











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")


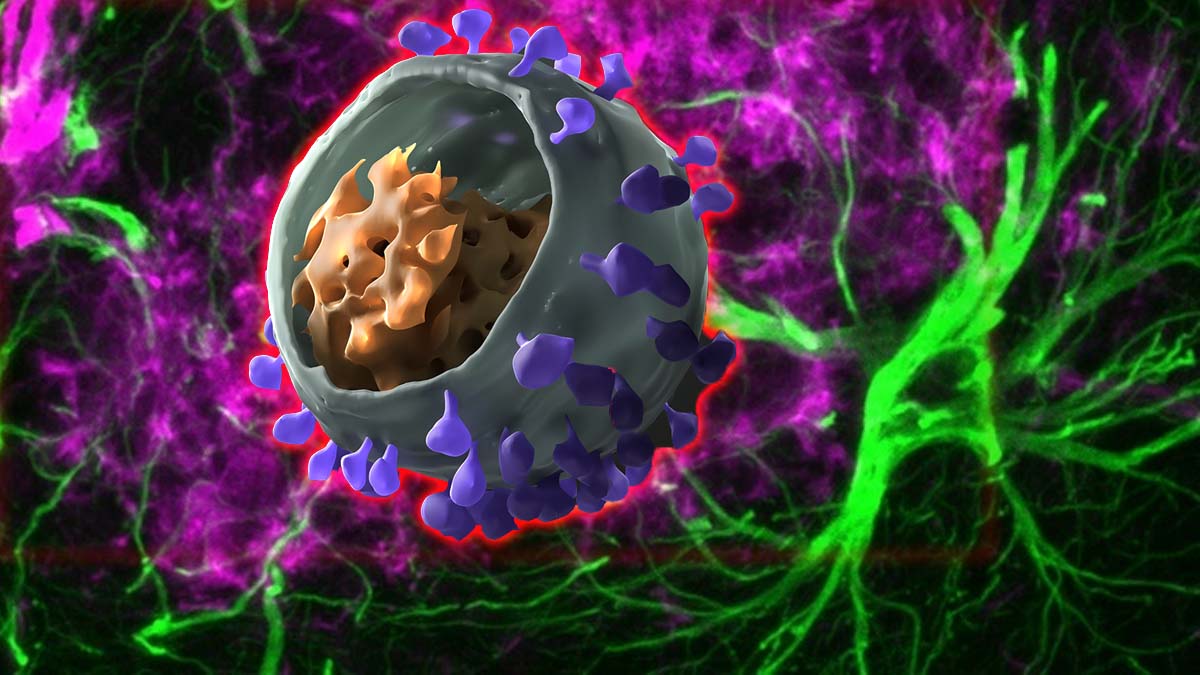
Researchers have found that astrocytes, a type of brain cell can harbor HIV and then spread the virus to immune cells that traffic out of the brain and into other organs.
HIV moved from the brain via this route even when the virus was suppressed by combination antiretroviral therapy (cART), a standard treatment for HIV. The study, conducted by researchers at Rush University Medical Center in Chicago and published in PLOS Pathogens, was funded by the National Institutes of Health.
“This study demonstrates the critical role of the brain as a reservoir of HIV that is capable of re-infecting the peripheral organs with the virus,” said Jeymohan Joseph, Ph.D., chief of the HIV Neuropathogenesis, Genetics, and Therapeutics Branch at NIH’s National Institute of Mental Health, which co-funded the study.
“The findings suggest that in order to eradicate HIV from the body, cure strategies must address the role of the central nervous system.”
HIV attacks the immune system by infecting CD4 positive (CD4+) T cells, a type of white blood cell that is vital to fighting off infection. Without treatment, HIV can destroy CD4+ T cells, reducing the body’s ability to mount an immune response – eventually resulting in AIDS.
cART, which effectively suppresses HIV infections, has helped many people with HIV live longer, healthier lives. But some studies have shown that many patients receiving antiretroviral drugs also show signs of HIV-associated neurocognitive disorders, such as thinking and memory problems. Researchers know that HIV enters the brain within eight days of infection, but less is known about whether HIV-infected brain cells can release virus that can migrate from the brain back into the body to infect other tissues.
The brain contains billions of astrocytes, which perform a variety of tasks — from supporting communication between brain cells to maintaining the blood-brain barrier.
