









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Since the Middle East respiratory syndrome coronavirus (MERS-CoV) first emerged in Saudi Arabia in 2012, there have been more than 2,400 confirmed cases of the infection, resulting in greater than 800 deaths – an alarming fatality rate of 35 percent.
For this reason, researchers have been eager to identify any risk factors that contribute to the development of severe or lethal disease.
Current clinical evidence points to diabetes as a major risk factor in addition to other comorbidities including kidney disease, heart disease, and lung disease.
Researchers from the University of Maryland School of Medicine (UMSOM) and the Johns Hopkins University School of Medicine have demonstrated in a new study, published earlier this week in the Journal of Clinical Investigation Insights, how diabetes contributes to mortality from MERS-CoV infections, and the finding could shed light on why other respiratory illnesses like the flu or pneumonia might strike those with diabetes more severely.
They investigated the connection between diabetes and MERS-CoV in a mouse model and discovered that although the virus did not replicate more readily in the diabetic mice compared to the healthy controls, the diabetic mice exhibited a delayed and prolonged inflammatory response in the lung.
Diabetic mice had lower levels of inflammatory cytokines and fewer inflammatory macrophages and T cells.
This indicates that the increased severity of MERS-CoV infection in patients with diabetes was likely due to a malfunction in the body’s response to infection.
“Understanding how diabetes contributes to disease severity following MERS-CoV infection in this context is critical,” said Matthew Frieman, Ph.D., associate professor of microbiology and immunology who is the corresponding author of the study.
“Our next step is to determine what drives the altered immune response in diabetics and how to reverse those effects with therapeutics for treatment of patients.”

Follow up research could also explore whether health care providers should double their efforts to manage and stabilize glucose levels in patients with diabetes experiencing a dangerous respiratory infection and whether better management would help mitigate the effects of these infections.
“This is an important finding for patients with diabetes and physicians who treat them,” said UMSOM Dean E. Albert Reece, MD, Ph.D., MBA, who is also the Executive Vice President for Medical Affairs, University of Maryland and the John Z. and Akiko K. Bowers Distinguished Professor.
“We have long known that diabetic patients have worse outcomes when they get a serious infectious disease, but this new insight on immune function could pave the way for better treatments.”
June 19th, 2017, a case of Middle East respiratory syndrome coronavirus (MERS-CoV) is reported by the National International Health Regulation (IHR) focal point of Lebanon.1 As a novel coronavirus, MERS-CoV was first identified in a patient from the Kingdom of Saudi Arabia in June 2012 and was originally named human coronavirus-EMC, as in Erasmus Medical Center.2
The case involved a man in Saudi Arabia who was admitted to the hospital with pneumonia and acute kidney injury.3
The virus was later isolated from the patient’s sputum.
