











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



New research at the University of Kentucky has confirmed that the presence of XX sex chromosomes increases the amount of fat circulating in the blood, which leads to narrowing of the arteries and ultimately a higher risk of heart attacks and coronary artery disease.
The research was published in June 2019 in Nature Communications.
The leading cause of death in women is coronary artery disease (CAD), but women develop CAD almost 10 years later than men.
For many years, scientists attributed this decade-long delay in disease development to the protective effects of sex hormones.
There is a lot of evidence that hormones like estrogen and progesterone protect the heart, but scientists had little data on the influence of the genetic component – the X chromosome – on the heart.
A team led by Lisa Cassis, a researcher in the UK College of Medicine’s pharmacology and nutritional sciences department, set out to understand the role of sex chromosomes in the cardiovascular system.
Cassis’s team studies chromosome effects in mice, and for this most recent discovery they were able to zero in on XX chromosomes by removing hormones.
According to Yasir Al-Siraj, a postdoctoral scholar and the paper’s first author, if the levels of circulating lipids transported by the blood are too high, they will start to accumulate in and on the artery wall, leading to plaque buildup.
These plaques harden and narrow the artery, reducing blood flow to the vital organs.
The team looked at lipids absorbed from the diet and made in the liver.
Cassis, who also serves as the UK vice president for research, said, “We looked at how our X sex chromosomes were influencing the levels of lipids in the blood and in the arteries.” What they found is that an XX sex chromosome combination promotes efficient use of fat.
Women need fat to bear and feed babies, Cassis explained:
“We’re set up, potentially through our XX sex chromosomes, so that we can effectively absorb that lipid from the diet and put it into our fat cells and maybe even make it in the liver.”
Everything is fine until women hit menopause and the protective effects of hormones disappear, leaving women with what Cassis calls “that XX thrifty, fat-absorbing kind of genotype.”
The team is looking at genes that are changed in the liver and in the intestine to find novel targets for drug development.
If they can find target genes that influence atherosclerosis, scientists can then explore the effects of existing drugs or develop new ones.
Al-Siraj said his next step is to study the role of the number of X chromosomes in atherosclerosis. “We don’t know if our findings are due to the presence of two X chromosomes or due to the absence of the Y chromosome,” he said.
These findings may also drive choice of diet for post-menopausal women. Cassis said, “For example, if they’re very effective fat absorbers, obviously, once they get post-menopausal, they need to be careful about the fat content.”
Sex differences in obesity and related diseases are well established.
Gonadal hormones are a major determinant of these sex differences.
However, sex differences in body size and composition are evident prior to exposure to gonadal hormones, providing evidence for gonadal-independent contributions attributable to the XX or XY sex chromosome complement.
Large-scale genetic studies have revealed male/female differences in the genetic architecture of adipose tissue amount and anatomical distribution. However, these studies have typically neglected the X and Y chromosomes.
Scope of the review
Here we discuss how the sex chromosome complement may influence obesity, lipid levels, and inflammation.
Human sex chromosome anomalies such as Klinefelter syndrome (XXY), as well as mouse models with engineered alterations in sex chromosome complement, support an important role for sex chromosomes in obesity and metabolism.
In particular, the Four Core Genotypes mouse model – consisting of XX mice with either ovaries or testes, and XY mice with either ovaries or testes -has revealed an effect of X chromosome dosage on adiposity, hyperlipidemia, and inflammation irrespective of male or female gonads. Mechanisms may include enhanced expression of genes that escape X chromosome inactivation.
Major conclusions
Although less well studied than effects of gonadal hormones, sex chromosomes exert independent and interactive effects on adiposity, lipid metabolism, and inflammation.
In particular, the presence of two X chromosomes has been associated with increased adiposity and dyslipidemia in mouse models and in XXY men. The enhanced expression of genes that escape X chromosome inactivation may contribute, but more work is required.Keywords: Genetics, Gonadal hormones, Adipose tissue, Sex chromosome anomalies, X chromosome inactivation, Mouse models
Sexual dimorphism in disease
The study of sex differences in physiology has gained traction as it pertains to understanding male/female differences in disease susceptibility.
Women more often than men suffer from autoimmune diseases (e.g., systemic lupus erythematosus, scleroderma, Sjogren’s syndrome), as well as osteoporosis, Alzheimer’s disease, and clinical depression [1], [2], [3], [4], [5], [6].
