










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")


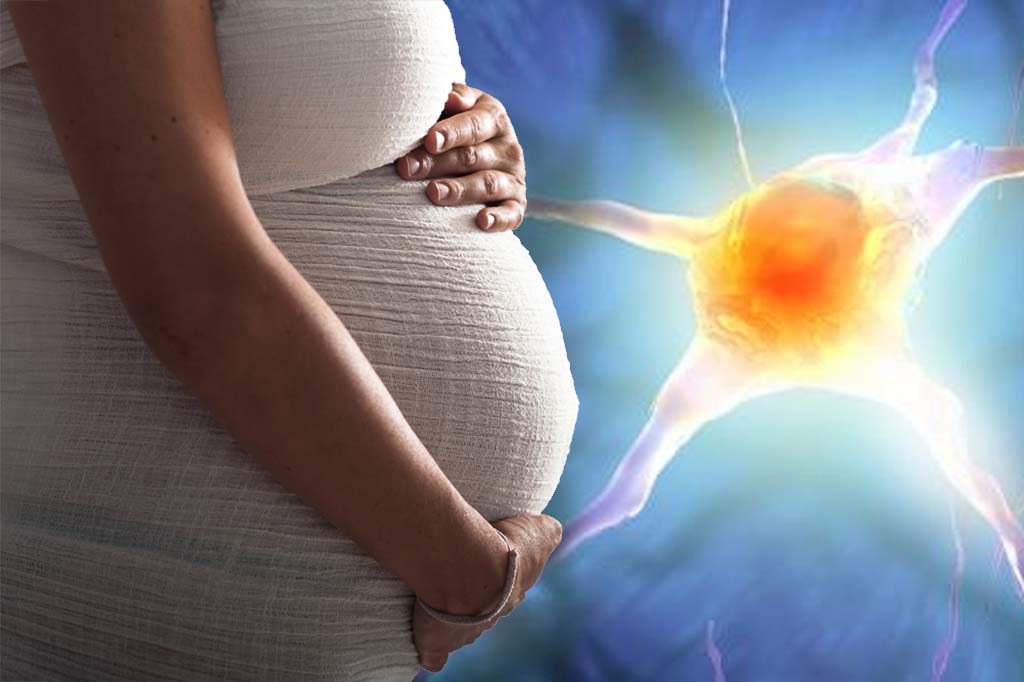
More than 30 significant risk factors have been identified for the development of psychotic disorders in offspring in research led by the NIHR Maudsley BRC.
It is the first comprehensive meta-analysis of pre- and perinatal risk factors for psychosis in nearly 20 years.
These prenatal and perinatal environmental risks, meaning risks during pregnancy and seven days after birth, have a significant effect on the likelihood of their child developing psychosis.
As a result, researchers suggest women at risk should be screened early on in their pregnancy so that those with these identified risks can be given additional support. The findings have been published in Lancet Psychiatry.
Gathering data from 152 studies published between 1977 and July 2019 and looking at 98 factors, researchers identified 30 significant risk factors and five protective factors.
Psychotic disorders are severe mental illnesses which cause abnormal thoughts, such as hallucinations or delusions, but they can affect each person in different ways.
In 2014, a survey found the 6% of people in England said they had experienced at least on symptom of psychosis.
Factors can be split into four categories; parental and familial, pregnancy, labour and delivery, and foetal growth and development.
Significant protective factors were mothers being aged between 20 – 29, first time mothers and higher birthweights in babies.
For risk factors, previous mental health conditions in either parents, nutritional deficiencies, low birthweight and giving birth in the colder months were found to increase the probability of a child developing psychosis.
Age related risk factors were either parent being under 20, mothers between 30-34 and fathers over 35.
Researchers also found that a lack of prenatal care visits poses a risk and marked this as a potential risk factor to combat with outreach campaigns.
This study will help guide future research in the field of psychosis, as well as form the basis for psychosis risk prediction models which could advance preventative strategies.
This study confirmed the importance of factors during labour and delivery, such as a fetus’ brain being deprived of oxygen and ruptured membranes, which are historically among the most consistently implicated risk factors.
Conversely, despite previous studies focusing on infections during pregnancy causing psychosis, this study found significant associations only for HSV-2 and maternal infections ‘not otherwise specified’ and found that influenza had no indication of a significant effect.
This study will help guide future research in the field of psychosis, as well as form the basis for psychosis risk prediction models which could advance preventative strategies.
Dr Paolo Fusar-Poli, Reader in Psychiatry and Youth Mental Health at Institute of Psychiatry, Psychology & Neuroscience (IoPPN), King’s College London said: ‘This study is confirming that psychotic disorders originate in the early phases of life with the accumulation of several environmental risk factors during the perinatal and prenatal phases.
