










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A new, hair-sprouting dollop of human skin created in the lab might one day help prevent hair loss.
Organoids are small, lab-grown cell groupings are designed to model real-world organs -in this case, skin.
A paper published in Nature describes the hairy creation as the first hair-baring human skin organoid made with pluripotent stem cells, or the master cells present during early stages of embryonic development that later turn into specific cell types.
The hirsute organoid’s development was led by Karl Koehler, Ph.D., formerly of Indiana University School of Medicine and now at Boston Children’s Hospital. An Oregon Health & Science University graduate student, Benjamin Woodruff, contributed by helping make the organoids as a post-baccalaureate research technician in the Stanford University lab of Stefan Heller, Ph.D.
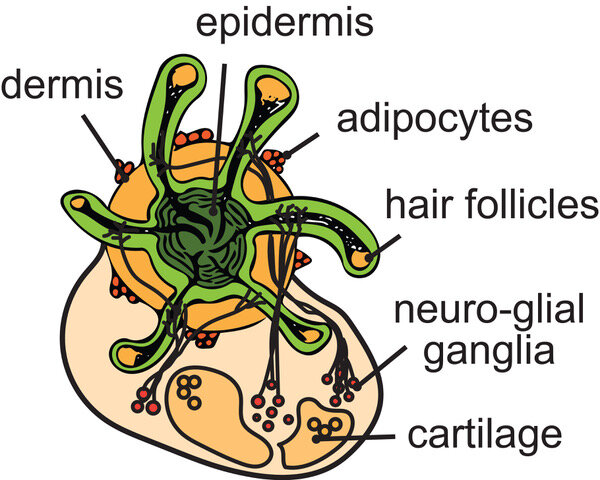
“This makes it possible to produce human hair for science without having to take it from a human,” explained Woodruff, who now is completing his first year of cell and developmental biology graduate studies at OHSU.
“For the first time, we could have, more or less, an unlimited source of human hair follicles for research.”
Having access to more hair-growing skin can help researchers better understand hair growth and development – and maybe even provide clues needed to reverse a retreating hair line.
Steps toward human hair follicle regeneration. Cell isolation: cell sources for bioengineering can be follicular (bulge stem cells, dermal papilla, and dermal sheath cells) and non‐follicular (keratinocytes, skin‐derived progenitors, and mesenchymal stem cells). Cell expansion: mesenchymal and epithelial cell sources are cultured in vitro.
Bioengineering: cell clustering in 3D instructive hair bulbs. Implantation: bulbs could generate functional hair follicles.

Hair loss (alopecia) affects a growing number of people worldwide. Limited efficacy and side effects of current pharmacological and surgical treatments have fostered the search for alternative therapeutic solutions. Great expectation has been recently put on hair follicle bioengineering, which is based on the development of functional hair follicles from an expanded population of hair‐inductive cells. However, human follicle neogenesis resorting to patient’s cells was not successfully achieved yet. Based on recent advances in the field, this review on cell‐based hair follicle tissue engineering systematically compiles the emerging strategies while disclosing the hurdles that still limit translation into the clinics.
Hair loss (alopecia) is a disease that affects a growing number of people worldwide and impacts individuals’ physical, psychological, and social well‐being.1 Patients with hair disorders suffer from emotional stress, embarrassment, and depression that severely compromise their life quality.2
Up to date, treatments include pharmacological and surgical (autologous hair transplant) interventions. Although hair restoration surgery is nowadays the most effective method, donor hair follicles (HFs) scarcity is often its major limitation.3
Besides, pharmacological treatments still not fully satisfy the patient’s needs and entail drastic side effects.4 Thus, the limited efficacy and possible side effects of the current treatments have fostered the search for alternative therapeutic solutions, capable of generating unlimited number of HFs de novo.
Noteworthy, stem cell‐based tissue engineering is emerging as the most thriving approach, aiming to reconstruct HFs in vitro to replace lost or damaged HFs as a consequence of disease, injury, or aging.
HF bioengineering approaches are based on the accumulated knowledge on reciprocal epithelial‐mesenchymal (EM) interactions controlling embryonic organogenesis and postnatal HF cyclic growth.
However, despite recent progress in the field, clinical applications of tissue engineering strategies for hair loss are still missing. Neogenesis of human follicles derived from cultured HF dermal cells has not been successfully achieved yet.