Autism occurs at higher rates in boys than girls, and sex differences are also observed in neurological diseases such as Parkinson’s disease and schizophrenia, to name a few [7], [8], [9].
In cardiovascular disease and stroke, there are differences between the sexes in age of onset, disease pathology, and mortality [10].
Some sex differences in human disease are directly related to the distinct sex chromosome complement in females (XX) and males (XY).
Males are more commonly afflicted with X-linked diseases such as color blindness, Duchenne muscular dystrophy, and hemophilia.
Women may be protected from (or experience attenuated severity of) these recessive disorders, because they possess two X chromosomes [11].
The random inactivation of one X chromosome in female cells during early development silences the X chromosome carrying a mutant gene in approximately half of female cells, allowing the exclusive expression of a wild-type gene in those cells.
An extreme example of the influence of XX versus XY genotype on disease severity is Rett syndrome, which results from mutation in the MECP2 gene on the X chromosome.
Females afflicted with Rett mutations experience progressive neuro-developmental deficiencies leading to impaired learning, communication, coordination, and other brain functions. By contrast, males with MECP2 mutations typically die in utero or in infancy [12].
Here we will discuss how the sex chromosome complement may influence factors that underlie metabolic disturbances such as obesity, dyslipidemia, and inflammation.
It is well known that gonadal hormones have strong effects on fat storage and susceptibility to related diseases such as cardiovascular disease and type 2 diabetes [13].
Since female gonads are typically found together with XX chromosomes, and male gonads with XY chromosomes, the independent roles of gonadal hormones and sex chromosomes has not been appreciated.
However, human sex chromosome anomalies such as Klinefelter syndrome (XXY) and Turner syndrome (XO), as well as mouse models with engineered alterations in sex chromosome complement, support an important role for sex chromosomes in obesity and metabolism.
Sex differences in human adiposity and obesity
Adipose tissue serves as an efficient energy storage depot. It also has active roles in fatty acid uptake from circulating lipoproteins produced in response to a meal, and in the regulated release of fatty acids for use by other tissues between meals or during physical activity.
There are key differences between males and females in adipose tissue distribution, with men accumulating greater amounts of visceral adipose tissue and women typically having greater fat accumulation in subcutaneous (gluteal–femoral) depots [14], [15].
Sex-specific fat distribution is influenced by several factors, including diet and hormonal status [16]. One contributor to sex bias in adipose tissue distribution may be the rate of direct fatty acid uptake by tissues, a process that occurs independently of lipoprotein lipase (the enzyme responsible for liberation of fatty acids from lipoproteins).
Direct fatty acid uptake is higher in the gluteal–femoral depot in women and in the abdominal depot in men [17].
Both overall fat mass and visceral fat accumulation are strongly associated with the development of cardiovascular disease, stroke, hypertension, and insulin resistance [15], [18], [19].
Standard measurements for fat accumulation in humans include body mass index (weight as a function of height), which reflects whole body adiposity, and waist-to-hip ratio, which provides an indication of fat distribution, with the waist measurement as a proxy for visceral fat and the hip measurement for gluteal fat [19].
Seminal studies performed in the 1980’s provided evidence that overall adiposity, as well as subcutaneous fat mass, have a heritability of approximately 30% [20].
This estimate was corroborated by a 1990 study in Caucasian male twins that showed a 31% heritability of waist-to-hip ratio, while a more recent population-based study estimated heritability of the same trait at 39% [21], [22].
Some estimates indicate that heritability of fat distribution is greater in women than in men [19].
Fat distribution and heritability also differ across ethnic groups [23], [24].
One approach to identify the genes contributing to sex differences in adipose tissue accumulation and distribution is genome-wide association studies (GWAS) in large human cohorts.
This approach types genetic variants across the genome and correlates their occurrence with a trait to identify loci that are associated. GWAS performed in hundreds of thousands of people have identified more than 100 genetic loci that harbor common genetic variants that influence adiposity [25], [26], [27].
Importantly, at least 17 loci that are associated with body mass index have also been identified in GWAS for type 2 diabetes [25].
Analysis of accumulated GWAS data has revealed a distinct genetic architecture for loci affecting adiposity in males and females.
For example, a meta-analysis of more than 50 GWAS studies with waist-to-hip ratio (adjusted for total fat) in more than 200,000 individuals identified 49 loci, 20 of which showed sex-specific effects, with 19 of these having stronger effects in women [28].