A different report appeared in September 2012, detecting a similar virus with 99.5% identity in a previous patient who initially developed symptoms in Qatar and had traveled to Saudi Arabia before the disease progression exacerbated.4
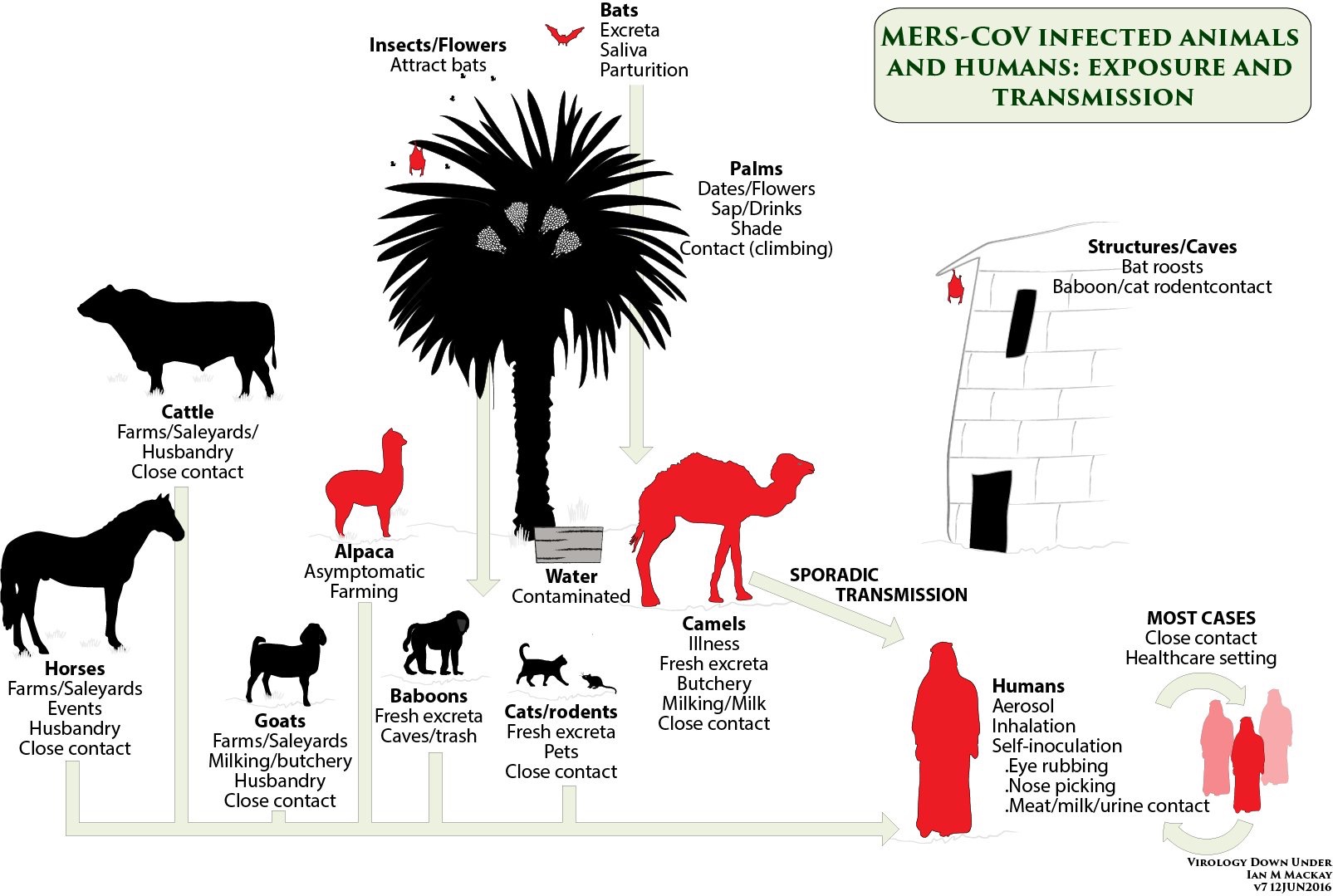
Seroepidemiologic and virologic research revealed MERS-CoV infection in dromedary camels (Camelus dromedarius) with isolated viruses from dromedaries competent in infecting the human respiratory tract.5,6
All cases of MERS-CoV have been established in the Middle East and North Africa region, Europe, East Asia and the United States.7
With numerous consecutive cases and clusters of infections having been reported since September 2012, the World Health Organization (WHO) was notified to date with 2,040 laboratory-confirmed cases of MERS-CoV infection, 712 fatalities and a total of 27 differing countries reporting cases.1
This paper discusses the virology, pathogenesis, and epidemiology of MERS-CoV, highlighting possible sources and modes of transmission and clinical manifestation along with treatment and prevention while providing a One Health approach to tackle the complex issue of MERS-CoV.