To understand whether HIV can move from the brain to peripheral organs, Lena Al-Harthi, Ph.D., and her research team at Rush University Medical Center transplanted HIV-infected or noninfected human astrocytes into the brains of immunodeficient mice.
The researchers found that the transplanted HIV-infected astrocytes were able to spread the virus to CD4+ T cells in the brain. These CD4+ T cells then migrated out of the brain and into the rest of the body, spreading the infection to peripheral organs such as the spleen and lymph nodes.
They also found that HIV egress from the brain occurred, albeit at lower levels, when animals were given cART. When cART treatment was interrupted, HIV DNA/RNA became detectable in the spleen — indicating a rebound of the viral infection.

“Our study demonstrates that HIV in the brain is not trapped in the brain — it can and does move back into peripheral organs through leukocyte trafficking,” said Dr. Al-Harthi. “It also shed light on the role of astrocytes in supporting HIV replication in the brain — even under cART therapy.”
This information has significant implications for HIV cure strategies, as such strategies need to be able to effectively target and eliminate reservoirs of HIV replication and reinfection, Dr. Al-Harthi added.
“HIV remains a major global public health concern, affecting 30 to 40 million people across the globe. To help patients, we need to fully understand how HIV affects the brain and other tissue-based reservoirs,” said May Wong, Ph.D., program director for the NeuroAIDS and Infectious Diseases in the Neuroenvironment at the NIH’s National Institute of Neurological Disorders and Stroke, which co-funded the study.
“Through additional studies that replicate these findings are needed, this study brings us another step closer towards that understanding.”
HIV-1 latent infection in the CNS
The formation of a latent reservoir allows HIV-1 to persist throughout a patient’s lifetime, even while adhering to ART. Several cellular reservoirs have been characterized to harbor low-level chronic HIV-1 infection, which contributes to a barrier for a sterile HIV-1 cure.
Examples of these reservoirs include: resting memory CD4+ T-lymphocytes, microglia, monocytes, macrophages, and astrocytes. Out of all these reservoirs, the resting memory CD4+ T-lymphocytes contain the highest HIV-1 load.
During the initial stages of CNS infiltration, HIV-1 predominantly infects perivascular macrophages, microglia, and astrocytes. These CNS cells satisfy most of the criteria for an HIV-1 reservoir, as described by Blankson et al.46.
For example, these cells have been shown to harbor integrated HIV-1 provirus, as demonstrated through methods such as in situ hybridization and laser capture microdissection coupled with PCR47.
The perivascular macrophages have a half-life of about 3 months48, while microglial cells and astrocytes have a half-life ranging from several months to years, potentially upward of an entire lifetime49.
Furthermore, these cells harbor the specific molecular machinery to facilitate the HIV-1 provirus into a latent state. For example, the CNS-derived HIV-1 long terminal repeat (LTR) promoter has lower basal transcriptional activity due to mutations in the core promoter region50.
Robust suppression of SIV LTR activity has also been corroborated in SIV infection models, as IFN-β induced the expression of a dominant-negative form of C/EBP-β in the CNS51. In HIV-1 postmortem brain specimens, CTIP2, HP1, MeCP2, and HDAC1 levels were all elevated, giving rise to the possibility that they may play a role in HIV-1 transcriptional silence in the CNS52.
Since astrocytes are the most abundant cell type in the CNS, their roles in HAND have been extensively studied. It remains controversial whether astrocytes can be productively and/or latently infected by HIV-1.
Early immunohistochemical studies using postmortem brain tissue from HAD patients demonstrated the presence of HIV-1 proteins in hippocampal astrocytes53. The highly sensitive PCR analysis of laser-captured single astrocytes revealed HIV-1 DNA in up to 19% of astrocytes from HAD patients54. Even in asymptomatic individuals, HIV-1 DNA was detected in laser-captured astrocytes47.
However, most in vivo studies only demonstrated the detection of early HIV-1 transcripts, such as Nef55. In vitro studies showed contradicting conclusions. For example, while primary astrocytes or astrocytic cell lines demonstrated either productive56 or non-productive57 infection, Boutet et al. reported that astrocytes remained completely resistant to HIV-1 infection58.
Nevertheless, persistent HIV-1 infection in astrocytic cell lines59 or primary human astrocytes60 may contribute to the pathogenesis of HAND in the ART era60. Endocytosis56 and cell-to-cell interactions44 have been identified as possible mechanisms of HIV-1 entry into astrocytes.