These loci represent a rich resource for the identification of sex-biasing genetic factors for body composition and fat distribution, although at present, these loci together account for only a few percent of the genetic variance in adiposity.
Much work remains to be done, including the identification of the causal variants at each locus and their mechanism of action to influence adiposity. Of note, these analyses did not take into account loci on the X or Y chromosomes, leaving a gap in our knowledge regarding how genetic variations on X and Y may contribute to observed sex differences in adiposity. The following sections describe studies outside of GWAS that have informed about the role of the X and Y chromosomes in adiposity and metabolic disease.
Sex differences in adipose tissue expansion
It has been suggested that adipocytes in gluteal–femoral depots (and other subcutaneous depots) confer better metabolic health because of the ability to expand to store more fat by recruiting new adipocytes [16], [29], [30].
It has previously been thought that male mice exhibit greater diet-induced fat mass expansion (in both visceral and subcutaneous depots) than females, and this is partly due to effects of sex hormones [31], [32], [33].
However, a recent study demonstrates that in C57BL/6J mice, the sex differences in diet-induced weight gain depend strongly on the age of the mice when fed a high fat diet. In juvenile mice (aged 6 weeks), feeding a high-fat diet for 3 months led to greater percent weight gain in males than females.
However, in adult mice (aged 31 weeks), the trend was reversed, and females gained substantially greater percent body weight in response to high-fat diet [34].
One potential contributor to sex differences in adipose tissue expansion is the numbers of adipocyte precursor cells (pluripotent stem cells that may differentiate into adipocytes, chondrocytes or osteoblasts) in mouse gonadal or subcutaneous fat depots.
On a low-fat chow diet, female C57BL/6J mice have more adipocyte precursor cells than males in gonadal (visceral) and inguinal (subcutaneous) fat pads [35], [36], [37]. When fed a high-fat diet (45% calories as fat), female mice showed increased adipocyte precursor cells and mature adipocytes in gonadal fat, but males did not increase mature fat cells in the gonadal fat pad [36].
Other studies, which employed pulse-labeling or lineage tracing to follow the fate of proliferating adipocyte progenitor cells, inferred that male gonadal fat exhibits hyperplasia in response to a high-fat diet, whereas females exhibit adipocyte hyperplasia in both gonadal and subcutaneous fat depots [38], [39].
The sex-specific patterns were reversed by ovariectomy in female mice, or estrogen administration in male mice, suggesting a role for gonadal hormones [37].
These studies have proved valuable, but additional studies of adipocyte recruitment and turnover in fat depots of both sexes are needed to clarify discrepancies between studies that have used distinct methodologies and to provide additional details.
A role for XX chromosome dosage in sex differences in adiposity
Conclusive studies of metabolic changes in XXY men and XO women have been hampered by the lack of suitable control groups with similar gonadal hormone levels, by limited statistical power due to small cohorts, and by heterogeneity among individuals in the genetic and environmental factors that influence obesity.
The use of mouse models makes it possible to overcome some of these limitations and to evaluate the relative contributions of sex chromosomes and gonadal hormones to sex differences.
A traditional approach used in mouse models to identify the cause of sex differences is to assess the action of gonadal hormones after puberty [70].
Adult female and male mice are gonadectomized and assessed to find whether the original sex difference is eliminated. If so, replacement of hormones by an implanted pump or daily injection is performed and mice observed to determine if the original sex difference is restored.
Follow-up studies may include identification of the hormone receptor that mediates the effect, which can be done through chemical or genetic inhibition of hormone receptor function. For sex differences that occur before puberty, or which are not altered by gonadectomy, the role of the surges in testosterone that occur pre- or –postnatally may be studied. For meaningful performance and interpretation of studies that manipulate gonadal hormones, many factors must be considered and are reviewed elsewhere [70]. If gonadal hormone differences cannot fully explain a sex difference, the potential effect of sex chromosome complement should be considered.
A mouse model that has been specifically designed to parse the effects of gonadal hormones as well as sex chromosomes is the Four Core Genotypes (FCG) mouse.
The model involves the relocation of the Sry gene, which promotes the development of testes, from the Y chromosome to an autosome such that gonadal sex can segregate independently from sex chromosome complement [70], [71].
The position of the Sry gene on an autosome does not alter the prenatal or adult androgen levels [72]. Studies performed with FCG mice involve the comparison of four genotypes, which shuffle the chromosomal and gonadal components of sex to generate XX mice with either ovaries or testes and XY mice with either ovaries or testes (referred to, respectively, as XX F, XX M, XY F, and XY M).