Virology and pathogenesis
MERS-CoV belongs to the family Coronaviridae and has a large RNA viral genome of approximately 26-33 kb and G+C contents varying from 32 to 43%.8,9
MERS-CoV descends from the 2C beta-coronavirus lineage existing in humans and camels.10 Although genomically related, it is separate from other human beta-coronaviruses such as severe acute respiratory syndrome (SARS) coronavirus, but it is similarly associated with few bat coronaviruses.2,3,11 The functional receptor of MERS-CoV is dipeptidyl peptidase 4 (DPP4) and its expression induces the infection. The DPP4 protein expresses large quantity conversion of amino acid sequences in various species.11,12 MERS-CoV has the capability of infecting multiple human cell lines.13 This mainly comprises the lower respiratory tract in which respiratory epithelial cells are highly susceptible, encouraging viral replication productivity in human macrophages and dendritic cells, inducing production of pro-inflammatory cytokines, targeting T lymphocytes and leading to their apoptosis.14 Kidneys are also significantly affected by MERS-Co viruses, in addition to intestinal, liver cells, and histiocytes.14
Furthermore, in vitro studies illustrate that MERS-CoV can infect non-human primate, porcine, bat, civet, rabbit, as well as horse cell lines.13,15 The pathogenesis of MERS-CoV infection to date is not well understood and further research is necessary. MERS-CoV causes disease of the lower respiratory tract and can be found in tracheal aspirates, sputum, or bronchoalveolar lavage fluid of symptomatic patients. According to highly conserved coronavirus RNA polymerase chain reaction (PCR) analysis (or Pan-Corona-PCR), viral shedding from nasal secretions, sneezing, coughing and sputum can be continuous up to six weeks, which builds concerns that asymptomatic individuals could also transmit the infection to others.6,16 It is important to note that DPP4 is expressed in the upper respiratory tract of camels, while in humans it is solely expressed in the lower respiratory tract, a fact which may explain the limited detection of human-to-human transmission to date.17 MERS coronaviruses are capable of causing human and veterinary outbreaks. This is due to the fact that CoV have the potential to recombine, mutate and jump between animals, thus infecting a wide variety of species.18 Moreover, recent phylogenetic analysis has demonstrated that MERS-CoV reported from camels and humans are clustered together, while bat and hedgehog MERS-CoVs established a basal paraphyletic group to the camel and human MERS-CoV clade.19 It is estimated that the genome-wide substitution rate of nucleotides in camels and humans MERS-CoVs is 4.81 × 10-4 per site per year.19
Five bat cells (M. ricketti lung, P. abramus kidney, R. sinicus kidney and lung, and R. leschenaultii kidney cells) have been reported to be susceptible to MERS-CoV EMC/2012.20 In addition, various MERS-CoV strains have exhibited different infectivity on bat cell lines. For instance, M. ricketti lungs sustained infection by strains D1189.1/15 and D1271.1/15, while R. sinicus lung and R. leschenaultii kidney exhibited infection by strain D1189.1/15. None of the bat cell lines have shown sustainable infection by strain D998/15.20
MERS-CoV causes mild or no respiratory and clinical symptoms in camels and as a consequence it is not easily recognized or difficult to clinically diagnose. MERS-CoV is mainly acquired in dromedaries of less than one year of age.21 A case in 2013 has been reported in a camel with MERS-CoV, symptomatic with fever and rhinorrhea.22 Bats on the other hand are believed to be carriers of MERS-CoV without demonstrating apparent clinical signs of the disease.23
