











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Developing safe and efficient bioformulations using approved materials and ingenious designs can accelerate the clinical translation process.
Scientists from the Institute of Process Engineering (IPE) of the Chinese Academy of Sciences have developed a new therapeutic tumor vaccine based on self-healing polylactic acid microcapsules, which can efficiently activate the immune system and inhibit tumor development.
This research was published in Science Advances on May 22.
Therapeutic cancer vaccines that harness the immune system to reject cancer cells have shown great promise for tumor treatment.

The research team, led by Prof. Ma Guanghui and Prof. Wei Wei from IPE, already designed and fabricated a variety of tumor vaccines in their previous work. Theses vaccines have been proven effective in different tumor models, such as lymphoma, melanoma and breast cancer.
The researchers were impelled to improve the earlier tumor vaccines, however, due to certain limitations. For example, Prof. Ma said that an unfavorable immunization microenvironment, along with a complicated preparation process and the need for frequent vaccinations significantly compromised their performance.
“Therefore, we designed a novel microcapsule-based formulation for high-performance cancer vaccinations,” said Prof. Ma.
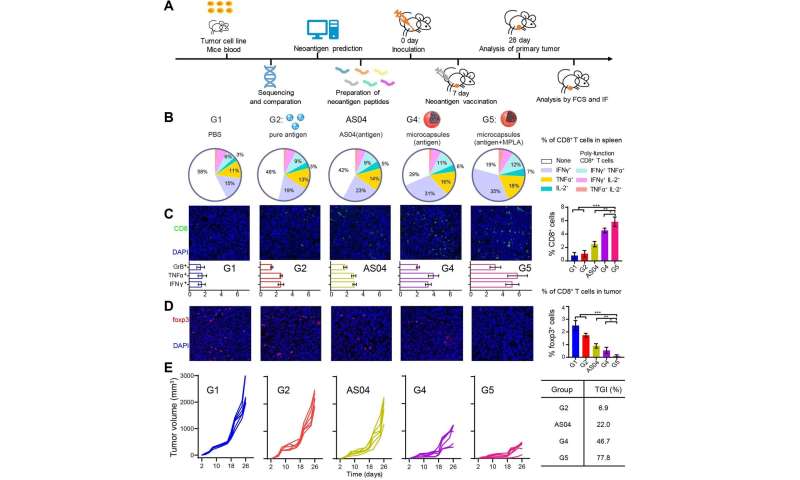
This study represents the first time researchers used self-healing microcapsules with post-encapsulation, multiple loading, and efficient modulation of immunization microenvironments in a tumor vaccine.
The special self-healing feature provides a mild and efficient paradigm for antigen microencapsulation. After vaccination, these microcapsules create a favorable immunization microenvironment in situ, wherein antigen release kinetics, recruited cell behavior and acid surrounding environment work in a synergetic manner.
Owing to synergetic effects, the vaccine succeeds in increasing antigen utilization, improving antigen presentation and activating antigen presenting cells. “As a result, effective T cell response, potent tumor inhibition, anti-metastatic effects and prevention of postsurgical recurrence are achieved with various types of antigens in different tumor models,” said Prof. Wei.
Moreover, the researchers verified the availability of the novel vaccine platform used in the neoantigen vaccine, which conforms to precision medicine, said Prof. Wei. Due to the simple post-encapsulation process, the clinicians were able to prepare the neoantigen formulation by themselves at any time.
A peer reviewer from Science Advances described the study as “comprehensive and rationally designed.” The reviewer also emphasized that the results are “impressive” and the work has “high value for therapeutic vaccines and cancer immunotherapy.”
Our immune system comprises a complex network of cells, tissues, and organs that work in harmony to protect the body against deadly diseases. This immune system attacks and eliminates foreign invading particles with exquisite specificity.
Diseases are caused by malfunctioning or underperforming immune response; an over-reactive immune system can cause autoimmunity, which may lead to the destruction of healthy tissue [1,2], and an underactive immune system can make our body more susceptible to infection [3].
Vaccines consist of a biological agent that resembles a disease-causing microorganism and improves immunity against that particular disease. They develop immunity that can control and adjust unbalanced immune systems that are either overreactive or underactive [4,5,6]. Weiner et al. described the first therapeutic vaccine against autoimmunity [7]. The development of vaccines has historically been based on Louis Pasteur’s “isolate, inactivate, inject” paradigm [8].
Currently, vaccines are considered to be one of the most effective tools for the prevention of infectious diseases. Thus, vaccine developments against bacterial infections, viral infections, and cancer are considered to be significant milestones in the field of medicine [9]. In the past, traditional vaccines made from pathogens in either killed or inactivated forms were considered efficient [8,10].
Vaccines with live-attenuated pathogens make use of the weakened type of microbe to create a stronger and enduring immune response. However, a significant concern arises when the weak pathogen might revert to its active form, causing severe disease condition. The mutagenic actions inside the infected host organism could generate more virulent strains.
In addition, pathogens in their inactive or killed forms cannot revert to active forms, and tend to stimulate weaker immune responses, which in turn requires multiple administrations of doses, hence limiting its practical use.
Usually, vaccines are comprised of an antigen, which acts as the target for causing immune response, and an adjuvant, which is co-administered with the antigen to enhance the immune response. A
luminum was the first-ever adjuvant, and was used primarily to increase antibody production, making it a suitable candidate for vaccine formulation [11].
However, aluminum adjuvants fail to generate strong cell-mediated immunity, and carry the risk of autoimmunity, and long-term brain inflammation, causing adverse health issues.
Adjuvants have been reported to cause both local as well as systemic toxicity. Certain adjuvants like Freund’s incomplete adjuvant, Quil A, induce local toxicity, whereas adjuvants based on pathogen associated molecular patterns like Aluminum adjuvants induce systemic toxicity [12].
Another adjuvant was Freund’s incomplete adjuvant in the form of mineral oil-in-water emulsion that contained heat-killed mycobacteria; it was found to be reactogenic in humans [13].
Although these adjuvants could induce local immune reactions, they failed to generate strong cell-mediated immunity, which demanded the development of new adjuvants for successful vaccine delivery.
According to a web-based central database pertaining to vaccine adjuvants, nearly a hundred vaccine adjuvants have been used in various vaccines against different pathogens, of which very few have received licenses for human use [14].
Recently, nanoparticles (NPs) have gained enormous attention as delivery vehicles for vaccines. Nanovaccine formulations not only provide enhanced antigen stability and immunogenicity, but also offer targeted delivery and prolonged release.
A high number of NP vaccines with varied physicochemical characteristics and properties have been approved for clinical use [15,16,17]. The primary purpose of the use of nano- and microparticle-based delivery systems is to enhance the duration of antigen presentation and dendritic cell (DC)-mediated antigen uptake, which result in the direct stimulation of DCs, and promote cross-presentation [15,16,18].
Furthermore, NPs help in protecting the antigen and adjuvant from premature enzymatic and proteolytic degradation [19]. Vaccine antigens can be delivered to the target site by either encapsulating them inside an NP, or by decorating them onto the surface of NPs.
The NP delivery systems can load multiple components in a single carrier, which enable a prolonged, simultaneous, and targeted delivery of antigens [20,21], adjuvants [22,23], DNA plasmids [24], and detained bacterial toxins [25].
The development of vaccine candidates is based on several factors, such as minimalist compositions, low immunogenicity, and formulations that boost antigen effectiveness [26,27,28,29].
Owing to their unique physicochemical characteristics, such as large surface area-to-volume ratio, controllable size and shape with different surface charge, NPs can be surface-engineered with peptides, proteins, polymers, cell-penetrating peptides, and other targeting ligands, which make them a versatile delivery vehicle for vaccine formulations.
Design of NPs based vaccines can assist for multimodal imaging to improve therapeutic level by visualizing the vaccine inside our body [30,31,32,33]. Although NPs have the abovementioned advantages, they have disadvantages, in that they lack colloidal stability in physiological conditions due to protein corona formations, and have undesirable interaction with the reticuloendothelial system (RES) [34,35].
Biomimetic NPs are a novel class of NPs that exhibit enhanced colloidal stability, while efficiently avoiding unwanted interaction with immune cells like RES, and prolonging circulation in the blood [36,37,38].
These nanovaccines involve carrier NPs that mimic biological membranes, and when administered in the body, achieve prolonged circulation and evasion of immune responses [39]. Among biomimetic NPs, liposomes are obtained by the dispersion of phospholipids in water, and have a high loading capacity, with the ability to co-deliver both hydrophobic and hydrophilic drugs [40].
Cell-membrane coated NPs are another type of biomimetic nanocarrier with a “core–shell” structure, in which the NP forms the hydrophobic core, and a thin layer of plasma membrane coating acts as the shell.
In this NP, various cell membranes are used to cloak synthetic NPs through a top-down fabrication method, thus preserving the physicochemical properties of the core synthetic NPs, while maintaining the cellular composition on its hydrophilic membrane shell [41].
Hu et al. [42] reported the first membrane-coated NPs where red blood cell (RBC) membranes were coated over a polymeric NP through extrusion [42]. Many types of membranes from different sources, such as RBCs [41,42,43], leukocytes [44,45,46], cytotoxic T-cells [47], NK cells [48], platelets [49], macrophages [44,50], and cancer cells [51,52], have been used in the preparation of membrane-coated NPs.
Another type of biomimetic nanovaccine can be self-assembling proteins, which are known to have high symmetry and stability, and can be structurally organized into particles of sizes (10–150) nm [53,54].
These self-assembling protein NPs play diverse physiological roles, and are selected as vaccine carriers owing to their ability to self-assemble and deploy into a definite structure that mimics a natural microbe architecture [55]. Virus-like particles (VLPs) are another type of biomimetic nanovaccine that contain noninfectious subsets of virus that lack genetic materials; they assemble without containing any viral RNA [56].
In this review, we discuss the recent advances in biomimetic nanovaccines and their applications in anti-bacterial therapy, anti-HIV therapy, anti-malarial therapy, anti-melittin therapy, and anti-tumor immunity.
Components of Biomimetic Immunomodulatory Nanovaccines
Biomimetic nanovaccines include a biomimetic carrier that is loaded with therapeutic molecules that are designed to deliver to the target site. Figure 1 shows that the various types of biomimetic NPs involve liposomes, protein NPs, cell-membrane decorated NPs, and VLPs.

