











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



In Sci-Fi movies, astronauts often enter an inactive state in “hibernation chambers” to cross the vastness of space.
This could cut down on the required amount of food and oxygen and to prevent serious side effects from low gravity, such as muscle wasting in zero-G condition.
A state of unconsciousness could also potentially minimize psychological challenges in space. Could humans hibernate in the future?
Why do some animals hibernate while others do not?
Do all animals have the potential to hibernate even if they never do so in nature?
Researchers from the University of Tsukuba in Japan opened the door to answering these questions by finding specific cells in the mouse brain that can trigger a hibernation-like state when activated. The study was published in the scientific journal Nature.
Animals usually enter hibernation when food becomes scarce in the winter. Their metabolism slows down, and their body temperature drops to a new set-point. This is like lowering the temperature on your thermostat in the winter–it reduces the amount of energy needed to maintain the body.
Along with a slower metabolism and a new set-point comes slower heart rate, weaker breathing, and less brain activity. Importantly, when animals come out of hibernation, their body and organs are healthy, even if they have lost a little weight.
Even though mice do not hibernate, researchers led by Takeshi Sakurai at the University of Tsukuba and Genshiro Sunagawa at the RIKEN Center for Biosystems Dynamics Research show that activating a specific type of cell in the mouse brain–dubbed Q neurons–caused them to enter a hibernation-like state for several days.
“The mice exhibited distinctive qualities that met the criteria for hibernation,” notes Sakurai. “In particular, the body temperature set-point lowered from about 96.8°F [36°C] to about 81°F [27°C], and the body functioned normally to maintain a lower body temperature around 22°C, even when the surrounding ambient temperature was dramatically reduced.”
The mice also showed all the signs of a reduced metabolism that are common during hibernation, including reduced heart rate, oxygen consumption, and respiration.
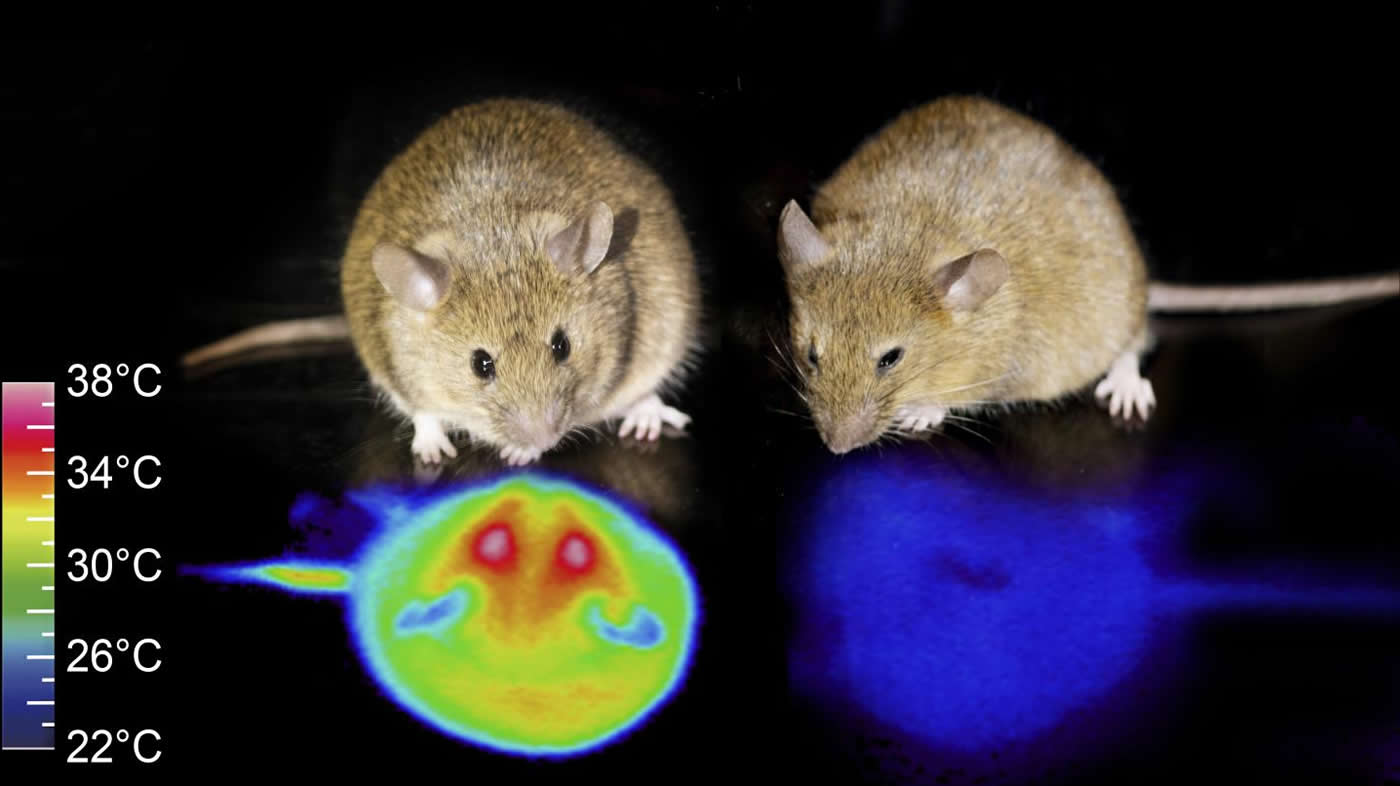
Being able to send mice into a hibernation-like state for days simply by artificially exciting Q neurons was somewhat unexpected.
“Even more surprising,” says first author Tohru Takahashi, “is that we were able to induce a similar hypometabolic state in rats, a species that neither hibernates nor has daily torpor.”