This review focus on the research approaches being developed to tackle the major limitations of human HF bioengineering, namely the loss of cellular function following in vitro HF cells expansion, the loss of in vivo tissue context/architecture, and the reconstruction of autologous functional HF germs for clinical procedures.
HF MORPHOGENESIS AND CYCLING: STEM CELL POPULATIONS
HF is a mini‐organ that forms during embryonic skin development. Its functional and cycling activities rely on a coordinated communication between the different cell populations from epithelial, mesenchymal, and neural crest stem cell origin,5 which additionally regulates adult skin homeostasis and wound repair.6, 7
Therefore, understanding the HF anatomy, as well as the stem‐cell populations operating during postnatal cyclic regeneration, is crucial for tissue engineering‐based solutions.
Follicular dermal stem cells exist in the dermis (skin‐derived precursors, SKP) able to regenerate dermal sheath (DS), and populate the dermal papilla (DP) at every growth cycle.8
Both DS and DP comprise mesenchymal cells with multi‐lineage differentiation capacity.9 In the mature HF, the DP is adjoined to connective tissue sheath (DS), together forming the dermal component of the mature HF10 (Figure (Figure1).1).
The DP is thought to be a master regulator of HF cycling, which consists in serial phases of growth (anagen), apoptotic‐driven regression (catagen), and rest (telogen).11 On the human scalp, anagen lasts 1‐6 years and it involves the complete regeneration of the cycling portion of the HF (Figure (Figure1).1).
At the telogen‐to‐anagen transition, DP stimulates epithelial hair follicle stem cells (HFSC) from the bulge region, which are adult multipotent cells holding self‐renewal capability and kept quiescent in their niche surrounded by the sebaceous gland (SG) in the outer root sheath.12, 13
When DP stimulatory signaling overcomes the threshold imposed by the inhibitory bulge microenvironment,14 HFSCs divide generating a new pool of progenitors at the bulge base called the secondary germ cells,15 which survive catagen‐driven apoptosis.16
These primed hair germ cells migrate to the bulb, while expanding and differentiating into transit‐amplifying cells (HF‐TACs) that attach to the basement membrane surrounding the DP lower half. HF‐TACs likely sit in place throughout much of anagen to fuel HF growth by differentiating into eight distinct epithelial lineages (eg, shaft, inner root sheath, and companion layer cells) and SGs17, 18, 19 (Figure1).

Cell population dynamics during hair follicle (HF) morphogenesis and cycling stages. Interactions between epithelial and mesenchymal cell populations in the skin determine the embryonic morphogenetic stages of induction, organogenesis and cytodifferentiation, as well as the postnatal hair cycling stages of anagen, catagen, and telogen. Inset on the right depicts the complex cell population dynamics operating during anagen. Bulge quiescent hair follicle stem cells (HFSCs) are activated by dermal papilla (DP) stimulatory signals to proliferate and generate HF‐TACs (HF‐TACs). HF‐TACs migrate out of the bulge and differentiate either in outer root sheath (ORS) or epithermal progenitors. HF‐TACs that migrate into the hair matrix give rise to several differentiated epithelial cell lineages (in different colors) that directly contribute to hair growth. Besides HFSCs, melanocyte stem cells (MeSC) in the bulge give rise to differentiated melanocytes that fuel pigment granules to the adjacent differentiating cells. Dashed lines represent cell trajectories and solid lines represent molecular signaling
In addition to HFSCs, melanocyte stem cells also reside in the bulge. During anagen, they are coordinately activated with HFSCs to generate mature melanocytes that produce and distribute pigment granules to the adjacent differentiating cells to form pigmented hair fibers.20 Catagen phase can last between 4 and 6 weeks, where keratinocytes and melanocytes undergo apoptotic processes.
This apoptotic‐driven regression causes DP to move upward, bringing it closer to the epithelial bulge.21 Following complete regression, HF enters a quiescent phase (telogen), which can last several months. The replacement of the old hair shaft fiber by the forming club fiber at the end of telogen is called “exogen.”22
Finally, HF macroenvironment also encompasses adipocyte tissue containing adipose‐derived stem cells in close commitment with hair growth regulation, as patented by increased adipose tissue thickness in anagen.6
Altogether, the DP, differentiated epithelial cells and the hair matrix constitute the cycling portion of the postnatal HF, being actively renewed each HF cycle. Conversely, the upper portion of the follicle (including bulge, SG appendage, and the infundibular epidermis) constitute the permanent portion of the postnatal HF, being formed during embryonic development and kept throughout life.23, 24
Any aberrant signaling affecting the communication between mesenchymal DP and the surrounding epithelial cells will disrupt the hair cyclic regeneration during postnatal life.