Schematic of different formulations of biomimetic nanovaccines and their advantages. a) Biomimetic nanovaccines maintain the germinal center and B-cells inside our body, which are responsible for the release of antiviral neutralizing antibody against viruses, b) biomimetic nanovaccines strengthen the humoral immune response by inducing higher DC maturation and stimulating cytotoxic T-cell to kill cancer cells, c) biomimetic nanovaccines can target infected blood cells, and induce a strong immune response inside our body, and d) biomimetic nanovaccines are suitable candidates for carrying antigens, adjuvants, and therapeutic molecules. (MPLA: monophosphoryl lipid A, STING: stimulator of interferon gene, POLY (I:C): polyinosinic:polycytidylic acid, MPER: membrane-proximal external region, HER-2: human epidermal growth factor receptor 2, OVA: ovalbumin and MSP: merozoite surface protein).
Types of Biomimetic Nanoparticles (Nps)
Table 1 describes the different type of biomimetic nanovaccines that are reported so far along with its applications.
Table 1 – Already reported biomimetic nanovaccines and its applications.
| Nanoparticles | Components | Application | References |
|---|---|---|---|
| Liposomes | Liposome-polycation-DNA NPs | DNA vaccine delivery | [24] |
| PLGA NPs with lipid antigens | Malarial vaccine delivery | [21] | |
| Cancer cell membranes with lipids coated onto polymeric NPs | TLR 7 delivery: Anticancer vaccine | [52] | |
| VLPs | Avian retrovirus with Gag fusion proteins | Intracellular protein delivery | [57] |
| Genetically modified VLP | Anti-viral protection | [58] | |
| Self-assembling proteins | Hollow vault protein | Suppress lung cancer proliferation | [59] |
| Cell membrane decorated NPs | Gastric epithelial cell membrane coated PLGA NPs loaded with antibiotics | Anti-bacerial therapy | [60] |
| Bacterial membrane coated Gold NPs | Antibacterial immunity | [61] |
Liposomes
Liposomes are biomimetic products that are formed by dispersing phospholipids in water [58,62,63]. They occur as either unilamellar vesicles with a single phospholipid bilayer, or as multilamellar vesicles with several concentric phospholipid shells separated by different layers of water. Liposomes can be modified to incorporate both hydrophobic and hydrophilic molecules into the phospholipid bilayer and aqueous core [64].
Liposomes can be used to encapsulate antigens within their core for delivery. They form virosomes when viral envelope glycoproteins are incorporated into their base [65,66]. Influenza virus was the primary focus for virosome studies which has been established for industrial application as human vaccine [67].
Five vaccines based on virosome are under clinical trials, and four virosome vaccines are approved for commercial application in various diseases [67]. One of the commonly used NPs for adjuvant delivery in DNA vaccines is liposome-polycation-DNA NPs; they are formed by the combination of cationic liposomes and cationic polymer-condensed DNA.
Liposome-polycation-DNA assembles to form a nanostructure, with the condensed DNA located inside the liposome with a size of 150 nm [24,68]. Moon et al. [21] reported the development of a malaria vaccine, which could be used for the delivery of polymeric PLGA NPs enveloped with lipid antigens.
In their work, Moon and coworkers developed a pathogen-mimicking nanovaccine, in which the candidate malarial antigen was conjugated to the lipid membrane and incorporated with an immunostimulatory molecule, monophosphoryl lipid A-MPLA, and further used to elicit immune responses against P. vivax sporozoites [21].
Yang et al. [52] used cancer cell membranes which were modified with lipids using the lipid-anchoring method, and then further coated them over polymeric NPs with a toll-like receptor 7 (TLR 7). This biomimetic membrane nanocarrier was reported for use as an anticancer vaccine, as well as for the delivery of TLR 7 as an adjuvant [52].
Virus-Like Particles (VLPs)
VLPs are molecules that resemble the structure of viruses without viral genetic material. These self-assembling NPs that lack infectious nucleic acid are formed by the self-assembly of biocompatible capsid proteins.
They are ideal nanovaccine systems, as they have the innate viral structure, which can interact with the immune system without any threat of causing infections [69,70]. These VLPs can act as vaccines have nano-size and a repetitive structural order, and could induce an immune response in the absence of an adjuvant [71].
VLPs assemble without encapsulating any viral RNA, and hence they are noninfectious and nonreplicating, as the genes coded for viral integrase are deleted before expression. This prevents packed genome integration into the host cell, as well as the recombination of the live or defective virus.
The first VLP vaccine was developed against the hepatitis B virus, which was later commercialized in 1986 [72]. VLP vaccines against hepatitis E and the human papillomavirus have been used in human since 2006 [73,74].
VLPs can be obtained from a variety of viruses, and can have different sizes ranging (20 to 800) nm; further, they can be obtained via different processes [56]. The initial approach to obtain VLPs involves the self-assembly of capsid proteins in the expression host, followed by purification of the assembled protein to avoid contaminants that are adhered or encapsulated.
However, in a few cases, for better quality and low contamination, the VLP structure needs to be disassembled and reassembled. Another emerging method to obtain VLPs is to use cell-free in vitro processing, wherein at first large-scale purification is performed to prevent contamination, and then assembly of VLP structures in vitro, to avoid their disassembly in a cell; commercialized VLPs are derived from a target virus by self-assembling its proteins.
For VLP to be used as a delivery vehicle, the target antigen from a virus different from the one used in the VLP is attached to the VLP surface; and this surface-modified VLP paves the way for its use in targeting various diseases. VLPs could be engineered to attach additional proteins on its surface, either through the fusion of proteins on the particle, or by expressing multiple antigens, which in turn protects against its source virus and other antigens present on its surface [75].
Polysaccharides and small organic molecules are non-protein antigens that can be chemically attached to the VLP surface to form bioconjugate particles [76]. The baculovirus expression system is mostly used to generate VLPs with an excellent safety profile, as baculovirus does not naturally infect human [77].
In another study, a safe and efficient VLP system based on avian retrovirus was designed such that the system was considered safe, as it could not replicate itself in human cells.
This system was considered as safe because the VLP constitutes only Gag fusion protein; a single VLP could deliver about (2000–5000) copies of the Gag fusion protein into the transduced cell. In another study, VLPs were created for delivery with two different approaches: the intracellular distribution of Gag fusion proteins, or by modifying the surface of VLPs for receptor/ligand-mediated delivery (Figure 2) [57].

Schematics of the generation of functional virus-like particle (VLP) and (A) the delivery of proteins of interest intracellularly, and (B) by receptor/ligand-mediated protein delivery. Reproduced with permission from Ref. [57]; Copyright © 2011, National Academy of Sciences.
Self-assembling Protein Nanoparticles (NPs)
Many naturally occurring proteins can self-assemble to form NPs with high symmetry and stability, and these NPs are structurally organized to form particles that range in size (10–150) nm [53,54]. These NPs with diverse physiological roles are selected as vaccine carriers, owing to their ability to self-assemble and deploy into a definite structure that mimics a natural microbe architecture [55].
Ferritin is a protein that protects cells from damage caused by Fenton reactions, in which iron catalyzes hydrogen peroxide, and converts it into highly toxic hydroxyl radical. Under oxidizing conditions, harmful reactive oxygen species are produced from free Fe (II), which can damage cellular machinery [78].
Ferritin has a hollow structure, and the ability to store iron within this hollow cavity; thus, it acts as a storage system for iron [79]. Ferritin can self-assemble into spherical nanostructures and be used to fuse with the influenza virus haemagglutinin (HA) genetically, and the recombined protein spontaneously assembles into a particle of octahedral symmetry.
This reforms into eight trimeric HA spikes, and elicits a stronger immune response, compared to an inactivated trivalent influenza virus [80]. Another type of self-assembling protein is the major vault protein (MVP). Champion et al. [81] reported that 96 units of the MVP self-assemble to form a barrel-shaped vault NP of length 70 nm and width 40 nm.
Further, they mentioned that genetically fused antigens that have minimal interactions could be loaded onto vault NPs that had self-assembled through mixing with MVPs. In their work, they encapsulated an immunogenic protein termed the major outer membrane protein of Chlamydia muridarum into hollow vault nanocapsules.
These hollow vault nanocapsules were modified to bind IgG for an enhanced immune response, to induce protective immunity at distant mucosal surfaces [81]. Wahome et al. reported another self-assembling protein NP, an adjuvant-free immunogen [82] obtained by the self-assembly of a monomeric chain into an ordered oligomeric form as an antigen-presenting system that could be suitable for vaccines.
This self-assembling protein NP was formed by incorporating the membrane-proximal external region (MPER) of HIV-1 gp41, which is identified as a target for a wide range of neutralizing antibodies, in the N-terminal pentamer, to produce an α-helical state of the 4E10 epitope, without causing structural changes in 2F5 epitopes.
These self-assembled NPs showed enhanced membrane-proximal region-specific titers, owing to the presence of a repetitive antigen display of MPER even without any adjuvant, thus resulting in the formation of an adjuvant-free immunogen as a potential HIV vaccine [82].
Cell Membrane-Decorated Nanoparticles (NPs)
As discussed in the previous section, cell membrane decorated NP has emerged as a promising method for camouflage by forming a thin layer of the cell membrane coating over the NPs. The camouflaged NPs inherit the properties of the source cells, depending on the source cells used.
For example, when RBCs are employed as the source membrane, membrane-coated NPs are found to possess immune evasion and prolonged circulation [42]. Biomimetic NPs attain these cell mimicking properties by the transference of the source cell’s membrane proteins onto the surface of NPs [39].
This functionalization approach is regarded as highly versatile, allowing the delivery of a wide range of cargoes that encompass various inner-core materials.
Targeted drug delivery employs the inherent adhering capability of source cells. For example, NPs camouflaged with a layer of cancer cell membranes showed inherited homotypic adhesion properties, and an intrinsic capacity to bind with the source cells [20,51].
In addition, NPs camouflaged with platelet membranes displayed the ability to mimic platelet binding with pathogens, such as methicillin-resistant Staphylococcus aureus, for targeted antibiotic delivery. Meanwhile, platelets help in recognizing tumor cells, including circulating tumor cells, through their ligand binding interactions.
