










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Le persone anziane sono più soggette a malattie infettive poiché la funzione del loro sistema immunitario diminuisce continuamente con l’avanzare dell’età.
Ciò diventa particolarmente evidente durante le epidemie di influenza stagionale o l’insorgenza di altre malattie virali come COVID-19 . Poiché l’efficacia della vaccinazione negli anziani è fortemente ridotta, questa fascia d’età è particolarmente vulnerabile a tali patogeni infettivi e spesso mostra il più alto tasso di mortalità.
Oltre al declino immunitario legato all’età, gli individui anziani sono comunemente colpiti dalla fragilità che influisce negativamente sulla qualità della vita.
Sebbene l’aspettativa di vita media per gli umani continui a crescere, vivere più a lungo è spesso associato a problemi di salute legati all’età.
Identificato il ruolo importante del grasso della pancia nei processi di invecchiamento
I ricercatori del Dipartimento di ricerca biomedica (DBMR) e dell’Istituto di patologia dell’Università di Berna e dell’Ospedale universitario di Berna (Inselspital) hanno deciso di identificare nuovi approcci per migliorare la salute in una popolazione in rapida crescita.
Per molti anni gli scienziati hanno ipotizzato che l’infiammazione cronica di basso grado acceleri i processi di invecchiamento e lo sviluppo di disturbi legati all’età.
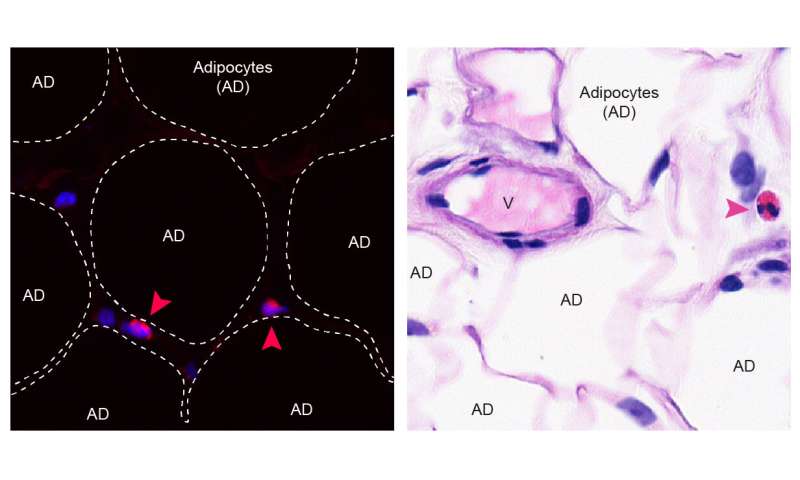
Un team internazionale di ricercatori sotto la guida bernese ha ora dimostrato che il tessuto adiposo viscerale , noto come grasso della pancia, contribuisce in modo decisivo allo sviluppo dell’infiammazione cronica di basso grado.
Lo scienziato attorno al dott. Mario Noti, precedentemente presso l’Istituto di patologia dell’Università di Berna e il dott. Alexander Eggel del Dipartimento di ricerca biomedica (DBMR) dell’Universität di Berna, hanno riferito che alcune cellule immunitarie nel grasso della pancia svolgono un ruolo essenziale ruolo nella regolazione dell’infiammazione cronica di basso grado e dei processi di invecchiamento a valle.
Potrebbero dimostrare che queste cellule immunitarie possono essere utilizzate per invertire tali processi. I risultati di questo studio sono stati pubblicati sulla rivista scientifica Nature Metabolism e sono stati ulteriormente evidenziati da un articolo editoriale di News and Views.
Grasso della pancia come fonte di infiammazione cronica
Il team attorno al Dr. Noti e al Dr. Eggel potrebbe dimostrare che un certo tipo di cellule immunitarie, note come eosinofili , che si trovano principalmente nella circolazione sanguigna, sono presenti anche nel grasso della pancia sia dell’uomo che dei topi.
Sebbene sia classicamente noto per fornire protezione dalle infezioni da parassiti e per promuovere la malattia allergica delle vie aeree, gli eosinofili situati nel grasso della pancia sono responsabili del mantenimento dell’omeostasi immunitaria locale.