The results of this study will advance our ability to detect individuals at risk of developing psychosis, predict their outcomes and eventually offer them preventive care.’
Whilst this study focused on the environmental factors there may also be genetic or epigenetic risks factors that are implicated in the onset of psychosis.
Epidemiologic studies over the past decades have repeatedly implicated prenatal environmental factors, including maternal immune activation (MIA), in the etiology of neuropsychiatric illnesses(1, 2).
Maternal infectious pathogens and inflammation are plausible risk factors for these outcomes and have been associated with schizophrenia(1–3), autism spectrum disorders(4–11) and bipolar disorder(12, 13).
Concurrent with the epidemiologic work are animal models of MIA that have documented behavioral, neurochemical, neuroanatomic, and neurophysiologic disruptions in the offspring(14–17). The use of animal models in translational research aims at complementing this work by establishing causal relationships, identifying cellular and molecular mechanisms, and exploring potential therapeutic interventions(14–17).
Despite progress on addressing these questions, there remain challenges regarding how to best approach the bidirectional “translation” between the findings of the epidemiologic and basic neuroscience studies of schizophrenia and other disorders.
The Epidemiological Perspective
A proliferation of epidemiologic studies have implicated maternal infection as a risk factor for neuropsychiatric illness(1–13). While early epidemiologic studies, which made use of ecologic data, suggested associations between influenza epidemics and schizophrenia among those exposed in utero, these findings were inconsistent.
Such studies have been supplanted in more recent years with birth cohort studies that utilize prospectively acquired serologic biomarkers of infection and/or inflammation in individual pregnancies.
As discussed in the next section, an increasing number of publications suggest associations between maternal infectious or inflammatory biomarkers and schizophrenia, autism spectrum disorders, and bipolar disorder(1, 2, 6, 14–16, 22).
Other studies, which have utilized prospective data on maternal infection acquired from records on clinically diagnosed infections, have also yielded evidence in support of these exposures as risk factors(1, 6).
The Basic Science Perspective
Motivated by epidemiologic findings, a plethora of animal models of MIA have been established over the last two decades. As extensively reviewed elsewhere(14–17), one class of animal models is based on prenatal exposure to live pathogens, such as influenza virus(23–26) or Toxoplasma gondii (T. gondii).
These models are particularly useful for the verification of causal relationships in epidemiological studies that assess the role of specific infectious pathogens. Another class of animal models makes use of immune-activating agents that primarily stimulate the innate immune system, such as the bacterial endotoxin, lipopolysaccharide (LPS), or the synthetic double-stranded RNA analog, polyriboinosinic-polyribocytidilic acid (poly(I:C)(14, 15, 17, 25, 27–29).
These were developed initially to test whether imbalances in maternal and/or fetal cytokines may be critical for the association between prenatal infection and postnatal brain pathology(30, 31). An important refinement of this second class of models was the application of individual cytokines as immune-activating agents(9, 32, 33).
This approach aims at addressing whether specific cytokines, or cytokine networks, mediate the association between MIA and neuropsychiatric illnesses. A third class of models is based on specific immunopathological processes that have been implicated in the etiology of neuropsychiatric illnesses. Two prominent examples of this class are animal models of maternal exposure to autism-related
maternal autoantibodies(34–37) and allergic disorders/asthma(38–40).
The majority of current MIA models are based on maternal exposure to non-virulent, immune-activating agents such as the viral mimetic poly(I:C)(14–17, 53). While this experimental approach offers some clear advantages(14–17, 53), it does not reproduce the full spectrum of immune responses normally induced by infectious pathogens.
For example, this method falls short in stimulating pathogen-specific humoral and cellular immune reactions, which may be part of the mechanism mediating the negative effects of maternal infection on the offspring. As discussed in detail elsewhere(53), one reason why the field shifted from MIA models that are based on exposure to infectious pathogens is that they require stringent biosafety levels, which in turn cannot be easily realized in many research laboratories.
Another reason for the increasing popularity of non-virulent immune activating agents such as poly(I:C) in MIA models is that they allow basic scientists to tightly control the intensity and duration of the (innate) maternal immune response.
This, in turn, allows researchers to identify sub- and supra-threshold effects of MIA on brain and behavioral functions in the offspring(28, 31), and whether these are influenced by the precise gestational timing of MIA(41, 42, 54–56).
Finally, MIA models using non-virulent immune activating agents were initially developed with the aim of exploring whether the association between prenatal infection and neurodevelopmental abnormalities is mediated by specific infectious processes, or alternatively, by components of immune responses that are commonly triggered by various infections, including cytokines(30, 31, 57).