Platelet membrane-camouflaged NPs were primarily formulated for the site-specific delivery of anticancer drugs. These persuasive applications inspired the development of cell membrane-camouflaged NPs for targeted antibiotic delivery against the H. pylori infection. Angsantikul et al. [60] reported a nanotherapeutic that was obtained by coating antibiotic-loaded poly(lactic-co-glycolic acid) (PLGA NPs) with a gastric epithelial cell membrane against an H. pylori infection.
In their study, it was found that the gastric epithelial cellular membrane-coated NP had the same surface antigens as the source cells that exhibit inherent adhesion towards H. pylori bacteria [60].
The use of bacterial membranes as vaccination materials has gained considerable interest. They can stimulate innate immunity and promote adaptive immune responses by exhibiting different pathogen associated-molecular patterns (PAMPs) for a large number of immunogenic antigens with adjuvant properties [83].
Camouflaging NPs with covering bacterial membranes results in the preservation of bacterial characteristics, and thus helps in mimicking natural antigen presentation by bacteria to the immune system. Gao et al. [83] reported a bacterial membrane coated NP for antibacterial therapy, in which gold NPs were coated with bacterial outer vesicles.
In this study, they chose E. coli bacteria, obtained its outer membranes, and coated gold NPs of 30 nm size with them; they found that this could induce rapid activation and DC maturation in the lymph nodes. Further, vaccination with these NPs produced long-lasting and robust antibody responses [83].
Exosomes
Exosomes are nanosized membrane-enclosed extracellular vesicles originated from the inner endosomal membrane. These vesicles are composed of a lipophilic bilayer with proteins and genetic materials such as micro RNAs, mRNAs, and DNAs [84].
Exosomes are the mediator between cells and can induce immune response by activating natural killer (NK) cells, dendritic cells (DC), and T lymphocytes cells [85]. Various physiological stimuli such as inflammation, oxidative stress and cell growth affects the secretion of exosomes from the cells which is used as a prominent diagnosis marker [86].
Exosomes are acts as vaccination against infection. It can be used as the carrier of pathogen antigens to by modulating the immune response and recruiting monocytes, macrophages, NK cells, and T cells against the infectious agents [87].
Cargoes Used for Immunomodulatory Nanovaccines
As mentioned in the earlier Section, biomimetic immunomodulatory nanovaccines are composed of 1) biomimetic NPs, and 2) the cargoes used. In this section, the different types of cargoes used for nanovaccines are explained.
Adjuvants
Adjuvants are ingredients used in vaccines to enable the body to produce a stronger immune response, and help vaccines work better. There are different mechanisms by which adjuvants elicit immune responses, which are as follows:
1) prolonged release of antigen at the site of injection,
2) cytokines and chemokine level gets upregulated,
3) recruitment of cells at the injection site,
4) antigen uptake and presentation to antigen-presenting cells increases,
5) APC activates and matures, resulting in the migration to draining lymph nodes, and
6) inflammasome activation [88,89,90].
Generally, adjuvants are classified based on their mechanism of action, physicochemical properties, and origin. Adjuvants can be classified as delivery systems or immune potentiators, depending on their action mechanism. Table 2 describes the partial list of adjuvants used in the abovementioned three categories.
Table 2 – Few types of adjuvants used and their classification.
| Immune Potentiators | Delivery Systems |
|---|---|
| dsRNA: Poly (I:C), Poly-IC:LC MPLA (monophosphoryl lipid A) LPS (Lipopolysaccharide) CpG oligodeoxynucleotides Flagellin Imiquimod (R837) Resiquimod (848) Saponins (QS-21) | Aluminum salts Incomplete Freund’s reagents Virus-like particles Polylactic ac |
Champion et al. reported a vault NP vaccine for inducing protective immunity at distant mucosal surfaces. These vault NPs contain immunogenic proteins, and hence they are considered as adjuvants [81].
In another study, Riitho et al. [91] formulated a biomimetic vaccine by encapsulating a viral protein inside a polymeric shell, wherein the viral protein was known to have effective cross-presentation by MHC class I.
These polymeric NPs were adjuvanted with polyinosinic: polycytidylic acid (poly(I:C)), and loaded with viral proteins that act as antigens. These nanovaccines exhibited significant virus-neutralizing activity, and they were effective against infections caused by the bovine virus diarrhea-virus [91]. Wang et al. [22] reported the use of a dual-functional nanomodulator to enhance CpG mediated cancer therapy. In their work, they synthesized manganese oxide nanosheets and conjugated anticancer drug doxorubicin (DOX) and CpG-silver nanoclusters as the adjuvant [22]. Yang et al. [52] reported the use of a lipid (DSPE-PEG-mannose) modified cancer cell membrane that was coated onto a polymeric NP loaded with adjuvant TLR 7 for an anticancer effect [52].
Recently, Le et al. [92] suggested an in situ nanoadjuvant as a tumor vaccine to prevent the long-term recurrence of tumors. In their study, polydopamine NPs were loaded with imiquimod, and then the NP surface was modified with programmed death-ligand 1 (PDL1) antibodies for the co-delivery of both antigen and adjuvants to the same antigen-presenting cells. This nanoadjuvant with PDL1 antibody could block PDL1 immune checkpoint in tumors, and it is expected to have combinational photothermal and immunotherapy effects [92].
Moon et al. reported the development of a recombinant antigen derived from the circumsporozoite protein, which is the most predominant membrane protein on sporozoites. Their initial work stated that this recombinant antigen, when mixed with conventional antigens could elicit an antigen-specific antibody response [21].
They used a lipid enveloped polymeric NP, and conjugated the malarial antigen into the lipid membrane with an immunostimulatory molecule monophosphoryl lipid A incorporated into the lipid membranes, which resulted in a pathogen-mimicking NP vaccine [21]. Another study suggested that antigen-loaded NPs that display monophosphoryl lipid A (MPLA) and further encapsulation with adjuvant CpG motifs and model antigen Ovalbumin could act as an efficient bacterial vaccine [93].
In that study, CpG potency was found to be enhanced when it was encapsulated inside the NP, which in turn highlights the importance of the biomimetic presentation of pathogen-associated molecular patterns. Because of MPLA with CpG, the pro-inflammatory, antigen-specific T helper 1 (Th 1) cellular and antibody-mediated immune responses were significantly increased [93]. Sahu et al. [23] reported the use of monophosphoryl lipid A (MPLA) NPs loaded with a Hepatitis B surface antigen (HBsAg) for delivery in the colon, which provided prolonged immunization against the Hepatitis B infection.
In this study, MPLA was the adjuvant; it activated toll-like receptor type 4 (TLR 4) and Hbs Ag that act as the antigens to be delivered, and thus enabled the simultaneous delivery of both adjuvant and antigens inside the colon.
The results indicated that it was effective in the generation of humoral and cellular immune responses [23]. Stimulator of interferon gene (STING) is a prominent agonist which stimulates cyclic dinucleotides (CDNs) to activate IRF3 and NFκB pathways and secrete various pro-inflammatory cytokines. Jack Hu et al. had developed pH sensitive capsid-like hollow polymeric nanoparticle loaded with STING agonist, cyclic diguanylate monophosphate (cdGMP), as a Middle East respiratory syndrome coronavirus (MERS-CoV) vaccine.
Delivery of both STING agonist and MERS-CoV receptor binding domain antigen in the surface of the nanoparticle mimicked as virus-liked nanoparticle and induced Th1 type immune response which is a prominent vaccine against the infection [94].
Detained Bacterial Toxins
A toxoid is a chemically or physically modified toxin that is no longer harmful but retains immunogenicity. Wang et al. [20] developed a nanotoxoid that consists of RBC membrane-coated polymeric NPs, and the membrane coating acts as a substrate for the pore-forming staphylococcal α-hemolysin (Hla) nanotoxoid, thereby effectively triggering the formation of germinal centers, and inducing high anti-Hla titers.
Further, the nanotoxoid formed showed superior protective immunity against methicillin-resistant Staphylococcus aureus (MRSA) skin infection (Figure 3) [20].

Scheme of biomimetic nanotoxoid showing protection against methicillin-resistant Staphylococcus aureus (MRSA)-induced skin infection. a) The normal condition of skin lesion formation in which MRSA bacteria employs hemolysin (Hla) and helps in colonizing the site. b) After nanotoxoid vaccination, anti-Hla and neutralize the toxins produced by MRSA. Reproduced with permissions from Ref. [20], Copyright © 2016, John Wiley and sons.
Recently, Wei et al. [44] reported a macrophage-membrane-coated nanotoxoid against pathogenic Pseudomonas aeruginosa. It has already been reported previously that alveolar macrophages have cationic proteins that can bind to the outer membrane of the bacteria Pseudomonas aeruginosa, and its flagella also get involved in phagocytosis.
Advantages of Nanovaccines
Biomimetic nanovaccines are ideal vaccine candidates, as they have unique physicochemical parameters, such as size, shape, and biomimicking property. This feature makes them a versatile delivery system for the delivery of antigens and adjuvants.
The main advantage of nanovaccines is their ability to incorporate both antigens and adjuvants within a single particle to produce maximum stimulation. The biomimicking property of these nanovaccines reduces interactions with RES cells, provides longer circulations, and prevents the burst release of adjuvants from its nano-formulation.
The synthesis methods and the choice of material used for NP formulations make the nanovaccine flexible, so that it can incorporate different molecules, such as proteins, polysaccharides, lipids, polymers, and nucleic acids.
The NP localization can be enhanced by modifying the NP surface with ligands that have specificity to immune cell receptors [95]. Moreover, antigens and adjuvants can be loaded into NPs either individually or for a combinatorial approach, and protect its molecule integrity from different enzymes, such as nucleases and phosphatases [96].
Besides these advantages, NP formulation also prevents adjuvants from degradation, protects the body from potential systemic toxicity caused by the premature release of adjuvants, and enhances immune response through extended cargo release [19].
Another advantage of biomimetic nanovaccines is their ability to target immune cells; because they are of nanosize, the nanovaccines drain into the lymphatic system, allowing for efficient delivery to lymph nodes, where immune cell density is high [97].