The genome of MERS-CoV contains 30,119 nucleotides and a minimum of 10 predicted open reading frames, 9 of which are suggested to be displayed from a set of nested 7 sub-genomic mRNA. ORF1a and ORF1b are the replicase opening frames which cover the 5′ end region of the genome, while the 3′ end encodes structural proteins, including spike (S), membrane (M), nucleocapsid (N), envelope (E), as well as various accessory proteins such as 3, 4a, 4b, 5 and 8b.24 MERS-CoV prompts cytopathic effects and replicates at high titers allowing for a rapid genome characterization.24 Replicase gene phylogenetic analysis has demonstrated that the full sequenced genome of coronavirus HCoV-EMC/2012 is closely related to Tylonycteris bat coronavirus HKU4 (BtCoV-HKU4) and to Pipistrellus bat coronavirus HKU5 (BtCoV-HKU5).16,25
The spike (S) protein of β-CoV is a key in cross-species transmission as it mediates virus-receptor recognition and induces viral pathogenesis. The receptor binding domain (RBD) on S protein’s N-terminal is vital for the penetration of β-CoV into host cells. Further, the severity of infection and cross-species efficiency is influenced by the mutations in CoV RBD.26
Nine sites in the S protein of MERS-CoV have been found to have significant positive selection. This entails that the S protein has encountered strong evolutionary pressure during its transmission from its natural host to humans. In addition, six of these positively selected sites occupy the RBD.19
Epidemiology
As large, enveloped, positive-stranded RNA viruses, coronaviruses are known to cause respiratory and enteric disease in various animals (including cattle, chicken, bat, mice, alpacas, swine, dogs, horse) and in humans.8,27
Four separate human coronavirus (hCoV) genera circulate the globe, of which some are promoters of the common cold, with an additional hCoV being SARS-CoV that caused an outbreak in 2003.1 In 2012, as discussed above, two cases of severe hCoV infection were notified, and were caused by a novel virus type. Both patients had travelled to or resided in Saudi Arabia. Shortly thereafter, numerous cases were reported, with at least 62% of human symptomatic cases having not been detected, indicating the true number of cases inclined to be greater than reported.28
A retrospective analysis traced the initial outbreak in April 2012 to a hospital in the city of Al-Zarqa, Jordan.29 Although outbreaks of MERS-CoV have been majorly reported in the Middle East, namely in Kuwait, Yemen, Oman, Iran, Lebanon, United Arab Emirates (Table 1), and specifically in Saudi Arabia (with 85.8% of the reported cases), imported cases have also been observed in Europe, Asia and North America. These include countries such as the United Kingdom, France, Germany, Italy, Greece, Netherlands, Egypt, Tunis, Malaysia, South Korea and the USA.30,31
Table 1
Reported human cases of MERS-CoV in the Arabian Peninsula
| Country | Total cases* | Recoveries | Death |
|---|---|---|---|
| Saudi Arabia | 1431 | 521 | 520 |
| United Arab Emirates | 74 | 14 | 8 |
| Jordan | 27 | 1 | 7 |
| Qatar | 14 | – | 4 |
| Oman | 9 | 1 | 4 |
| Kuwait | 4 | – | 2 |
| Lebanon | 2 | 1 | – |
| Tunisia | 3 | 2 | 1 |
| Egypt | 1 | 1 | – |
| Iran | 5 | – | – |
| Algeria | 1 | – | 1 |
| Turkey | 1 | – | – |
All data are provided by the WHO and Health Ministries in 201630 and 2017.1
*Total cases also include asymptomatic cases or seropositive antibodies presence.
In the Middle East, the number of cases has significantly increased during March and April 2014, later declining in mid-May of 2014.3,10 However, there has been a notable outbreak of 186 confirmed cases across 16 hospitals present in South Korea in the period of May and June 2015 from an index case.32
Two positive cases of MERS-CoV have been reported in Lebanon so far. The national IHR Focal Point of Lebanon reported the country’s first MERS-CoV infection on May 8th, 2014 as laboratory-confirmed case.33
A Lebanese 60-year-old male healthcare worker presenting high fever was admitted to the hospital, with symptoms of dyspnea and productive cough, and tested positive for MERS-CoV.1
The patient reported neither contact with laboratory confirmed cases nor with animals and no travel history during the 14 days prior to symptom onset. However, the patient had previously travelled around a couple of Gulf regions.