A two-by-two comparison of these four genotypes allows identification of effects that are dictated by either gonadal type or sex chromosome type (Figure 1). To eliminate the contribution of circulating gonadal hormones, the FCG mice are often studied after gonadectomy as adults.

The Four Core Genotypes (FCG) mouse model. The FCG mouse model uncouples chromosomal sex from gonadal sex to reveal contributions of each to traits of interest. Traits that are influenced by gonadal hormones (or other effects conferred by ovaries or testes) appear similar between XX and XY mice with ovaries, and between XX and XY mice with testes, whereas the groups with ovaries and testes differ from one another. Conversely, traits that are influenced by sex chromosome complement appear similar between XX animals with ovaries or testes, and XY animals with ovaries or testes, and the XX and XY groups differ from one another. For information on the derivation of FCG mice, see Ref. [71].
Using the FCG model, effects of sex chromosome complement on several metabolic traits have been identified, including obesity, plasma lipid profile, food intake, and hypertension [73], [74], [75], [76], [77].
In C57BL/6 FCG mice that were gonadectomized as adults to remove acute effects of gonadal hormones, mice with XX versus XY chromosomes (regardless of male or female gonads present originally) gained weight more rapidly on a chow or high-fat diet, and accumulated greater body fat, particularly in the inguinal subcutaneous fat depot (Figure 2) [73].
Increased weight gain in XX compared to XY mice was promoted, at least in part, by enhanced food intake by XX mice, specifically during the light phase of the circadian cycle [74].
As nocturnal animals, mice consume approximately 70% of their calories during the dark phase, and they rest and snack (30% of total calories) during the light phase.
The enhanced food intake by XX mice in the light phase was due to anticipatory eating beginning about 1.5 h before their XY counterparts [74].
In addition to increased adipose tissue accumulation, XX mice fed a high-fat diet for 16 weeks developed obesity-related conditions such as fatty liver, elevated total blood cholesterol levels, and elevated insulin levels [73].
It is likely that these metabolic disturbances are secondary to the enhanced fat accumulation in XX mice fed a high-fat diet since they are not seen in XX mice fed a chow diet.

Summary of the influence of sex chromosome complement on adiposity and related traits in mice and humans. (A) Studies of the Four Core Genotypes mice revealed that the presence of XX chromosomes leads to increased body weight, fat mass and related morbidities compared to XY mice (all mice gonadectomized as adults to remove acute effects of gonadal hormones). (B) Studies of XY* mice determined that the presence of two X chromosomes promotes greater body weight and fat mass than those with one X chromosome, with no evident effect of the Y chromosome (all mice gonadectomized as adults). (C) XXY compared to XY men show greater abdominal fat, insulin resistance and related metabolic abnormalities. XXY men also have reduced gonadal hormone levels. (D) XO compared to XX women show increased body fat, but also suffer from developmental and hormonal abnormalities. Data are summarized from studies cited in the main text.
Data from the FCG mice clearly showed an effect of XX compared to XY chromosomes on adiposity and obesity-related morbidities.
But these studies alone did not distinguish between an effect of X chromosome dosage (two versus one X) or an effect of the presence or absence of the Y chromosome.
Analysis of mouse models with additional sex chromosome genotypes made it possible to identify the relevant sex chromosome effect.
By making use of an unusual Y chromosome with sequence duplications in the pseudoautosomal region (known as the XY* model, [71]), mice with XX, XY, XO and XXY chromosomes were assessed for body weight and adiposity.
The results showed that the presence of two X chromosomes (XX and XXY mice) led to higher body weight/fat than one X chromosome (XY and XO); the presence of the Y chromosome did not have an influence (Figure 2) [73].
More information: Yasir AlSiraj et al, XX sex chromosome complement promotes atherosclerosis in mice, Nature Communications (2019). DOI: 10.1038/s41467-019-10462-z
Journal information: Nature Communications
Provided by University of Kentucky
Sex Determination
Humans have 46 chromosomes, which exist in pairs. Therefore, we have 23 pairs of chromosomes. Of these, 22 are called autosomes, and one pair is called the sex chromosomes. Autosomes determine our characteristics, such as height, hair color, skin color, metabolic rate, etc. while the sex chromosomes determine the sex of the individual. There are 2 sex chromosomes – X and Y. Females have 22 autosomes and two X chromosomes, i.e., 22 + XX, while males have 22 autosomes and an X and Y chromosome each, i.e., 22 + XY.
