggests that the non-motor regions may fold into a compact, self-interacting structure.
Using crosslinking mass spectrometry the authors show that the microtubule-binding interface of the motor domain interacts with the FHA domain and the third coiled-coil in the stalk.
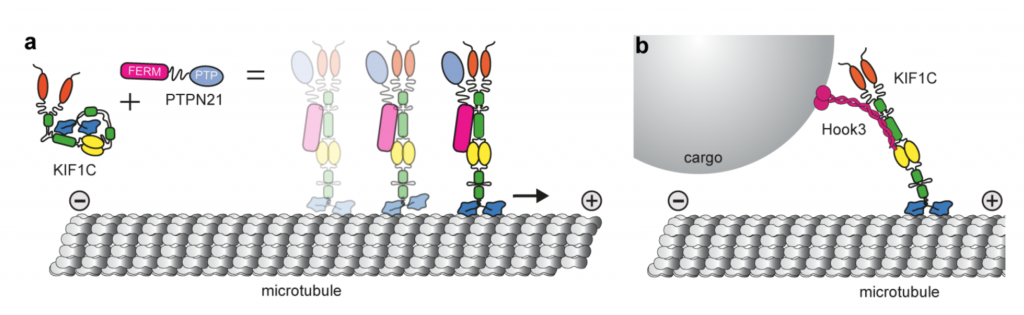
This indicates that such self-interactions may block the motor domain from associating with the microtubule and subsequent processivity(Fig. a).
They also find that depletion of a previously characterized KIF1C interactor, PTPN21 (protein tyrosine phosphatase N21), phenocopies KIF1C depletion resulting in a reduction in the frequency of podosome formation.
Interestingly, rescue is not dependent on the catalytic activity of PTPN21, as a minimal region of PTPN21 containing only the FERM domain (Fig. a) was sufficient to rescue the phenotype, suggesting that the scaffolding function was important for KIF1C activation.
By imaging vesicle-transport in both RPE1 cells and primary hippocampal neurons Siddiqui et al. show that the reduction of both plus and minus end directional transport observed upon partial depletion of KIF1C can be rescued by over expression of the PTPN21 FERM domain.
This demonstrates that this activation mechanism is common between different cell types.
Using in vitro reconstitutions, the authors show that the PTPN21 FERM domain is able to activate KIF1C.
This is shown by an increase in both landing rate and frequency of motor movement. Using crosslinking mass spectrometry, they find that PTPN21 FERM domain forms crosslinks with the FHA/coiled-coil 3 (CC3) region of KIF1C responsible for its auto inhibition.
To further confirm that this region is important for autoregulation, they generated a KIF1C_delCC3 mutant lacking the CC3 domain. Single molecule motility assays indicate that KIF1C_delCC3 has a higher landing rate than KIF1C in vitro.
Additionally, the landing rate of the mutant is no longer increased by inclusion of PTPN21-FERM, indicating that inhibition is fully relieved in the absence of the coiled-coil 3 domain.
Finally, Siddiqui et al. use a BioID to look for other KIF1C stalk interactors. The authors identified 240 proteins which are pulled down with KIF1C-BioID2 but not KIF1C_del623-825-BioID2.
Interestingly, their top hit is Hook3, a dynein/dynactin activator that may be involved in mediating bidirectional cargo transport.
In vitro they demonstrate that Hook3 is able to activate KIF1C in a similar fashion to PTPN21 and show co-transport of Hook3-Alexa647 with KIF1C in their single molecule kymographs.
Overall this paper shows that KIF1C is a dimeric kinesin-3 motor and is autoinhibited through a direct interaction between the FHA/CC3 stalk region and the microtubule-binding region in the motor domain.
This inactive, compact structure can be relieved through interaction with binding partners such as PTPN21 or Hook3, increasing the landing rate of the motor on microtubules in vitro and enabling bidirectional cargo transport in cells (Fig. a,b).
More information: Nida Siddiqui et al. PTPN21 and Hook3 relieve KIF1C autoinhibition and activate intracellular transport, Nature Communications (2019). DOI: 10.1038/s41467-019-10644-9
Journal information: Nature Communications
Provided by University of Warwick



{kind=link}