The second Lebanese case involved a 39-year-old male from Saudi Arabia. He developed mild symptoms and as a healthcare worker along the enhanced MERS-CoV surveillance in Saudi Arabia, a nasal swab was collected on June 11th, 2017 testing negative for the infection; the same day the patient travelled to Lebanon with no reporting symptoms.1 On June 15th, the patient developed gastrointestinal symptoms, diagnosis confirmed pneumonia, and the following day he tested positive for MERS-CoV. MERS-CoV appears to possess a predilection for males as approximately 64% cases are male with a median age of 48 years (range 9 months to 94 years) with nearly 35% case-fatality rate.1,31
A seasonal pattern of cases has been observed, with a peak each year between April and June, although the reason has yet to be elucidated.10 A potential explanation for this could be the camel birthing season, when younger camels are more prone to infection as opposed to mature ones. Moreover, it may be argued that the current upsurge in the number of infected patients is due to enhanced case detection and active surveillance programs.10
Sources and transmission
In an effort to examine the possibilities of domestic animals as reservoir, serological studies were performed in Saudi Arabia from 2010 to 2013, and in 2014, where sera from sheep, goats, cattle, and chickens from various geographical areas within the country were collected. All samples tested seronegative for MERS-CoV. Similar studies on equids in the United Arab Emirates and Spain, goats, sheep and cattle in Jordan and Europe also indicated negative results.34,35,36,37 Dromedary camels have been demonstrated to be the primary animal host for MERS-CoV.5,6,14,38
Yet, all tested healthy alpacas in a recent Qatar study have been demonstrated to be positive for MERS-CoV.38
Human-to-human transmission occurs but seems limited to families and close contacts, or is due to nosocomial infection.2,39
Camel-to-human transmission of MERS-CoV has been evident in Saudi Arabia where full-genome sequencing of MERS-CoV isolated from a patient with fatal infection and from one of his dromedary camels proved to be positive for MERS-CoV RNA and the isolates were identical.6
Both stool and nasal secretions of dromedaries have homologous genome sequencing to MERS-CoV.2
As the virus targets the respiratory tract, nasal and mucosal secretions in humans and dromedaries eliminated through sneezing and coughing are primary spreading mechanisms in zoonotic and inter-human transmissions.7
MERS-CoV is closely associated with coronaviruses found in bats, suggesting that bats may be a reservoir of MERS-CoV.3 This prompted several researchers to screen bats in Lebanon, yet no evidence for antibodies to MERS-CoV were found.40
The majority of the CoV-positive camels existed in Saudi Arabian wholesale markets; this is where indigenous camels were mixed with imported camels from Sudan and Somalia while local camels demonstrated significantly higher rates of infection with MERS-CoVs as well as other CoVs than those imported.39 However, since its presence in the 1980s, high seropositivity against MERS-CoV among camels in the Arabian Peninsula and Africa indicates that the virus has circulated in camels prior to being introduced to the human population.35,37,39
MERS-CoV infectivity is dependent upon the median incubation period, which is approximately 5 days for limited human-to-human transmission, ranging from 2 to 14 days.
The approximate time from the onset of MERS-CoV illness to hospitalization is 4 days, with an approximate duration of 5 days for critically ill patients to intensive care unit (ICU) admission and approximately 12 days from onset of viral symptoms to death.7 Immunosuppression caused by malnutrition, deficiencies, stress, parasites, concurrent infections, etc., could aggravate the course of MERS-CoV infection.41
Phylogenetic analysis of MERS-CoV’s full-genome indicated that MERS-CoV’s evolution within camels has resulted in diverse lineages of which all have caused human infections, perpetuating a low barrier for interspecies transmission.39 Training and milking of camels have been associated with MERS-CoV infection.42
Consumption of raw camel milk or improperly cooked meat has illustrated high risk of MERS-CoV infection.1 The emergence of intensive camel livestock system and large trade between countries, particularly from Somalia to Gulf regions, leading to roughly 77,000 live camels being exported each year, can significantly contribute to multiple disease transmissions.43
Clinical manifestation and diagnosis
In terms of fatality rates, around one third of patients with MERS have died; however, as mild cases of MERS-CoV might have been missed by the current surveillance techniques, specifically in the developing countries, this ratio might be an overestimation of the actual mortality rate.1,2
In infected people, MERS-CoV causes a range of illnesses from asymptomatic to acute respiratory illness with mild to severe respiratory courses and death.1,2 Symptomatic MERS-CoV is typically introduced following an incubation period of 2 to 14 days.7
Confirmed cases include any patient sample with q-PCR positive or shown seropositivity for MERS-CoV irrespective of the presence of clinical signs and symptoms.7 It is suggested to use q-PCR for targeting the region upstream of the envelope gene (upE) as a screening assay in addition to ORF1a target gene as a confirmatory assay.44
Chest X-ray shows lung fibrosis, ground glass opacity, and pleural thickening, yet the abnormalities may not always be present in MERS CoV-infected individuals. Presentation with fever, chills, cough, shortness of breath (most likely with severe hypoxemic respiratory failure), and myalgia have been reported with MERS-CoV. Mild cases are found among children and young adults.