Although the latter seem to be crucial contributing factors to many associations(2, 15), it is unlikely that distinct pathogens mediate the negative effects of maternal infection on the offspring through the same immune responses and pathophysiological mechanisms(58).
To fully appreciate and approach this complexity, we believe that the field would benefit from a revival and extension of experimental approaches that make use of prenatal exposure to distinct virulent and non-virulent agents.
Another limitation of MIA models is that they typically exclude “real-life” influences in humans other than the primary exposure of interest, which may mediate or modify the effects of the exposures, and play contributory roles in disease outcomes.
This limitation may also be one of the reasons why the findings from animal models of MIA appear more robust in terms of their effects on adult brain functions, as compared to epidemiological studies that explore the association between markers of infection and risk of mental disorders (see next section).
In keeping with their “artificial” nature, the findings from animal models of MIA may also be associated with the potential of providing data that are not borne out by human studies. The recent discovery that maternal segmented filamentous bacteria (SFB) mediate the association between prenatal poly(I:C)-induced immune activation and autism-related brain abnormalities in mouse offspring(59) is an illustrative example.
SFB are a family of autochthonous, apathogenic bacteria occurring in the ileum of rodents and other vertebrate species and have been shown to potently influence immune development and functions in mice(60). Contrary to mice, however, it remains controversial whether, and to what extent, SFB modulate the same immune parameters in humans(61–63).
The use of cross-species approaches in MIA models is one possible solution to minimize the potential of overinterpreting or oversimplifying the findings obtained in a certain animal species or strain (64–71).
While the majority of these models have been developed in rodent species, most notably rats and mice, some have recently been extended to species that are evolutionarily and ethologically close to humans, including rhesus monkeys(8, 64, 69, 70). Whereas rodents are separated evolutionarily from humans by more than 70 million years, rhesus monkeys diverged from human evolution approximately 25 million years ago and thus exhibit greater similarity to humans in terms of genetics, immunology, neurobiology, and behavior (71,72).
Compared with rodents, rhesus monkeys are also more comparable to humans regarding placental physiology, gestational timelines, pre- and postnatal brain development, and cortical architecture(71, 72). Thus, the inclusion of species that are more similar genetically to humans (e.g., rhesus monkeys) can aid in interpreting the outputs of rodent MIA models in terms of what they might mean for pathological symptoms in humans(65–68), thereby enhancing the cross-species transfer of information and translatability to the clinical condition in humans(71, 72)
REVIEW OF THE FINDINGS OF MIA STUDIES OF NEUROPSYCHIATRIC OUTCOMES AND RELATEDNESS OF FINDINGS BETWEEN EPIDEMIOLOGY AND BASIC SCIENCE
A key question is how the findings from these two disciplines can complement and inform one another with regard to furthering our understanding of the role of MIA in neuropsychiatric outcomes. In particular, we consider “reverse translational” approaches to this question, that is, whether human findings on maternal infection can “predict’ parallel findings in experimental model systems.
We first consider the parallels between the findings in epidemiologic and animal studies for schizophrenia, bipolar disorder, and autism spectrum disorders. Since a full review of the findings of MIA and neuropsychiatric outcomes is beyond the scope of this article, we highlight some key results and refer the reader to several comprehensive reviews(1, 2, 6, 14–17, 22). Below, we focus on the potential areas of concordance between epidemiologic and basic science studies for each of these disorders.
Schizophrenia
To date, MIA and offspring psychiatric outcomes have been most commonly investigated for schizophrenia. We focus here on select findings that are based on biomarkers of infection. Though not all findings have been replicated, key epidemiologic results include associations between maternal infectious pathogens (influenza virus, herpes simplex virus (HSV)), T. gondii, rubella, and bacterial pathogens) and inflammatory biomarkers (cytokines, C-reactive protein) and schizophrenia(1, 19).
Maternal exposure to influenza during early to mid-gestation, as quantified by antibody in maternal sera, has been associated with a threefold increased risk of schizophrenia in the Child Health and Development Study (CHDS), based on a large birth cohort in northern California(3). Elevated T. gondii IgG has been related to a twofold elevation in schizophrenia risk in this same birth cohort(73).
Maternal genital/reproductive infections have also been related to schizophrenia in this cohort(74). Maternal exposure to HSV-2 has been associated with non-affective and affective psychoses in the National Collaborative Perinatal Project(75) though not in the birth cohorts of the CHDS or the Finnish Prenatal Studies (FiPS), which is based on a large national birth cohort in Finland (76).