Although we do not know the answer yet, the possibility that humans also have Q neurons that can be used to induce a similar state is tantalizing.
“People might not want to hibernate for the same reasons as animals,” explains Sunagawa. “But there are medical reasons for wanting to place people in suspended animation, such as during emergency transport or critically ill conditions as in severe pneumonia, when the demand for oxygen cannot meet the supply.”
Sparing oxygen is not only for medicine. “In the future,” Sakurai added, “we may put human in a hibernation-like state for missions to Mars and beyond.”
Introduction
Hibernation is an adaptation to extreme environmental conditions, including cold temperatures and reduced food availability. In small mammalian hibernators, it is characterized by reduced metabolism, by severely decreased heart and respiratory rates and by dramatic lowering of body temperatures to few degrees above ambient temperature (Nedergaard and Cannon, 1990; Nurnberger, 1995).
Also, the brain temperature drops to 2–3°C above ambient temperature (Strumwasser, 1959). Still, hibernators are able to arouse from the state of torpor without any damage. The mechanisms underlying the entrance into and arousal from torpor state are not fully clear. However, the central nervous system (CNS) is supposed to be a fundamental key player in its regulation (Heller, 1979).
This presumes that brain function is maintained under extreme hypothermia during deep torpor, though the membrane potential of neurons, the properties of ion channels and receptors, the efficiency of synaptic transmission and the conduction velocity of neuronal activity are strongly dependent on temperature, and significant, functional disruptions are already observed few degrees below physiological tissue temperature (Hodgkin and Huxley, 1952; Westerfield et al., 1978; Schiff and Somjen, 1985; Kufel and Wojcik, 2018).
How does the brain of a hibernator deal with the low levels of brain temperatures?
What changes in neuronal signal processing occur when the brain temperature drops to values near or below freezing point? Various approaches have been used to unravel the physiology of neurons and synapses in the hibernating brain.
We presently provide a compact overview on in vivo and in vitro electrophysiological data as well as histochemical, molecular and biochemical results comprising the evaluation of activity markers.