The selection of biomimetic NP plays an essential role in improving vaccine efficiency. Biomimetic nanovaccines helps in shielding the NPs to be recognized from mononuclear phagocytic system and helps in immune escape. Shielding the NPs protects the cargoes from premature release and further modification of the surface of nanovaccines with certain receptors enhances the targeting ability as well as helps in enhanced accumulation [98].
Clinical Aspects of Biomimetic Nanovaccines
Very few vaccine candidates have successfully reached the clinic after preclinical evaluations. Most vaccines that are available now in the market can elicit only humoral responses, thereby availing the need for the development of vaccines that can generate strong cellular responses for certain infectious diseases and cancer.
One of such biomimetic nanovaccines is “Mosquirix”, which was proved to be effective against malaria. This nanovaccine constituted the circumsporozoite protein of Plasmodium falciparum and MPLA 4 with a saponin adjuvant QS-21 [152].
Another nanovaccine, which is currently under clinical trials as “Vaxfectin®”, is cationic liposomal formulation by encapsulating therapeutic DNA vaccines against the herpes simplex virus type-2 (HSV-2).
Vaxfectin® nanovaccines are also used for DNA immunization against influenza virus H5N1, and are also under clinical trials [153]. Another FDA-approved nanovaccine is Inflexal®V, where the HA surface molecules of the influenza virus are directly fused with lipid components, and used as a subunit influenza vaccine [154].
Generalized modulus for membrane antigens (GMMA) was derived from the outer membrane of genetically modified gram-negative bacteria. It can produce Penta-acylated lipopolysaccharide, and these vaccines were used against bacterial infection Shigellosis, and are in clinical trials now [155].
In addition to the nanovaccines, as mentioned earlier, Stimuvax® is another therapeutic liposome vaccine against cancer. It has a lipo-peptide called Tecemotide, which is used as an antigen target specific tumor antigens.
However, this vaccine failed in the III phase of clinical trials [156]. Another liposomal therapeutic vaccine, which is a modified form of Stimuvax®, is currently under clinical trials; this nanovaccine is composed of a synthetic peptide (antigen), an MPLA immunoadjuvant, and lipids [157].
Another biomimetic nanovaccine is Epaxal, a viral liposomal nanovaccine that uses viral glycoprotein fused with lipids as an adjuvant, and that is used against hepatitis A infection [158].
1. Rosenblum M.D., Remedios K.A., Abbas A.K. Mechanisms of human autoimmunity. J. Clin. Investig. 2015;125:2228–2233. doi: 10.1172/JCI78088. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
2. Wang L., Wang F.S., Gershwin M.E. Human autoimmune diseases: A comprehensive update. J. Intern. Med. 2015;278:369–395. doi: 10.1111/joim.12395. [PubMed] [CrossRef] [Google Scholar]
3. Arias C.A., Murray B.E. Antibiotic-resistant bugs in the 21st century—A clinical super-challenge. N. Engl. J. Med. 2009;360:439–443. doi: 10.1056/NEJMp0804651. [PubMed] [CrossRef] [Google Scholar]
4. Larché M., Wraith D.C. Peptide-based therapeutic vaccines for allergic and autoimmune diseases. Nat. Med. 2005;11:S69. doi: 10.1038/nm1226. [PubMed] [CrossRef] [Google Scholar]
5. Wraith D.C. Therapeutic peptide vaccines for treatment of autoimmune diseases. Immunol. Lett. 2009;122:134–136. doi: 10.1016/j.imlet.2008.11.013. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
6. Anderson R.P., Jabri B. Vaccine against autoimmune disease: Antigen-specific immunotherapy. Curr. Opin. Immunol. 2013;25:410–417. doi: 10.1016/j.coi.2013.02.004. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
7. Anderton S.M., Wraith D.C. Hierarchy in the ability of T cell epitopes to induce peripheral tolerance to antigens from myelin. Eur. J. Immunol. 1998;28:1251–1261. doi: 10.1002/(SICI)1521-4141(199804)28:04<1251::AID-IMMU1251>3.0.CO;2-O. [PubMed] [CrossRef] [Google Scholar]
8. Plotkin S. History of vaccination. Proc. Natl. Acad. Sci. USA. 2014;111:12283–12287. doi: 10.1073/pnas.1400472111. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
9. Whitney C.G., Zhou F., Singleton J., Schuchat A. Benefits from immunization during the vaccines for children program era—United States, 1994–2013. MMWR Morb. Mortal. Wkly. Rep. 2014;63:352. [PMC free article] [PubMed] [Google Scholar]
10. Lee H.B., Yoon S.Y., Singh B., Oh S.H., Cui L., Yan C., Kang S.K., Choi Y.J., Cho C.S. Oral Immunization of FMDV Vaccine Using pH-Sensitive and Mucoadhesive Thiolated Cellulose Acetate Phthalate Microparticles. Tissue Eng. Regen. Med. 2018;15:1–11. doi: 10.1007/s13770-017-0082-x. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
11. Glenny A.T., Pope C.G., Waddington H., Wallace U. Immunological notes. XVII–XXIV. J. Pathol. Bacteriol. 1926;29:31–40. doi: 10.1002/path.1700290106. [CrossRef] [Google Scholar]
12. Petrovsky N. Comparative Safety of Vaccine Adjuvants: A Summary of Current Evidence and Future Needs. Drug Saf. 2015;38:1059–1074. doi: 10.1007/s40264-015-0350-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
13. Opie E.L., Freund J. An Experimental Study of Protective Inoculation with Heat Killed Tubercle Bacilli. J. Exp. Med. 1937;66:761. doi: 10.1084/jem.66.6.761. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
14. Sun B., Xia T. Nanomaterial-based vaccine adjuvants. J. Mater. Chem. B. 2016;4:5496–5509. doi: 10.1039/C6TB01131D. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
15. Krishnamachari Y., Geary S.M., Lemke C.D., Salem A.K. Nanoparticle delivery systems in cancer vaccines. Pharm. Res. 2011;28:215–236. doi: 10.1007/s11095-010-0241-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
16. Zhang X.Q., Dahle C.E., Baman N.K., Rich N., Weiner G.J., Salem A.K. Potent antigen-specific immune responses stimulated by codelivery of CpG ODN and antigens in degradable microparticles. J. Immunother. 2007;30:469–478. doi: 10.1097/CJI.0b013e31802fd8c6. [PubMed] [CrossRef] [Google Scholar]
17. Hokmabad V.R., Davaran S., Aghazadeh M., Alizadeh E., Salehi R., Ramazani A. A Comparison of the Effects of Silica and Hydroxyapatite Nanoparticles on Poly(ε-caprolactone)-Poly(ethylene glycol)-Poly(ε-caprolactone)/Chitosan Nanofibrous Scaffolds for Bone Tissue Engineering. Tissue Eng. Regen. Med. 2018;15:735–750. doi: 10.1007/s13770-018-0140-z. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
18. Joshi V.B., Geary S.M., Salem A.K. Biodegradable particles as vaccine delivery systems: Size matters. AAPS J. 2013;15:85–94. doi: 10.1208/s12248-012-9418-6. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
19. Bishop C.J., Kozielski K.L., Green J.J. Exploring the role of polymer structure on intracellular nucleic acid delivery via polymeric nanoparticles. J. Control. Release. 2015;219:488–499. doi: 10.1016/j.jconrel.2015.09.046. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
20. Wang F., Fang R.H., Luk B.T., Hu C.M.J., Thamphiwatana S., Dehaini D., Angsantikul P., Kroll A.V., Pang Z., Gao W. Nanoparticle-Based Antivirulence Vaccine for the Management of Methicillin-Resistant Staphylococcus aureus Skin Infection. Adv. Funct. Mater. 2016;26:1628–1635. doi: 10.1002/adfm.201505231. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
21. Moon J.J., Suh H., Polhemus M.E., Ockenhouse C.F., Yadava A., Irvine D.J. Antigen-Displaying Lipid-Enveloped PLGA Nanoparticles as Delivery Agents for a Plasmodium vivax Malaria Vaccine. PLoS ONE. 2012;7:e31472. doi: 10.1371/journal.pone.0031472. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
22. Wang L., Hu C., Shao L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017;12:1227. doi: 10.2147/IJN.S121956. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
23. Sahu K.K., Pandey R.S. Immunological evaluation of colonic delivered Hepatitis B surface antigen loaded TLR-4 agonist modified solid fat nanoparticles. Int. Immunopharmacol. 2016;39:343–352. doi: 10.1016/j.intimp.2016.08.007. [PubMed] [CrossRef] [Google Scholar]
24. Li S., Rizzo M., Bhattacharya S., Huang L. Characterization of cationic lipid-protamine–DNA (LPD) complexes for intravenous gene delivery. Gene Ther. 1998;5:930. doi: 10.1038/sj.gt.3300683. [PubMed] [CrossRef] [Google Scholar]
25. Hu C.M.J., Fang R.H., Luk B.T., Zhang L. Nanoparticle-detained toxins for safe and effective vaccination. Nat. Nanotechnol. 2013;8:933–938. doi: 10.1038/nnano.2013.254. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
26. Oberg A.L., Kennedy R.B., Li P., Ovsyannikova I.G., Poland G.A. Systems biology approaches to new vaccine development. Curr. Opin. Immunol. 2011;23:436–443. doi: 10.1016/j.coi.2011.04.005. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