These include low-grade fever, runny nose, sore throat, headache, and abdominal pain.7 In critically ill patients, gastrointestinal symptoms have been observed such as nausea, vomiting, or diarrhea, while acute kidney damage has been reported in approximately half of these patients.45
Severe MERS-CoV infection is outlined by progression to acute respiratory distress, with a 2 days median period between hospitalization and ICU admission.7,45
Laboratory confirmed cases reported first signs that included fever, cough and upper respiratory tract illness, followed within a week by progressive lower respiratory tract signs and lymphopenia. MERS-CoV can progress to acute respiratory distress syndrome and multiorgan system failure. Patients commonly present to a hospital with pneumonia in addition to concurrent bacterial infections.7,46
Three cases of MERS-CoV reported onset with widespread bilateral hyperintense lesions via T2 weighted image on magnetic resonance imaging (T2WI MRI) within the white matter and subcortical regions of the frontal, temporal and parietal lobes including the basal ganglia, corpus callosum, pons, cerebellum, and upper cervical cord, suggestive of encephalitis.47
A high risk of severe course of MERS-CoV infection is seen in patients with comorbid conditions, such as diabetes, renal, cardiac and chronic pulmonary disease, cancer, as well as in immunocompromised persons. Prognostic signs of fatal courses include acute nephritis and shock.1,2,47
Treatment and prevention
No vaccination or treatment is currently available for MERS-CoV.1 A great challenge exists in developing MERS-CoV infection models for several reasons. For one, small and larger animal models that aid in initial screening do not express the DPP4 receptor.48
In addition, pharmaceutical companies have meager incentive for producing MERS-CoV vaccine as clinical trials are very costly and the timeline to approved vaccine use is 10 years or longer.49 If, however, sustained human-to-human MERS-CoV transmission were to occur, the benefit of developing a safe and effective vaccine would outweigh any cost.49
Supportive therapy necessitates respiration and circulatory support, preservation of renal and hepatic function, and prevention of secondary infections.2,50 Administration of both ribavirin and interferon-α2b may decrease viral load, as they decrease the multiplication of MERS-CoV in Vero as well as LLC-MK2 cells.2,50
It is hypothesized that DPP4 inhibitors may restrain viral replication; this yet requires further study.2 Further, the development of vaccination targeting receptor binding subdomain (RBSD) of MERS-CoV is under investigation. Recently, a candidate DNA vaccine developed from MERS-Cov spike protein subunit 1 (S1) is under study.48 As dromedaries are speculated as the source of transmission of MERS-CoV to humans particularly in the Arabian Peninsula, travelers are advised to follow basic safety precautions, including proper hand washing prior and post touching an animal, and to refrain from consuming raw meat and milk and direct contact with camels.7
More information: Kirsten A. Kulcsar et al, Comorbid diabetes results in immune dysregulation and enhanced disease severity following MERS-CoV infection, JCI Insight (2019). DOI: 10.1172/jci.insight.131774
Provided by University of Maryland School of Medicine



{kind=link}