We refer to both earlier and more recent data, thereby focusing on the temperature-dependency of neuronal activity during hibernation.
Structural plasticity which underlies functional changes (e.g., synapse and spine degradation, protein synthesis) is not considered here, but was intensively discussed in two recent reviews (Arendt and Bullmann, 2013; Horowitz and Horwitz, 2019).
Electrophysiological Characterization of Neuronal Activity in the Hibernating Brain in vivo
Activity of Single-Units and Neuronal Populations
Brain activity was reported to dramatically change during hibernation.
Electroencephalograms (EEGs) that serve as a measure of synchronous firing of a population of neurons were nearly isoelectric (i.e., no significant changes in voltages) in hibernating ground squirrels and hamsters at lowest body temperature during torpor state (Chatfield et al., 1951; Mihailovic, 1972).
Single-unit recordings obtained by chronically implanted electrodes in the lateral posterior thalamus and posterior thalamic nuclear group of ground squirrels confirmed these observations: firing rates of spontaneous neuronal activity were systematically reduced with decreasing body temperature, and neurons eventually stopped firing at body temperatures of 15–18°C, remained silent for 10–28 h (deep torpor), and only began firing again when body temperature increased (Krilowicz et al., 1988).
Along with the reduction in firing rate, the waveform of the action potential dramatically changed with amplitude becoming smaller and spike width being increased at lower body temperatures (Krilowicz et al., 1989). These data imply that neuronal activity in hibernating animals is highly dependent on body and brain temperature.
Activity of Sensory Systems and Arousal
Although electrical activity is greatly reduced and almost silent at the lowest body temperature in torpor, this does not mean that information processing is suspended. Rather, sensory stimulation was shown to elicit specific responses in the CNS, even at very low body temperatures (<10°C).
Peripheral stimulation of hibernating hamsters led to electrical responses in the cerebral cortex at a body temperature of 9.1°C (Chatfield et al., 1951). Similar observations were made in the ground squirrel where touching the torpid animal with a glass rod induced activity in the brainstem reticular formation, partly followed by arousal of the animals (Strumwasser, 1959).
Also, acoustic stimulation of ground squirrels was reported to evoke a behavioral response or even a complete arousal (Strumwasser, 1959). Auditory brainstem recordings (ABRs) in ground squirrels during arousal revealed that acoustic stimulation indeed elicits stimulus-driven responses even at body temperatures as low as 9.8°C (Hamill et al., 1989).
In the woodchuck, auditory stimulation resulted in specific ABR responses, although only waves 1 and 2 (representing the 8th nerve and the cochlear nucleus) were detectable at lowest body temperature (Katbamna et al., 1992). These data imply that peripheral nerves and primary central brain regions might be more resistant to hypothermia-induced activity changes compared to higher central brain regions.
Regional Pattern of Activity Changes
Reduction of brain activity as the animal enters torpor follows a regional pattern, with EEG activity being reduced first in the neocortex and then in the reticular formation, followed by thalamocortical structures and the limbic system, with activity ceasing last in the hippocampus, if at all.
During arousal, activity reappears and increases in reverse order: i.e., first in the limbic system (hippocampus) and then in the hypothalamus, thalamus, reticular formation, and cortex (South et al., 1968; Mihailovic, 1972; Heller, 1979).
Notably, the neocortex is the first structure where activity is depressed during entrance into torpor and the last to regain activity on arousal, a finding that has focused studies on information processing during torpor to subcortical brain regions.
Taken together, in vivo studies (Figure 1) convincingly show that although neuronal activity is attenuated throughout the brain in torpor, some brain regions are more active than others.
These data led to the hypothesis that the limbic system-midbrain circuitry (interconnections between the limbic system, hypothalamus and brainstem reticular formation) contributes to neural control of entrance into and arousal from torpor (Heller, 1979; Beckman and Stanton, 1982).

Source:
University of Tsukuba



{kind=link}