27. Delany I., Rappuoli R., De Gregorio E. Vaccines for the 21st century. EMBO Mol. Med. 2014;6:708–720. doi: 10.1002/emmm.201403876. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
28. Mamo T., Poland G.A. Nanovaccinology: The next generation of vaccines meets 21st century materials science and engineering. Vaccine. 2012;30:6609–6611. doi: 10.1016/j.vaccine.2012.08.023. [PubMed] [CrossRef] [Google Scholar]
29. Park W., Heo Y.J., Han D.K. New opportunities for nanoparticles in cancer immunotherapy. Biomater. Res. 2018;22:24. doi: 10.1186/s40824-018-0133-y. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
30. Pillarisetti S., Uthaman S., Huh K.M., Koh Y.S., Lee S., Park I.K. Multimodal Composite Iron Oxide Nanoparticles for Biomedical Applications. Tissue Eng. Regen. Med. 2019;16:451–465. doi: 10.1007/s13770-019-00218-7. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
31. Singh R.K., Kim H.W. Inorganic nanobiomaterial drug carriers for medicine. Tissue Eng. Regen. Med. 2013;10:296–309. doi: 10.1007/s13770-013-1092-y. [CrossRef] [Google Scholar]
32. Yahyapour R., Farhood B., Graily G., Rezaeyan A., Rezapoor S., Abdollahi H., Cheki M., Amini P., Fallah H., Najafi M., et al. Stem Cell Tracing Through MR Molecular Imaging. Tissue Eng. Regen. Med. 2018;15:249–261. doi: 10.1007/s13770-017-0112-8. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
33. Yang C., Park G.K., McDonald E.J., Choi H.S. Targeted Near-Infrared Fluorescence Imaging for Regenerative Medicine. Tissue Eng. Regen. Med. 2019;16:433–442. doi: 10.1007/s13770-019-00219-6. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
34. Corbo C., Molinaro R., Tabatabaei M., Farokhzad O.C., Mahmoudi M. Personalized protein corona on nanoparticles and its clinical implications. Biomater. Sci. 2017;5:378–387. doi: 10.1039/C6BM00921B. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
35. Corbo C., Molinaro R., Parodi A., Toledano Furman N.E., Salvatore F., Tasciotti E. The impact of nanoparticle protein corona on cytotoxicity, immunotoxicity and target drug delivery. Nanomedicine. 2015;11:81–100. doi: 10.2217/nnm.15.188. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
36. Fang R.H., Kroll A.V., Gao W., Zhang L. Cell Membrane Coating Nanotechnology. Adv. Mater. 2018;30:1706759. doi: 10.1002/adma.201706759. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
37. Gao W., Hu C.M.J., Fang R.H., Luk B.T., Su J., Zhang L. Surface Functionalization of Gold Nanoparticles with Red Blood Cell Membranes. Adv. Mater. 2013;25:3549–3553. doi: 10.1002/adma.201300638. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
38. Rao M. Stem cells for therapy. Tissue Eng. Regen. Med. 2013;10:223–229. doi: 10.1007/s13770-013-1081-1. [CrossRef] [Google Scholar]
39. Vijayan V., Uthaman S., Park I.K. Cell Membrane-Camouflaged Nanoparticles: A Promising Biomimetic Strategy for Cancer Theragnostics. Polymers. 2018;10:983. doi: 10.3390/polym10090983. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
40. Peer D., Karp J.M., Hong S., Farokhzad O.C., Margalit R., Langer R. Nanocarriers as an emerging platform for cancer therapy. Nat. Nanotechnol. 2007;2:751. doi: 10.1038/nnano.2007.387. [PubMed] [CrossRef] [Google Scholar]
41. Hu C.M.J., Fang R.H., Zhang L. Erythrocyte-Inspired Delivery Systems. Adv. Healthc. Mater. 2012;1:537–547. doi: 10.1002/adhm.201200138. [PubMed] [CrossRef] [Google Scholar]
42. Hu C.M.J., Zhang L., Aryal S., Cheung C., Fang R.H., Zhang L. Erythrocyte membrane-camouflaged polymeric nanoparticles as a biomimetic delivery platform. Proc. Natl. Acad. Sci. USA. 2011;108:10980–10985. doi: 10.1073/pnas.1106634108. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
43. Gao M., Liang C., Song X., Chen Q., Jin Q., Wang C., Liu Z. Erythrocyte-Membrane-Enveloped Perfluorocarbon as Nanoscale Artificial Red Blood Cells to Relieve Tumor Hypoxia and Enhance Cancer Radiotherapy. Adv. Mater. 2017;29:1701429. doi: 10.1002/adma.201701429. [PubMed] [CrossRef] [Google Scholar]
44. Wei X., Ran D., Campeau A., Xiao C., Zhou J., Dehaini D., Jiang Y., Kroll A.V., Zhang Q., Gao W., et al. Multiantigenic Nanotoxoids for Antivirulence Vaccination against Antibiotic-Resistant Gram-Negative Bacteria. Nano Lett. 2019;19:4760–4769. doi: 10.1021/acs.nanolett.9b01844. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
45. He W., Frueh J., Wu Z., He Q. How Leucocyte Cell Membrane Modified Janus Microcapsules are Phagocytosed by Cancer Cells. ACS Appl. Mater. Interfaces. 2016;8:4407–4415. doi: 10.1021/acsami.5b10885. [PubMed] [CrossRef] [Google Scholar]
46. He H., Guo C., Wang J., Korzun W.J., Wang X.Y., Ghosh S., Yang H. Leutusome: A Biomimetic Nanoplatform Integrating Plasma Membrane Components of Leukocytes and Tumor Cells for Remarkably Enhanced Solid Tumor Homing. Nano Lett. 2018;18:6164–6174. doi: 10.1021/acs.nanolett.8b01892. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
47. Wei X., Zhang G., Ran D., Krishnan N., Fang R.H., Gao W., Spector S.A., Zhang L. T-Cell-Mimicking Nanoparticles Can Neutralize HIV Infectivity. Adv. Mater. 2018;30:1802233. doi: 10.1002/adma.201802233. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
48. Pitchaimani A., Nguyen T.D.T., Marasini R., Eliyapura A., Azizi T., Jaberi-Douraki M., Aryal S. Biomimetic Natural Killer Membrane Camouflaged Polymeric Nanoparticle for Targeted Bioimaging. Adv. Funct. Mater. 2019;29:1806817. doi: 10.1002/adfm.201806817. [CrossRef] [Google Scholar]
49. Hu C.M.J., Fang R.H., Wang K.C., Luk B.T., Thamphiwatana S., Dehaini D., Nguyen P., Angsantikul P., Wen C.H., Kroll A.V., et al. Nanoparticle biointerfacing by platelet membrane cloaking. Nature. 2015;526:118. doi: 10.1038/nature15373. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
50. Xuan M., Shao J., Dai L., He Q., Li J. Macrophage Cell Membrane Camouflaged Mesoporous Silica Nanocapsules for In Vivo Cancer Therapy. Adv. Healthc. Mater. 2015;4:1645–1652. doi: 10.1002/adhm.201500129. [PubMed] [CrossRef] [Google Scholar]
51. Zhu J.Y., Zheng D.W., Zhang M.K., Yu W.Y., Qiu W.X., Hu J.J., Feng J., Zhang X.Z. Preferential cancer cell self-recognition and tumor self-targeting by coating nanoparticles with homotypic cancer cell membranes. Nano Lett. 2016;16:5895–5901. doi: 10.1021/acs.nanolett.6b02786. [PubMed] [CrossRef] [Google Scholar]
52. Yang R., Xu J., Xu L., Sun X., Chen Q., Zhao Y., Peng R., Liu Z. Cancer Cell Membrane-Coated Adjuvant Nanoparticles with Mannose Modification for Effective Anticancer Vaccination. ACS Nano. 2018;12:5121–5129. doi: 10.1021/acsnano.7b09041. [PubMed] [CrossRef] [Google Scholar]
53. Lee L.A., Wang Q. Adaptations of nanoscale viruses and other protein cages for medical applications. Nanomed. Nanotechnol. Biol. Med. 2006;2:137–149. doi: 10.1016/j.nano.2006.07.009. [PubMed] [CrossRef] [Google Scholar]
54. Kang S., Pushko P., Bright R., Smith G., Compans R. Vaccines for Pandemic Influenza. Springer; Berlin, Germany: 2009. Influenza virus-like particles as pandemic vaccines; pp. 269–289. [PubMed] [Google Scholar]
55. Castón J.R., Carrascosa J.L. Structure and Physics of Viruses. Springer; Berlin, Germany: 2013. The basic architecture of viruses; pp. 53–75. [PubMed] [Google Scholar]
56. Roldão A., Mellado M.C.M., Castilho L.R., Carrondo M.J.T., Alves P.M. Virus-like particles in vaccine development. Expert Rev. Vaccines. 2010;9:1149–1176. doi: 10.1586/erv.10.115. [PubMed] [CrossRef] [Google Scholar]
57. Kaczmarczyk S.J., Sitaraman K., Young H.A., Hughes S.H., Chatterjee D.K. Protein delivery using engineered virus-like particles. Proc. Natl. Acad. Sci. USA. 2011;108:16998. doi: 10.1073/pnas.1101874108. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
58. Wu C.Y., Yeh Y.C., Chan J.T., Yang Y.C., Yang J.R., Liu M.T., Wu H.S., Hsiao P.W. A VLP vaccine induces broad-spectrum cross-protective antibody immunity against H5N1 and H1N1 subtypes of influenza A virus. PLoS ONE. 2012;7:e42363. doi: 10.1371/journal.pone.0042363. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
59. Bai H., Wang C., Qi Y., Xu J., Li N., Chen L., Jiang B., Zhu X., Zhang H., Li X., et al. Major vault protein suppresses lung cancer cell proliferation by inhibiting STAT3 signaling pathway. BMC Cancer. 2019;19:454. doi: 10.1186/s12885-019-5665-6. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
60. Angsantikul P., Thamphiwatana S., Zhang Q., Spiekermann K., Zhuang J., Fang R.H., Gao W., Obonyo M., Zhang L. Coating nanoparticles with gastric epithelial cell membrane for targeted antibiotic delivery against Helicobacter pylori infection. Adv. Ther. 2018;1:1800016. doi: 10.1002/adtp.201800016. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
61. Gao W., Fang R.H., Thamphiwatana S., Luk B.T., Li J., Angsantikul P., Zhang Q., Hu C.M.J., Zhang L. Modulating antibacterial immunity via bacterial membrane-coated nanoparticles. Nano Lett. 2015;15:1403–1409. doi: 10.1021/nl504798g. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