Neonatal antibodies to T. gondii and cytomegalovirus have been associated with non-affective psychosis in adulthood(77). In our study of maternal cytokines in the CHDS, we observed that increased interleukin-8 (IL-8) was related to schizophrenia(78).
In the FiPS, we found that maternal C-reactive protein (CRP), a non-specific biomarker of inflammation, was associated with an increased risk of schizophrenia(19). Since it is unlikely that associations between biomarkers of inflammation are accounted for by one or a small group of infections, these findings may point to a common pathogenic pathway by which different infections give rise to schizophrenia.
Since their initial establishment, animal models of MIA have repeatedly documented structural and functional phenotypes that are implicated in schizophrenia and related psychotic disorders(2, 14–17).
On the basis of early epidemiologic findings on maternal influenza and schizophrenia(1), Fatemi et al. pioneered an experimental mouse model of prenatal exposure to human influenza virus in mice(23–26, 31).
As reviewed elsewhere(14, 15), maternal influenza infection in mice led to a variety of behavioral, neurochemical, morphological, and transcriptional changes in the offspring, many of which are implicated in schizophrenia and related disorders.
These findings are thus strongly related to, and provide experimental support for, the association between maternal influenza infection and risk of schizophrenia(1, 3, 76). Since then, many additional investigations based on “reverse translational” animal models of MIA have yielded a wealth of new data supporting the predictive potential of the epidemiologic studies.
For example, deficits in sensorimotor gating, impairments in selective or sustained attention, deficiencies in working memory, and hyper-responsiveness to psychotomimetic drugs have been found in various rodent models of MIA, including prenatal exposure to influenza virus, the viral mimetic poly(I:C), the bacterial endotoxin LPS, and selected inflammatory cytokines(14–17). Some of these deficits show a maturational delay in their appearance and can be mitigated by symptomatic or preventive treatments with antipsychotic medications(14–17).
Notably, the fact that prenatal exposure to various immune-activating agents can elicit similar phenotypes is consistent with epidemiological findings suggesting that the association between MIA and schizophrenia is not limited to a single infectious or inflammatory condition(1, 15).
Despite the similarities between MIA models, however, there are also some notable differences between the models with respect to the nature of brain and behavioral changes. For example, whereas prenatal poly(I:C) exposure in rats and mice has been shown to induce cellular, neurochemical and behavioral phenotypes that are characteristic of a hyperdopaminergic state(27, 79, 80), prenatal LPS exposure may rather induce a hypodopaminergic state in adult rodent offspring(81).
Prenatal LPS exposure in the rhesus monkey was also found to cause a significant increase in global white matter volume(64), whereas an opposite pattern (i.e., decreased white matter volume) was observed in rhesus monkey offspring born to influenza-infected mothers(69).
Besides the notable influence of prenatal timing and the genetic background discussed above, such differences may arise because different immunogens can induce a distinct set of neuroimmune abnormalities across brain development, and consequently, may lead to differing long-term deficits in brain structure and function.
This notion would also be consistent with epidemiological findings that appear to suggest that not all infectious pathogens have the same potential to increase neuropsychiatric disease risk(1, 13). As discussed more extensively below, a closer examination of the commonalities and differences between the mediating factors and outcomes of distinct MIA models should help to further address this important issue.
Another question is whether animal models can also “predict” certain epidemiological associations. While comparatively little work has been conducted, our recent findings support this assertion.
We developed an environmental “two-hit” model in mice, in which prenatal exposure to mild but physiologically relevant MIA served as the “first hit”, and subchronic exposure to unpredictable, psychological stressors in pubescence as the “second hit”(50).
Hence, this multifactorial model incorporates two environmental risk factors that have each been associated with increased risk of psychiatric disorders such as schizophrenia. We showed that combined exposure to the two environmental adversities acted in synergy to induce psychosis-related neural and behavioral abnormalities in adult mice(50).
These results provided the first evidence suggesting that prenatal immune adversities can function as a neurodevelopmental disease primer, which in turn can increase the offspring’s vulnerability to the detrimental neuropathological effects of subsequent stress exposure during pubescence(50).
These basic-science findings have recently been translated to a large population-based epidemiological study, which comprised nearly 1 million Danish persons born between 1980 and 1998(82). In that study, Danish nationwide registers were linked to estimate the independent and joint effects of exposure to prenatal infection and peripubertal psychological trauma on the risk of schizophrenia(82).
Prenatal exposure to infection was defined based on hospital admissions with an infection during pregnancy, whereas exposure to traumatizing experiences during the period of peripuberty (from age 8 to 14 years) was defined according to Danish standards and included parental deaths, maltreatment or physical and/or sexual abuse, and maternal and paternal histories of crime and occupational situations(82).