The concept of latent HIV-1 infection in astrocytes has been also controversial. Early studies in stable cell lines after HIV-1 cDNA transfection demonstrated latency and reactivation-like properties61. Subsequent studies with latency-reversing agents showed the epigenetic regulation of HIV-1 latency in primary astrocytes57 and human neural stem cells (NSC)-derived astrocytes
Recent reports demonstrated that human astrocytes sustain long-term productive HIV-1 infection without the establishment of reversible viral latency60. Although the frequency of HIV-infected astrocytes is very low, the large number of total astrocytes in the brain, cell-to-cell transmission, and long-term HIV-1 infection may justify the important contribution of long-lived astrocytes to HAND persistence.
Outstanding questions and future directions
Collectively, the literature shows that adult neurogenesis is compromised in HIV-1 infected patients in both the pre- and post-ART eras90,110. NSCs are capable of being productively and latently infected by HIV-1.
It is likely that the integrated proviral DNA may persist as the NSCs differentiate into neurons, astrocytes, or oligodendrocytes. Or, the latent provirus may be reactivated as the NSCs differentiate into rapidly amplifying NPCs, spreading the infection to neighboring infiltrating immune cells and astrocytes.
This secondary infection could re-seed the latent reservoir in NSCs or damage the NSCs and their progeny cells (Fig. 2).

Working Model for Neural stem cells as an HIV-1 Reservoir. Productive infection of neural stem cells by HIV-1 results in integration of the proviral genome in these cells. As these neural stem cells differentiate into neural progenitor cells, the integrated provirus robustly reactivates and generates new virions. Neuroblasts and/or glioblasts could potentially harbor the integrated proviral genome, which could lead to viral integration in neurons, astrocytes, and oligodendrocytes. This new HIV-1 virus further infects microglia and infiltrating immune cells in the brain, which subsequently infects astrocytes. Astrocytic infection could result from direct cell-to-cell contact with infected immune cells or from stimulation from inflammatory cytokines, which could upregulate the receptors needed for HIV-1 infection. These astrocytes would release viral proteins that damage neuron, astrocyte, and oligodendrocyte function. In addition, these secreted virions could subsequently infect other neural stem cells, and re-seed that specific reservoir for further HIV-1 latency establishment.
The HIV-1 proteins gp120 and Tat exert differential effects on adult neurogenesis at the molecular, cellular, and neurobehavioral levels. However, most of the reported studies focused on single viral proteins (Tat or gp120) in a specific cellular type (GFAP+ astrocytes and/or NSCs). HIV-1 patients in the ART era have the entire HIV-1 proviral genome integrated into their cells.
To mimic the persistent and latent HIV-1 infection during the ART era, two rodent models have been generated: the HIV-1 Tg26 mouse111 and the HIV-1 transgenic rat112. Both models contain multiple copies of a replication-deficient HIVNL4–3 strain integrated into their cells and express various degrees of HIV-1 transcripts in different organs/tissues111,112, particularly in the neurogenic zones80.
Even though spatial learning and memory deficits have been demonstrated in the HIV-1 transgenic rat113, they have not been directly correlated with alterations of adult neurogenesis. Our recent in vitro and in vivo study demonstrated that HIV-1 Tg26 mice exhibited significant deficits in the early- and late-stage neurogenesis80.
It would be interesting to determine how neurogenesis is modulated in rodent models with acute or chronic infection (i.e., direct infection with Eco-HIV in wild-type mice or HIV-1 infection in humanized mouse models with engrafted human brain organoids).
Finally, it would be of importance to assess any gender-specific differences in adult neurogenesis in the context of HIV-1 infection. Given that women make up a majority of the HIV-1-infected population worldwide and clinical studies evaluating the incidence of HIV-1/HAND in male and female patients have been inconsistent114, modeling any possible gender-specific neurogenesis deficits in animal models of HIV-1 disease would significantly benefit the HAND research.
Finally, the effects of neurogenesis in the aging HAND population should be evaluated. Since the prevalence of HIV-1 infection in patients aged 50 and older is increasing, and hippocampal neurogenesis depletes with aging, understanding the role of aging in the development of HAND is of clinical relevance and significance.