62. Henriksen-Lacey M., Korsholm K.S., Andersen P., Perrie Y., Christensen D. Liposomal vaccine delivery systems. Expert Opin. Drug Deliv. 2011;8:505–519. doi: 10.1517/17425247.2011.558081. [PubMed] [CrossRef] [Google Scholar]
63. Heurtault B., Frisch B., Pons F. Liposomes as delivery systems for nasal vaccination: Strategies and outcomes. Expert Opin. Drug Deliv. 2010;7:829–844. doi: 10.1517/17425247.2010.488687. [PubMed] [CrossRef] [Google Scholar]
64. Bose R.J., Lee S.H., Park H. Lipid-based surface engineering of PLGA nanoparticles for drug and gene delivery applications. Biomater. Res. 2016;20:34. doi: 10.1186/s40824-016-0081-3. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
65. Sharma S., Mukkur T., Benson H.A., Chen Y. Pharmaceutical aspects of intranasal delivery of vaccines using particulate systems. J. Pharm. Sci. 2009;98:812–843. doi: 10.1002/jps.21493. [PubMed] [CrossRef] [Google Scholar]
66. Khatri K., Goyal A.K., Gupta P.N., Mishra N., Mehta A., Vyas S.P. Surface modified liposomes for nasal delivery of DNA vaccine. Vaccine. 2008;26:2225–2233. doi: 10.1016/j.vaccine.2008.02.058. [PubMed] [CrossRef] [Google Scholar]
67. Moser C., Amacker M., Zurbriggen R. Influenza virosomes as a vaccine adjuvant and carrier system. Expert Rev. Vaccines. 2011;10:437–446. doi: 10.1586/erv.11.15. [PubMed] [CrossRef] [Google Scholar]
68. Li Sd Huang L.Y. In vivo gene transfer via intravenous administration of cationic lipid–protamine–DNA (LPD) complexes. Gene Ther. 1997;4:891. [PubMed] [Google Scholar]
69. Noad R., Roy P. Virus-like particles as immunogens. Trends Microbiol. 2003;11:438–444. doi: 10.1016/S0966-842X(03)00208-7. [PubMed] [CrossRef] [Google Scholar]
70. Grgacic E.V.L., Anderson D.A. Virus-like particles: Passport to immune recognition. Methods. 2006;40:60–65. doi: 10.1016/j.ymeth.2006.07.018. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
71. Zhang L.F., Zhou J., Chen S., Cai L.L., Bao Q.Y., Zheng F.Y., Lu J.Q., Padmanabha J., Hengst K., Malcolm K., et al. HPV6b virus like particles are potent immunogens without adjuvant in man. Vaccine. 2000;18:1051–1058. doi: 10.1016/S0264-410X(99)00351-5. [PubMed] [CrossRef] [Google Scholar]
72. André F.E. Overview of a 5-year clinical experience with a yeast-derived hepatitis B vaccine. Vaccine. 1990;8:S74–S78. doi: 10.1016/0264-410X(90)90222-8. [PubMed] [CrossRef] [Google Scholar]
73. Cutts F.T., Franceschi S., Goldie S., Castellsague X., de Sanjose S., Garnett G., Edmunds W.J., Claeys P., Goldenthal K.L., Harper D.M., et al. Human papillomavirus and HPV vaccines: A review. Bull. World Health Organ. 2007;85:719–726. doi: 10.2471/BLT.06.038414. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
74. Yatvin M.B., Weinstein J.N., Dennis W.H., Blumenthal R. Design of liposomes for enhanced local release of drugs by hyperthermia. Science. 1978;202:1290–1293. doi: 10.1126/science.364652. [PubMed] [CrossRef] [Google Scholar]
75. Kingsman S.M., Kingsman A.J. Polyvalent recombinant antigens: A new vaccine strategy. Vaccine. 1988;6:304–306. doi: 10.1016/0264-410X(88)90174-0. [PubMed] [CrossRef] [Google Scholar]
76. Maurer P., Jennings G.T., Willers J., Rohner F., Lindman Y., Roubicek K., Renner W.A., Müller P., Bachmann M.F. A therapeutic vaccine for nicotine dependence: Preclinical efficacy, and phase I safety and immunogenicity. Eur. J. Immunol. 2005;35:2031–2040. doi: 10.1002/eji.200526285. [PubMed] [CrossRef] [Google Scholar]
77. Hu Y.C. Baculovirus as a highly efficient expression vector in insect and mammalian cells. Acta Pharmacol. Sin. 2005;26:405–416. doi: 10.1111/j.1745-7254.2005.00078.x. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
78. Andrews S.C. The Ferritin-like superfamily: Evolution of the biological iron storeman from a rubrerythrin-like ancestor. Biochim. Et Biophys. Acta (BBA) Gen. Subj. 2010;1800:691–705. doi: 10.1016/j.bbagen.2010.05.010. [PubMed] [CrossRef] [Google Scholar]
79. Chasteen N.D., Harrison P.M. Mineralization in ferritin: An efficient means of iron storage. J. Struct. Biol. 1999;126:182–194. doi: 10.1006/jsbi.1999.4118. [PubMed] [CrossRef] [Google Scholar]
80. Kanekiyo M., Wei C.J., Yassine H.M., McTamney P.M., Boyington J.C., Whittle J.R., Rao S.S., Kong W.P., Wang L., Nabel G.J. Self-assembling influenza nanoparticle vaccines elicit broadly neutralizing H1N1 antibodies. Nature. 2013;499:102. doi: 10.1038/nature12202. [PubMed] [CrossRef] [Google Scholar]
81. Champion C.I., Kickhoefer V.A., Liu G., Moniz R.J., Freed A.S., Bergmann L.L., Vaccari D., Raval-Fernandes S., Chan A.M., Rome L.H. A vault nanoparticle vaccine induces protective mucosal immunity. PLoS ONE. 2009;4:e5409. doi: 10.1371/journal.pone.0005409. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
82. Wahome N., Pfeiffer T., Ambiel I., Yang Y., Keppler O.T., Bosch V., Burkhard P. Conformation-specific Display of 4E10 and 2F5 Epitopes on Self-assembling Protein Nanoparticles as a Potential HIV Vaccine. Chem. Biol. Drug Des. 2012;80:349–357. doi: 10.1111/j.1747-0285.2012.01423.x. [PubMed] [CrossRef] [Google Scholar]
83. Kuehn M.J., Kesty N.C. Bacterial outer membrane vesicles and the host–pathogen interaction. Genes Dev. 2005;19:2645–2655. doi: 10.1101/gad.1299905. [PubMed] [CrossRef] [Google Scholar]
84. Lim W., Kim H.S. Exosomes as Therapeutic Vehicles for Cancer. Tissue Eng. Regen. Med. 2019;16:213–223. doi: 10.1007/s13770-019-00190-2. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
85. Théry C., Ostrowski M., Segura E. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 2009;9:581. doi: 10.1038/nri2567. [PubMed] [CrossRef] [Google Scholar]
86. Ståhl A.L., Johansson K., Mossberg M., Kahn R., Karpman D. Exosomes and microvesicles in normal physiology, pathophysiology, and renal diseases. Pediatr. Nephrol. 2019;34:11–30. doi: 10.1007/s00467-017-3816-z. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
87. Zhang W., Jiang X., Bao J., Wang Y., Liu H., Tang L. Exosomes in pathogen infections: A bridge to deliver molecules and link functions. Front. Immunol. 2018;9:90. doi: 10.3389/fimmu.2018.00090. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
88. Cox J.C., Coulter A.R. Adjuvants—A classification and review of their modes of action. Vaccine. 1997;15:248–256. doi: 10.1016/S0264-410X(96)00183-1. [PubMed] [CrossRef] [Google Scholar]
89. Hoebe K., Janssen E., Beutler B. The interface between innate and adaptive immunity. Nat. Immunol. 2004;5:971–974. doi: 10.1038/ni1004-971. [PubMed] [CrossRef] [Google Scholar]
90. Fraser C.K., Diener K.R., Brown M.P., Hayball J.D. Improving vaccines by incorporating immunological coadjuvants. Expert Rev. Vaccines. 2007;6:559–578. doi: 10.1586/14760584.6.4.559. [PubMed] [CrossRef] [Google Scholar]
91. Riitho V., Walters A.A., Somavarapu S., Lamp B., Rümenapf T., Krey T., Rey F.A., Oviedo-Orta E., Stewart G.R., Locker N., et al. Design and evaluation of the immunogenicity and efficacy of a biomimetic particulate formulation of viral antigens. Sci. Rep. 2017;7:13743. doi: 10.1038/s41598-017-13915-x. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
92. Le Q.V., Suh J., Choi J.J., Park G.T., Lee J.W., Shim G., Oh Y.K. In Situ Nanoadjuvant-Assembled Tumor Vaccine for Preventing Long-Term Recurrence. ACS Nano. 2019;13:7442–7462. doi: 10.1021/acsnano.9b02071. [PubMed] [CrossRef] [Google Scholar]
93. Siefert A.L., Caplan M.J., Fahmy T.M. Artificial bacterial biomimetic nanoparticles synergize pathogen-associated molecular patterns for vaccine efficacy. Biomaterials. 2016;97:85–96. doi: 10.1016/j.biomaterials.2016.03.039. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
94. Lin L.C.W., Huang C.Y., Yao B.Y., Lin J.C., Agrawal A., Algaissi A., Peng B.H., Liu Y.H., Huang P.H., Juang R.H. Viromimetic STING Agonist-Loaded Hollow Polymeric Nanoparticles for Safe and Effective Vaccination against Middle East Respiratory Syndrome Coronavirus. Adv. Funct. Mater. 2019;29:1807616. doi: 10.1002/adfm.201807616. [CrossRef] [Google Scholar]
95. Chen P., Liu X., Sun Y., Zhou P., Wang Y., Zhang Y. Dendritic cell targeted vaccines: Recent progresses and challenges. Hum. Vaccines Immunother. 2016;12:612–622. doi: 10.1080/21645515.2015.1105415. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
96. Deng L., Mohan T., Chang T.Z., Gonzalez G.X., Wang Y., Kwon Y.M., Kang S.M., Compans R.W., Champion J.A., Wang B.Z. Double-layered protein nanoparticles induce broad protection against divergent influenza A viruses. Nat. Commun. 2018;9:359. doi: 10.1038/s41467-017-02725-4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
97. Bachmann M.F., Jennings G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010;10:787. doi: 10.1038/nri2868. [PubMed] [CrossRef] [Google Scholar]