Confirming the hypothesis initially put forward by the environmental “two-hit” model in mice(50), the Danish population-based epidemiological study demonstrated that exposure to prenatal infection and peripubertal psychological trauma was associated with a significantly higher risk of developing schizophrenia (in males) compared to exposure to either insult alone, and the interaction between infection and trauma attained statistical significance(82).
These findings suggest that the cross-fertilization between basic research in animals and risk factor epidemiology may offer the potential of predicting yet undiscovered associations between MIA and neuropsychiatric illnesses.
Bipolar disorder
Thus far, only a few epidemiologic studies have evaluated MIA in relation to bipolar disorder in offspring. Our group has demonstrated that maternal influenza, documented by antibodies in prenatal sera(19) and physician diagnoses(83), has been associated with a fivefold increased risk of bipolar disorder.
While most other studies suggest no association between maternal infectious pathogens and bipolar disorder(13), one study found that maternal exposure to the type I strain of T. gondii was related to an increased risk of affective psychoses in offspring, which includes bipolar disorder(84).
Even though animal models of MIA have not specifically explored their validity for bipolar disorder, some of the experimentally induced phenotypes may be relevant for this neuropsychiatric illness as well. For example, deficits in sensorimotor gating, as seen in various rodent MIA models(2, 14–17), are also present in acutely manic(85) and remitted bipolar disorder patients(86).
Moreover, several animal studies have reported the emergence of depression-like behaviors in offspring exposed to MIA(87, 88). The latter phenotypes may not only be relevant for unipolar depression, but also for depressive episodes in bipolar disorder. The investigation of other core behavioral symptoms of bipolar disorder, such as poor decision-making, altered risk-taking behavior, impulsivity, and loss of inhibitory control remain unexplored in MIA models.
Additional work is also necessary to evaluate whether MIA-induced deficits can be mitigated by pharmacological treatments used in bipolar disorder, including the mood stabilizer lithium, and anticonvulsants such as valproate and lamotrigine(89).
Autism
In recent years, maternal infection and inflammation have been investigated in relation to autism spectrum disorders. Although findings are mixed, and more work is necessary, evidence has emerged linking maternal inflammation to risk of autism spectrum disorder in offspring.
In the FiPS birth cohort, our group demonstrated that elevated maternal levels of CRP, a nonspecific biomarker of inflammation, in early to mid-gestation, was related to an increased risk of autism spectrum disorders in offspring(4).
However, in the Early Markers of Autism (EMA) study in California, maternal mid-pregnancy CRP levels were related to a decreased risk of autism spectrum disorders(90). In studies of cytokines and chemokines in archived maternal serum samples in the EMA study, significantly increased levels of these analytes were related to autism spectrum disorders(5, 11). In amniotic fluid samples from a Danish study, several cytokines including tumor necrosis factor-alpha (TNFα) and several inflammatory interleukins were related to autism spectrum disorders in offspring(7).
Moreover, maternal fever has been associated with autism. Although replication of these findings is necessary, they suggest that MIA may also be related to autism spectrum disorders. Consistent with this interpretation, other maternal immune factors, including maternal autoantibodies targeting fetal proteins, have been associated with increased autism spectrum disorder risk in the offspring (for review see (91)).
These findings include significant associations between paired maternal antibody reactivity to fetal brain proteins with the 37 and 73kDa molecular weight bands and diagnosis of ASD in children(92). Within proteins corresponding to the 37-, 39-, and 73 kDa bands, maternal autoantibodies recognized seven developmentally regulated proteins in the fetal brain, including lactate dehydrogenase A and B, stress-induced phosphoprotein 1, and collapsin response mediator proteins 1 and 2(91, 92).
Several of these proteins are critical for normal brain development, including neuronal migration and neural network formation.
Animal models further support the hypothesis that MIA is an environmental risk factor for autism spectrum disorders. For example, prenatal exposure to the viral mimetic poly(I:C), the bacterial endotoxin LPS, or allergies/asthma, can all induce behavioral abnormalities that are reminiscent of core symptoms of autism spectrum disorders, including deficits in social interaction and communication as well as high levels of repetitive behaviors(9, 15, 38, 93).
These manipulations also cause brain morphological and cellular abnormalities implicated in autism spectrum disorders, including abnormal cerebellar development, impaired expression of the extracellular matrix protein reelin, and altered synapse density and neural connectivity(2, 15, 25). Importantly, some of these rodent findings have been extended to rhesus monkeys, both at the behavioral and brain morphological levels(8, 64, 70, 94).



{kind=link}