This therefore proves that X is the female chromosome, and Y is the male chromosome. Why then, are men not YY? Does having an X chromosome make them only half men? Or would having YY make them super-men?
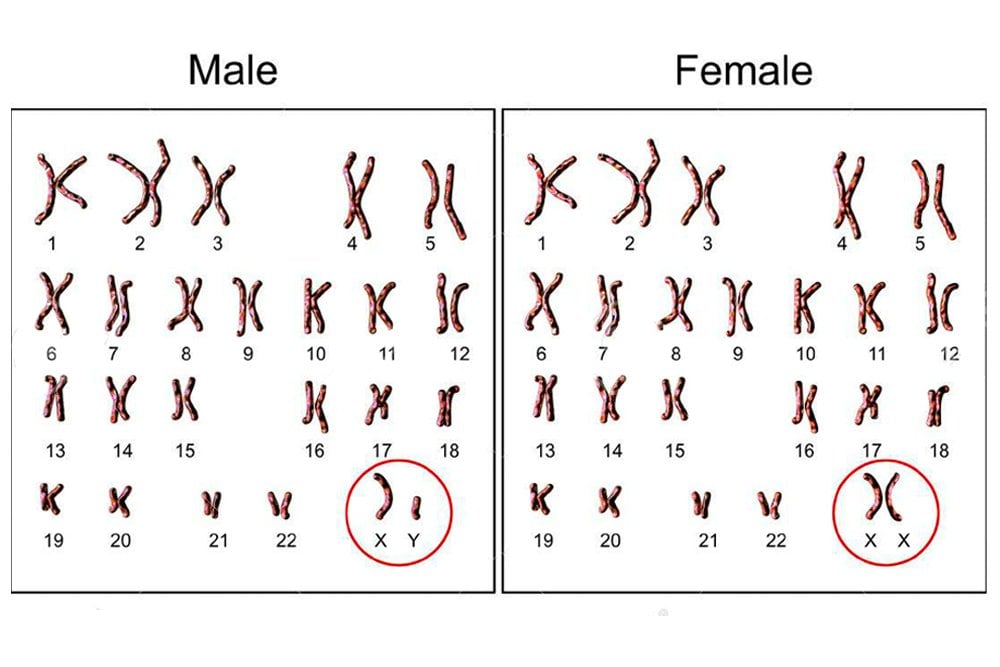
Sex determination in humans
Why isn’t YY possible?
While they are called sex chromosomes, the X and Y chromosomes also carry information for purposes apart from sex determination. Let’s do a brief recap before we delve into these details. Chromosomes are the genetic material in our cells. Each chromosome has a large number of genes on it, which are the actual information-carrying structures.
While our sex chromosomes are responsible for determining our sexual nature, they also have genes for various other functions. These functions may or may not be related to our sexual nature and development. Now, here is where the tricky part begins. The X chromosome has about 800-900 genes, in contrast to the Y chromosome, which has a meager 60-75 genes. However unbelievable, that is the truth.
The X chromosome has certain genes on it that are imperative for the survival of the fetus. Therefore, for any individual to survive, it requires at least one X chromosome. If an organism is YY, it won’t be a super-man, it would just be super-dead!
What genes are missing on the Y chromosome?
There are a total of 20,000 genes in humans. The X chromosome has approximately 5% of the DNA required for a person to survive, while the Y chromosome has less than 2%. Obviously, without an X chromosome, the individual would perish, but are the genes on the X chromosome even that important?
The X chromosome has a gene known as DMD, which is responsible for the formation of Dystrophin, a protein. Dystrophin is required for the formation of the skeletal and cardiac muscles. It strengthens the muscles and protects them from damage. It is also present in small amounts in the nerves of brain cells.
Another critical gene is the CYBB gene. This codes for the formation of a protein known as cytochrome b-245. This protein is required for the formation of an enzyme called NADPH oxidase, which is an essential part of the immune system.
X and Y chromosome
These are just 2 among a number of genes that are missing from the Y chromosome. As is clearly evident, without these genes, it is impossible for a human to survive, so every individual requires at least one X chromosome.
Although the Y chromosome lacks these genes now, there was a time when it did have the required genes for the normal survival of a human.
Unfortunately, over the course of 300 million years of evolution, it lost almost 95% of its genes.



{kind=link}