HF BIOENGINEERING: CELL SOURCES AND CHALLENGES
Stem cell‐based regenerative medicine is emerging as the most thriving approach for hair loss treatment by holding the potential of HF cloning, that is, the production of bioengineered instructive germs from human HF cells expanded in vitro to generating fully functional HFs upon transplant into the patient’s bald scalp.
Rationally, such a regenerative therapy may only be possible if combining receptive‐epithelial and inductive‐mesenchymal populations to mimic the well‐orchestrated interactions controlling lifelong HF cycles, which are deeply affected during hair loss.25
Ideally, a cell‐based regenerative medicine therapy would be autologous, that is, resort to patients’ cells derived from small amounts of tissue biopsies (eg, HF punch). Thus, researchers in the field have been mainly focused on developing therapeutic bioengineering solutions using dissociated HFSCs and DP cells (DPCs) isolated from HF biopsies. HFSCs and DPCs retrieved from nonbalding scalp follicles should first be expanded in culture to produce bioengineered structures in vitro with hair regenerative potential.
Still, an allogeneic cell source could be alternatively used for HF regenerative therapy. Two decades ago, transgender transplantation of microdissected DP and DS was shown to successfully induce HFs.26 This study not only pointed to the need of an inductive dermal component for HF regeneration, but it also disclosed the possibility of using an allogeneic cell source for therapy.
Indeed, HF proved to be an immune‐privileged site, as it does not express MHC (major histocompatibility complex) class I antigens.27, 28 Regardless of autologous vs allogeneic therapy, the relevance of HFSCs, and mainly DPCs, on tissue‐engineering approaches for treating alopecia has been the focus of intensive research over the last decade.
REFERENCES
1. Marks DH, Penzi LR, Ibler E, et al. The medical and psychosocial associations of alopecia: recognizing hair loss as more than a cosmetic concern. Am J Clin Dermatol. 2018;20:195‐200. [PubMed] [Google Scholar]
2. Hadshiew IM, Foitzik K, Arck PC, et al. Burden of hair loss: stress and the underestimated psychosocial impact of telogen effluvium and androgenetic alopecia. J Investig Dermatol. 2004;123:455‐457. [PubMed] [Google Scholar]
3. Rose PT. Hair restoration surgery: challenges and solutions. Clin Cosmet Investig Dermatol. 2015;8:361‐370. [PMC free article] [PubMed] [Google Scholar]
4. Talavera‐Adame D, Newman D, Newman N. Conventional and novel stem cell based therapies for androgenic alopecia. Stem Cells Cloning. 2017;10:11‐19. [PMC free article] [PubMed] [Google Scholar]
5. Hardy MH. The secret life of the hair follicle. Trends Genet. 1992;8:55‐61. [PubMed] [Google Scholar]
6. Rompolas P, Greco V. Stem cell dynamics in the hair follicle niche. Semin Cell Dev Biol. 2014;25–26:34‐42. [PMC free article] [PubMed] [Google Scholar]
7. Ito M, Liu Y, Yang Z, et al. Stem cells in the hair follicle bulge contribute to wound repair but not to homeostasis of the epidermis. Nat Med. 2005;11:1351. [PubMed] [Google Scholar]
8. Rahmani W, Abbasi S, Hagner A, et al. Hair follicle dermal stem cells regenerate the dermal sheath, repopulate the dermal papilla, and modulate hair type. Dev Cell. 2014;31:543‐558. [PubMed] [Google Scholar]
9. Jahoda CAB, Whitehouse CJ, Reynolds AJ, et al. Hair follicle dermal cells differentiate into adipogenic and osteogenic lineages. Exp Dermatol. 2003;12:849‐859. [PubMed] [Google Scholar]