98. Yu W., He X., Yang Z., Yang X., Xiao W., Liu R., Xie R., Qin L., Gao H. Sequentially responsive biomimetic nanoparticles with optimal size in combination with checkpoint blockade for cascade synergetic treatment of breast cancer and lung metastasis. Biomaterials. 2019;217:119309. doi: 10.1016/j.biomaterials.2019.119309. [PubMed] [CrossRef] [Google Scholar]
99. Grenni P., Ancona V., Caracciolo A.B. Ecological effects of antibiotics on natural ecosystems: A review. Microchem. J. 2018;136:25–39. doi: 10.1016/j.microc.2017.02.006. [CrossRef] [Google Scholar]
100. Jung S.W., Oh S.H., Lee I.S., Byun J.H., Lee J.H. In Situ Gelling Hydrogel with Anti-Bacterial Activity and Bone Healing Property for Treatment of Osteomyelitis. Tissue Eng. Regen. Med. 2019;16:479–490. doi: 10.1007/s13770-019-00206-x. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
101. Dhanasooraj D., Kumar R.A., Mundayoor S. Vaccine delivery system for tuberculosis based on nano-sized hepatitis B virus core protein particles. Int. J. Nanomed. 2013;8:835. [PMC free article] [PubMed] [Google Scholar]
102. Bai J., Yang E., Chang P.S., Ryu S. Preparation and characterization of endolysin-containing liposomes and evaluation of their antimicrobial activities against gram-negative bacteria. Enzym. Microb. Technol. 2019;128:40–48. doi: 10.1016/j.enzmictec.2019.05.006. [PubMed] [CrossRef] [Google Scholar]
103. Xia Q., Zhang Y., Li Z., Hou X., Feng N. Red blood cell membrane-camouflaged nanoparticles: A novel drug delivery system for antitumor application. Acta Pharm. Sin. B. 2019;9:675–689. doi: 10.1016/j.apsb.2019.01.011. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
104. Esté J.A., Cihlar T. Current status and challenges of antiretroviral research and therapy. Antivir. Res. 2010;85:25–33. doi: 10.1016/j.antiviral.2009.10.007. [PubMed] [CrossRef] [Google Scholar]
105. Mamo T., Moseman E.A., Kolishetti N., Salvador-Morales C., Shi J., Kuritzkes D.R., Langer R., Andrian Uv Farokhzad O.C. Emerging nanotechnology approaches for HIV/AIDS treatment and prevention. Nanomedicine. 2010;5:269–285. doi: 10.2217/nnm.10.1. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
106. Nayak D., Boxi A., Ashe S., Thathapudi N.C., Nayak B. Stavudine loaded gelatin liposomes for HIV therapy: Preparation, characterization and in vitro cytotoxic evaluation. Mater. Sci. Eng. C. 2017;73:406–416. doi: 10.1016/j.msec.2016.12.073. [PubMed] [CrossRef] [Google Scholar]
107. Bui T., Dykers T., Hu S.L., Faltynek C.R., Ho R. Effect of MTP-PE liposomes and interleukin-7 on induction of antibody and cell-mediated immune responses to a recombinant HIV-envelope protein. J. Acquir. Immune Defic. Syndr. 1994;7:799–806. [PubMed] [Google Scholar]
108. Hanson M.C., Abraham W., Crespo M.P., Chen S.H., Liu H., Szeto G.L., Kim M., Reinherz E.L., Irvine D.J. Liposomal vaccines incorporating molecular adjuvants and intrastructural T-cell help promote the immunogenicity of HIV membrane-proximal external region peptides. Vaccine. 2015;33:861–868. doi: 10.1016/j.vaccine.2014.12.045. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
109. Poteet E., Lewis P., Chen C., Ho S.O., Do T., Chiang S., Labranche C., Montefiori D., Fujii G., Yao Q. Toll-like receptor 3 adjuvant in combination with virus-like particles elicit a humoral response against HIV. Vaccine. 2016;34:5886–5894. doi: 10.1016/j.vaccine.2016.10.036. [PubMed] [CrossRef] [Google Scholar]
110. Andersson A.M.C., Ragonnaud E., Seaton K.E., Sawant S., Folgori A., Colloca S., Labranche C., Montefiori D.C., Tomaras G.D., Holst P.J. Effect of HIV-1 envelope cytoplasmic tail on adenovirus primed virus encoded virus-like particle immunizations. Vaccine. 2016;34:5344–5351. doi: 10.1016/j.vaccine.2016.08.089. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
111. Kovacs J.M., Nkolola J.P., Peng H., Cheung A., Perry J., Miller C.A., Seaman M.S., Barouch D.H., Chen B. HIV-1 envelope trimer elicits more potent neutralizing antibody responses than monomeric gp120. Proc. Natl. Acad. Sci. USA. 2012;109:12111–12116. doi: 10.1073/pnas.1204533109. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
112. Pejawar-Gaddy S., Kovacs J.M., Barouch D.H., Chen B., Irvine D.J. Design of lipid nanocapsule delivery vehicles for multivalent display of recombinant Env trimers in HIV vaccination. Bioconjugate Chem. 2014;25:1470–1478. doi: 10.1021/bc5002246. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
113. Campbell E.M., Hope T.J. HIV-1 capsid: The multifaceted key player in HIV-1 infection. Nat. Rev. Microbiol. 2015;13:471. doi: 10.1038/nrmicro3503. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
114. Dennis E., Peoples V., Johnson F., Bibbs R., Topps D., Bopda-Waffo A., Coats M. Utilizing nanotechnology to combat malaria. J. Infect. Dis. Ther. 2015;3:229. [Google Scholar]
115. Rahman K., Khan S.U., Fahad S., Chang M.X., Abbas A., Khan W.U., Rahman L., Haq Z.U., Nabi G., Khan D. Nano-biotechnology: A new approach to treat and prevent malaria. Int. J. Nanomed. 2019;14:1401. doi: 10.2147/IJN.S190692. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
116. Ibrahim S., Tagami T., Ozeki T. Effective-Loading of Platinum–Chloroquine into PEGylated Neutral and Cationic Liposomes as a Drug Delivery System for Resistant Malaria Parasites. Biol. Pharm. Bull. 2017;40:815–823. doi: 10.1248/bpb.b16-00914. [PubMed] [CrossRef] [Google Scholar]
117. Peeters P., Huiskamp C.W., Eling W., Crommelin D. Chloroquine containing liposomes in the chemotherapy of murine malaria. Parasitology. 1989;98:381–386. doi: 10.1017/S003118200006145X. [PubMed] [CrossRef] [Google Scholar]
118. Urbán P., Estelrich J., Cortés A., Fernàndez-Busquets X. A nanovector with complete discrimination for targeted delivery to Plasmodium falciparum-infected versus non-infected red blood cells in vitro. J. Control. Release. 2011;151:202–211. doi: 10.1016/j.jconrel.2011.01.001. [PubMed] [CrossRef] [Google Scholar]
119. Marques J., Valle-Delgado J.J., Urbán P., Baró E., Prohens R., Mayor A., Cisteró P., Delves M., Sinden R.E., Grandfils C. Adaptation of targeted nanocarriers to changing requirements in antimalarial drug delivery. Nanomed. Nanotechnol. Biol. Med. 2017;13:515–525. doi: 10.1016/j.nano.2016.09.010. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
120. Tyagi R.K., Garg N.K., Dalai S.K., Awasthi A. Transdermal immunization of P. falciparum surface antigen (MSP-119) via elastic liposomes confers robust immunogenicity. Hum. Vaccines Immunother. 2016;12:990–992. doi: 10.1080/21645515.2015.1116656. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
121. Seth L., Ferlez K.M.B., Kaba S.A., Musser D.M., Emadi S., Matyas G.R., Beck Z., Alving C.R., Burkhard P., Lanar D.E. Development of a self-assembling protein nanoparticle vaccine targeting Plasmodium falciparum Circumsporozoite Protein delivered in three Army Liposome Formulation adjuvants. Vaccine. 2017;35:5448–5454. doi: 10.1016/j.vaccine.2017.02.040. [PubMed] [CrossRef] [Google Scholar]
122. Leach A., Vekemans J., Lievens M., Ofori-Anyinam O., Cahill C., Owusu-Agyei S., Abdulla S., Macete E., Njuguna P., Savarese B. Design of a phase III multicenter trial to evaluate the efficacy of the RTS, S/AS01 malaria vaccine in children across diverse transmission settings in Africa. Malar. J. 2011;10:224. doi: 10.1186/1475-2875-10-224. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
123. Collins K.A., Snaith R., Cottingham M.G., Gilbert S.C., Hill A.V. Enhancing protective immunity to malaria with a highly immunogenic virus-like particle vaccine. Sci. Rep. 2017;7:46621. doi: 10.1038/srep46621. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
124. Rajendrakumar S., Mohapatra A., Singh B., Revuri V., Lee Y.K., Kim C., Cho C.S., Park I.K. Self-assembled, adjuvant/antigen-based nanovaccine mediates anti-tumor immune response against melanoma tumor. Polymers. 2018;10:1063. doi: 10.3390/polym10101063. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
125. Kroll A.V., Jiang Y., Zhou J., Holay M., Fang R.H., Zhang L. Biomimetic Nanoparticle Vaccines for Cancer Therapy. Adv. Biosyst. 2019;3:1800219. doi: 10.1002/adbi.201800219. [CrossRef] [Google Scholar]
126. Han S.W., Kim Y.Y., Kang W.J., Kim H.C., Ku S.Y., Kang B.C., Yun J.W. The Use of Normal Stem Cells and Cancer Stem Cells for Potential Anti-Cancer Therapeutic Strategy. Tissue Eng. Regen. Med. 2018;15:365–380. doi: 10.1007/s13770-018-0128-8. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
127. Lou J., Zhang L., Zheng G. Advancing Cancer Immunotherapies with Nanotechnology. Adv. Ther. 2019;2:1800128. doi: 10.1002/adtp.201800128. [CrossRef] [Google Scholar]
128. Kunjachan S., Ehling J., Storm G., Kiessling F., Lammers T. Noninvasive imaging of nanomedicines and nanotheranostics: Principles, progress, and prospects. Chem. Rev. 2015;115:10907–10937. doi: 10.1021/cr500314d. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