REFERENCE
- Blankson JN, Persaud D, Siliciano RF. The challenge of viral reservoirs in HIV-1 infection. Annu Rev Med. 2002;53:557–93.
- Thompson KA, Cherry CL, Bell JE, McLean CA. Brain cell reservoirs of latent virus in presymptomatic HIV-infected individuals. Am J Pathol. 2011;179:1623–9.
- Koppensteiner H, Brack-Werner R, Schindler M. Macrophages and their relevance in human immunodeficiency virus Type I infection. Retrovirology. 2012;9:82.
- Sofroniew MV, Vinters HV. Astrocytes: biology and pathology. Acta Neuropathol. 2010;119:7–35.
- Gray LR, Cowley D, Crespan E, et al. Reduced basal transcriptional activity of central nervous system-derived HIV Type 1 long terminal repeats. AIDS Res Hum Retroviruses. 2013;29:365–70.
- Barber SA, Gama L, Dudaronek JM, et al. Mechanism for the establishment of transcriptional HIV latency in the brain in a simian immunodeficiency virus-macaque model. J Infect Dis. 2006;193:963–70.
- Desplats P, Dumaop W, Smith D, et al. Molecular and pathologic insights from latent HIV-1 infection in the human brain. Neurology. 2013;80:1415–23.
- Chompre G, Cruz E, Maldonado L, et al. Astrocytic expression of HIV-1 nef impairs spatial and recognition memory. Neurobiol Dis. 2013;49:128–36.
- Churchill MJ, Wesselingh SL, Cowley D, et al. Extensive astrocyte infection is prominent in human immunodeficiency virus-associated dementia. Ann Neurol. 2009;66:253–8.
- Ranki A, Nyberg M, Ovod V, et al. Abundant expression of HIV nef and rev proteins in brain astrocytes in vivo is associated with dementia. AIDS. 1995;9:1001–8.
- Chauhan A, Mehla R, Vijayakumar TS, Handy I. Endocytosis-mediated HIV-1 entry and its significance in the elusive behavior of the virus in astrocytes. Virology. 2014;456–457:1–9.
- Chauhan A. Enigma of HIV-1 latent infection in astrocytes: an in-vitro study using protein kinase C agonist as a latency reversing agent. Microbes Infect. 2015;17:651–9.
- Boutet A, Salim H, Taoufik Y, et al. Isolated human astrocytes are not susceptible to infection by M-and T-tropic HIV-1 strains despite functional expression of the chemokine receptors CCR5 and CXCR4. Glia. 2001;34:165–77.
- Dewhurst S, Sakai K, Bresser J, et al. Persistent productive infection of human glial cells by human immunodeficiency virus (HIV) and by infectious molecular clones of HIV. J Virol. 1987;61:3774–82.
- Barat C, Proust A, Deshiere A, et al. Astrocytes sustain long-term productive HIV-1 infection without establishment of reactivable viral latency. Glia. 2018;66:1363–81.
- Atwood WJ, Tornatore CS, Meyers K, Major EO. HIV-1 mRNA transcripts from persistently infected human fetal astrocytes. Ann N Y Acad Sci. 1993;693:324–5.
- Schneider M, Tigges B, Meggendorfer M, et al. A new model for post-integration latency in macroglial cells to study HIV-1 reservoirs of the brain. AIDS. 2015;29:1147–59.
80. Putatunda R, Zhang Y, Li F, et al. Adult neurogenic deficits in HIV-1 tg26 transgenic mice. J Neuroinflammation. 2018;15:287.
- Dickie P, Felser J, Eckhaus M, et al. HIV-associated nephropathy in transgenic mice expressing HIV-1 genes. Virology. 1991;185:109–19. [PubMed] [Google Scholar]
- Reid W, Sadowska M, Denaro F, et al. An HIV-1 transgenic rat that develops HIV-related pathology and immunologic dysfunction. Proc Natl Acad Sci U S A. 2001;98:9271–6.
- Vigorito M, LaShomb AL, Chang SL. Spatial learning and memory in HIV-1 transgenic rats. J Neuroimmune Pharmacol. 2007;2:319–28.
- Hestad KA, Menon JA, Silalukey-Ngoma M, et al. Sex differences in neuropsychological performance as an effect of human immunodeficiency virus infection: a pilot study in Zambia, Africa. J Nerv Ment Dis. 2012;200:336–42. [PMC free article]
Source:
NIH/NIMH



{kind=link}