10. Millar SE. Molecular mechanisms regulating hair follicle development. J Invest Dermatol. 2002;118:216‐225. [PubMed] [Google Scholar]
11. Cotsarelis G. Epithelial stem cells: a folliculocentric view. J Invest Dermatol. 2006;126:1459‐1468. [PubMed] [Google Scholar]
12. Fuchs E. The tortoise and the hair: slow‐cycling cells in the stem cell race. Cell. 2009;137:811‐819. [PMC free article] [PubMed] [Google Scholar]
13. Rompolas P, Deschene ER, Zito G, et al. Live imaging of stem cell and progeny behaviour in physiological hair‐follicle regeneration. Nature. 2012;487:496‐499. [PMC free article] [PubMed] [Google Scholar]
14. Woo WM, Zhen HH, Oro AE. Shh maintains dermal papilla identity and hair morphogenesis via a Noggin‐Shh regulatory loop. Genes Dev. 2012;26:1235‐1246. [PMC free article] [PubMed] [Google Scholar]
15. Ito M, Kizawa K, Hamada K, et al. Hair follicle stem cells in the lower bulge form the secondary germ, a biochemically distinct but functionally equivalent progenitor cell population, at the termination of catagen. Differentiation. 2004;72:548‐557. [PubMed] [Google Scholar]
16. Panteleyev AA, Jahoda CA, Christiano AM. Hair follicle predetermination. J Cell Sci. 2001;114:3419‐3431. [PubMed] [Google Scholar]
17. Greco V, Chen T, Rendl M, et al. A two‐step mechanism for stem cell activation during hair regeneration. Cell Stem Cell. 2009;4:155‐169. [PMC free article] [PubMed] [Google Scholar]
18. Yang H, Adam RC, Ge Y, et al. Epithelial‐mesenchymal micro‐niches govern stem cell lineage choices. Cell. 2017;169:483‐496.e413. [PMC free article] [PubMed] [Google Scholar]
19. Lim YS, Harland DP, Dawson TL Jr. Wanted, dead and alive: why a multidisciplinary approach is needed to unlock hair treatment potential. Exp Dermatol. 2019;28:517‐527. [PubMed] [Google Scholar]
20. Slominski A, Wortsman J, Plonka PM, et al. Hair follicle pigmentation. J Invest Dermatol. 2005;124:13‐21. [PMC free article] [PubMed] [Google Scholar]
21. Cotsarelis G, Sun T‐T, Lavker RM. Label‐retaining cells reside in the bulge area of pilosebaceous unit: implications for follicular stem cells, hair cycle, and skin carcinogenesis. Cell. 1990;61:1329‐1337. [PubMed] [Google Scholar]
22. Higgins CA, Westgate GE, Jahoda CA. From telogen to exogen: mechanisms underlying formation and subsequent loss of the hair club fiber. J Invest Dermatol. 2009;129:2100‐2108. [PubMed] [Google Scholar]
23. Sennett R, Rendl M. Mesenchymal–epithelial interactions during hair follicle morphogenesis and cycling. Semin Cell Dev Biol. 2012;23:917‐927. [PMC free article] [PubMed] [Google Scholar]
24. Paus R, Foitzik K. In search of the “hair cycle clock”: a guided tour. Differentiation. 2004;72:489‐511. [PubMed] [Google Scholar]
25. Garza LA, Yang CC, Zhao T, et al. Bald scalp in men with androgenetic alopecia retains hair follicle stem cells but lacks CD200‐rich and CD34‐positive hair follicle progenitor cells. J Clin Invest. 2011;121:613‐622. [PMC free article] [PubMed] [Google Scholar]
26. Reynolds AJ, Lawrence C, Cserhalmi‐Friedman PB, et al. Trans‐gender induction of hair follicles. Nature. 1999;402:33‐34. [PubMed] [Google Scholar]
27. Westgate GE, Craggs RI, Gibson WT. Immune privilege in hair growth. J Invest Dermatol. 1991;97:417‐420. [PubMed] [Google Scholar]
28. Christoph T, Muller‐Rover S, Audring H, et al. The human hair follicle immune system: cellular composition and immune privilege. Br J Dermatol. 2000;142:862‐873. [PubMed] [Google Scholar]
More information: Jiyoon Lee et al, Hair-bearing human skin generated entirely from pluripotent stem cells, Nature (2020). DOI: 10.1038/s41586-020-2352-3



{kind=link}