129. Ichihashi T., Satoh T., Sugimoto C., Kajino K. Emulsified phosphatidylserine, simple and effective peptide carrier for induction of potent epitope-specific T cell responses. PLoS ONE. 2013;8:e60068. doi: 10.1371/journal.pone.0060068. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
130. Talesh G.A., Ebrahimi Z., Badiee A., Mansourian M., Attar H., Arabi L., Jalali S.A., Jaafari M.R. Poly (I: C)-DOTAP cationic nanoliposome containing multi-epitope HER2-derived peptide promotes vaccine-elicited anti-tumor immunity in a murine model. Immunol. Lett. 2016;176:57–64. doi: 10.1016/j.imlet.2016.05.016. [PubMed] [CrossRef] [Google Scholar]
131. Shariat S., Badiee A., Jalali S.A., Mansourian M., Yazdani M., Mortazavi S.A., Jaafari M.R. P5 HER2/neu-derived peptide conjugated to liposomes containing MPL adjuvant as an effective prophylactic vaccine formulation for breast cancer. Cancer Lett. 2014;355:54–60. doi: 10.1016/j.canlet.2014.09.016. [PubMed] [CrossRef] [Google Scholar]
132. Chen M., Chen M., He J. Cancer cell membrane cloaking nanoparticles for targeted co-delivery of doxorubicin and PD-L1 siRNA. Artif. Cells Nanomed. Biotechnol. 2019;47:1635–1641. doi: 10.1080/21691401.2019.1608219. [PubMed] [CrossRef] [Google Scholar]
133. Wang D., Dong H., Li M., Cao Y., Yang F., Zhang K., Dai W., Wang C., Zhang X. Erythrocyte–Cancer Hybrid Membrane Camouflaged Hollow Copper Sulfide Nanoparticles for Prolonged Circulation Life and Homotypic-Targeting Photothermal/Chemotherapy of Melanoma. ACS Nano. 2018;12:5241–5252. doi: 10.1021/acsnano.7b08355. [PubMed] [CrossRef] [Google Scholar]
134. Kroll A.V., Fang R.H., Jiang Y., Zhou J., Wei X., Yu C.L., Gao J., Luk B.T., Dehaini D., Gao W. Nanoparticulate delivery of cancer cell membrane elicits multiantigenic antitumor immunity. Adv. Mater. 2017;29:1703969. doi: 10.1002/adma.201703969. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
135. Kimiz-Gebologlu I., Gulce-Iz S., Biray-Avci C. Monoclonal antibodies in cancer immunotherapy. Mol. Biol. Rep. 2018;45:2935–2940. doi: 10.1007/s11033-018-4427-x. [PubMed] [CrossRef] [Google Scholar]
136. Zhou B., Jiang Q., Xiao X., Xu X., Xu Y., Kong Y., Zhang W., Zeng Y., Liu X., Luo B. Assisting anti-PD-1 antibody treatment with a liposomal system capable of recruiting immune cells. Nanoscale. 2019;11:7996–8011. doi: 10.1039/C9NR01434A. [PubMed] [CrossRef] [Google Scholar]
137. Lizotte P.H., Wen A.M., Sheen M.R., Fields J., Rojanasopondist P., Steinmetz N.F., Fiering S. In situ vaccination with cowpea mosaic virus nanoparticles suppresses metastatic cancer. Nat. Nanotechnol. 2015;11:295. doi: 10.1038/nnano.2015.292. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
138. Chattopadhyay S., Chen J.Y., Chen H.W., Hu C.M.J. Nanoparticle Vaccines Adopting Virus-like Features for Enhanced Immune Potentiation. Nanotheranostics. 2017;1:244–260. doi: 10.7150/ntno.19796. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
139. Patel J.M., Vartabedian V.F., Kim M.C., He S., Kang S.M., Selvaraj P. Influenza virus-like particles engineered by protein transfer with tumor-associated antigens induces protective antitumor immunity. Biotechnol. Bioeng. 2015;112:1102–1110. doi: 10.1002/bit.25537. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
140. Wang C., Fiering S.N., Steinmetz N.F. Cowpea Mosaic Virus Promotes Anti-Tumor Activity and Immune Memory in a Mouse Ovarian Tumor Model. Adv. Ther. 2019;2:1900003. doi: 10.1002/adtp.201900003. [CrossRef] [Google Scholar]
141. Kang T., Li C., Du T., Wu Y., Yang Y., Liu X., Zhang Q., Xu X., Gou M. A biomimetic nanoparticle-enabled toxoid vaccine against melittin. Int. J. Nanomed. 2018;13:3251–3261. doi: 10.2147/IJN.S156346. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
142. Chen Y., Chen M., Zhang Y., Lee J.H., Escajadillo T., Gong H., Fang R.H., Gao W., Nizet V., Zhang L. Broad-Spectrum Neutralization of Pore-Forming Toxins with Human Erythrocyte Membrane-Coated Nanosponges. Adv. Healthc. Mater. 2018;7:1701366. doi: 10.1002/adhm.201701366. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
143. Dykman L.A., Staroverov S.A., Mezhenny P.V., Fomin A.S., Kozlov S.V., Volkov A.A., Laskavy V.N., Shchyogolev S.Y. Use of a synthetic foot-and-mouth disease virus peptide conjugated to gold nanoparticles for enhancing immunological response. Gold Bull. 2015;48:93–101. doi: 10.1007/s13404-015-0165-1. [CrossRef] [Google Scholar]
144. Teng Z., Sun S., Chen H., Huang J., Du P., Dong H., Xu X., Mu S., Zhang Z., Guo H. Golden-star nanoparticles as adjuvant effectively promotes immune response to foot-and-mouth disease virus-like particles vaccine. Vaccine. 2018;36:6752–6760. doi: 10.1016/j.vaccine.2018.09.030. [PubMed] [CrossRef] [Google Scholar]
145. Wang C., Zhuang Y., Zhang Y., Luo Z., Gao N., Li P., Pan H., Cai L., Ma Y. Toll-like receptor 3 agonist complexed with cationic liposome augments vaccine-elicited antitumor immunity by enhancing TLR3–IRF3 signaling and type I interferons in dendritic cells. Vaccine. 2012;30:4790–4799. doi: 10.1016/j.vaccine.2012.05.027. [PubMed] [CrossRef] [Google Scholar]
146. Cruz L.J., Rueda F., Simón L., Cordobilla B., Albericio F., Domingo J.C. Liposomes containing NY-ESO-1/tetanus toxoid and adjuvant peptides targeted to human dendritic cells via the Fc receptor for cancer vaccines. Nanomedicine. 2014;9:435–449. doi: 10.2217/nnm.13.66. [PubMed] [CrossRef] [Google Scholar]
147. Shimizu T., Lila A.S.A., Kawaguchi Y., Shimazaki Y., Watanabe Y., Mima Y., Hashimoto Y., Okuhira K., Storm G., Ishima Y. A Novel Platform for Cancer Vaccines: Antigen-Selective Delivery to Splenic Marginal Zone B Cells via Repeated Injections of PEGylated Liposomes. J. Immunol. 2018;201:2969–2976. doi: 10.4049/jimmunol.1701351. [PubMed] [CrossRef] [Google Scholar]
148. Fantappie L., de Santis M., Chiarot E., Carboni F., Bensi G., Jousson O., Margarit I., Grandi G. Antibody-mediated immunity induced by engineered Escherichia coli OMVs carrying heterologous antigens in their lumen. J. Extracell. Vesicles. 2014;3:24015. doi: 10.3402/jev.v3.24015. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
149. Li L.L., Xu J.H., Qi G.B., Zhao X., Yu F., Wang H. Core–shell supramolecular gelatin nanoparticles for adaptive and “on-demand” antibiotic delivery. ACS Nano. 2014;8:4975–4983. doi: 10.1021/nn501040h. [PubMed] [CrossRef] [Google Scholar]
150. Arms L., Smith D.W., Flynn J., Palmer W., Martin A., Woldu A., Hua S. Advantages and limitations of current techniques for analyzing the biodistribution of nanoparticles. Front. Pharmacol. 2018;9:802. doi: 10.3389/fphar.2018.00802. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
151. Sercombe L., Veerati T., Moheimani F., Wu S.Y., Sood A.K., Hua S. Advances and challenges of liposome assisted drug delivery. Front. Pharmacol. 2015;6:286. doi: 10.3389/fphar.2015.00286. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
152. Didierlaurent A.M., Laupèze B., Di Pasquale A., Hergli N., Collignon C., Garçon N. Adjuvant system AS01, helping to overcome the challenges of modern vaccines. Expert Rev. Vaccines. 2017;16:55–63. doi: 10.1080/14760584.2016.1213632. [PubMed] [CrossRef] [Google Scholar]
153. Smith L.R., Wloch M.K., Ye M., Reyes L.R., Boutsaboualoy S., Dunne C.E., Chaplin J.A., Rusalov D., Rolland A.P., Fisher C.L. Phase 1 clinical trials of the safety and immunogenicity of adjuvanted plasmid DNA vaccines encoding influenza A virus H5 hemagglutinin. Vaccine. 2010;28:2565–2572. doi: 10.1016/j.vaccine.2010.01.029. [PubMed] [CrossRef] [Google Scholar]
154. Glück R., Metcalfe I.C. New technology platforms in the development of vaccines for the future. Vaccine. 2002;20:B10–B16. doi: 10.1016/S0264-410X(02)00513-3. [PubMed] [CrossRef] [Google Scholar]
155. Gerke C., Colucci A.M., Giannelli C., Sanzone S., Vitali C.G., Sollai L., Rossi O., Martin L.B., Auerbach J., Di Cioccio V., et al. Production of a Shigella sonnei Vaccine Based on Generalized Modules for Membrane Antigens (GMMA), 1790GAHB. PLoS ONE. 2015;10:e0134478. doi: 10.1371/journal.pone.0134478. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
156. Kroemer G., Zitvogel L., Galluzzi L. Victories and deceptions in tumor immunology. OncoImmunology. 2013;2:e23687. doi: 10.4161/onci.23687. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
157. Bradbury P.A., Shepherd F.A. Immunotherapy for Lung Cancer. J. Thorac. Oncol. 2008;3:S164–S170. doi: 10.1097/JTO.0b013e318174e9a7. [PubMed] [CrossRef] [Google Scholar]
158. Bovier P.A. Epaxal®: A virosomal vaccine to prevent hepatitis A infection. Expert Rev. Vaccines. 2008;7:1141–1150. doi: 10.1586/14760584.7.8.1141. [PubMed] [CrossRef] [Google Scholar]
More information: “Self-healing microcapsules synergetically modulate immunization microenvironments for potent cancer vaccination” Science Advances (2020). DOI: 10.1126/sciadv.aay7735



{kind=link}
{kind=link}