Con l’aumentare dell’età la frequenza degli eosinofili nel grasso della pancia diminuisce, mentre aumenta il numero di macrofagi pro-infiammatori. A causa di questo squilibrio delle cellule immunitarie, il grasso della pancia si trasforma in una fonte di mediatori pro-infiammatori che si accumulano sistematicamente nella vecchiaia.
La terapia cellulare con eosinofili promuove il ringiovanimento
In una fase successiva, i ricercatori hanno studiato la possibilità di invertire le menomazioni legate all’età ripristinando l’equilibrio delle cellule immunitarie nel tessuto adiposo viscerale.
“In diversi approcci sperimentali, siamo stati in grado di dimostrare che i trasferimenti di eosinofili da topi giovani a soggetti anziani hanno risolto non solo un’infiammazione locale ma anche sistemica di basso grado”, afferma Dr Eggel.
“In questi esperimenti, abbiamo osservato che gli eosinofili trasferiti si stavano dirigendo selettivamente nel tessuto adiposo “, aggiunge il Dr. Noti. Questo approccio ha avuto un effetto ringiovanente sull’organismo invecchiato.
Di conseguenza, gli animali anziani hanno mostrato miglioramenti significativi nella forma fisica, come valutato dai test di resistenza e aderenza. Inoltre, la terapia ha avuto un effetto ringiovanente sul sistema immunitario che si manifesta in migliori risposte vaccinali di topi anziani.
Tradurre i risultati in cliniche
“I nostri risultati indicano che i processi biologici dell’invecchiamento e le alterazioni funzionali associate sono più plastiche di quanto precedentemente ipotizzato”, afferma il Dr. Noti. È importante sottolineare che anche i cambiamenti osservati correlati all’età nella distribuzione delle cellule immunitarie adipose nei topi sono stati confermati nell’uomo.
“Una direzione futura della nostra ricerca sarà ora quella di sfruttare le conoscenze acquisite per l’istituzione di approcci terapeutici mirati per promuovere e sostenere un invecchiamento in buona salute nell’uomo”, afferma Dr Eggel.
Neutrofili ed eosinofili
I neutrofili sono la popolazione più abbondante di globuli bianchi nell’uomo, svolgendo ruoli importanti nel processo infiammatorio e nella risposta immunitaria primaria [123]. Sono stati descritti principalmente come cellule immunitarie di breve durata e per il loro ruolo nella clearance dei patogeni extracellulari.
Tuttavia, questo concetto è stato superato con le scoperte della loro capacità di vivere molto più a lungo del previsto e di produrre diverse citochine chiave, chemochine e recettori Fc [124].
Oggi, l’evidenza ha dimostrato importanti contributi dei neutrofili sia nella promozione che nell’inibizione dello sviluppo e della crescita del tumore [125]. Questa dicotomia suggerisce che i neutrofili sono composti da diversi sottogruppi cellulari.
Tuttavia, l’identificazione di questi sottogruppi è stata banale a causa della mancanza di robusti marcatori di cellule superficiali [125], limitando la loro caratterizzazione di proprietà fisiche e funzionalità [126].
Si pensa che i neutrofili supportino la crescita e la progressione del tumore.
Un rapporto più elevato tra neutrofili e linfociti rappresenta un marcatore prognostico sfavorevole per una varietà di tipi di cancro come il carcinoma polmonare e mammario [126].
In effetti, i neutrofili possono supportare la colonizzazione metastatica in un modello murino di carcinoma mammario, orchestrando la formazione della nicchia premetastatica nel polmone [127]. I neutrofili sono reclutati nel microambiente tumorale da una serie di citochine CXC attraenti rilasciate dalle cellule tumorali [124].
Inoltre, i neutrofili sono in grado di sostenere la progressione del tumore attraverso il rilascio di fattori proangiogenici come VEGF e MMP9 [128,129] e, mantenendo un microambiente tumorale immunosoppressivo attraverso l’espressione dell’enzima che consuma arginina Arg1 e produzione ROS [130].
La deplezione di arginina e ROS inibiscono direttamente l’attività antitumorale delle cellule T e, pertanto, i neutrofili possono influire negativamente sull’efficacia dell’immunoterapia del cancro basata sulle cellule T (Figura 2).
Come accennato in precedenza, i neutrofili possono anche esercitare attività citotossica contro le cellule tumorali. Durante le prime fasi della formazione del tumore quando l’ambiente immunosoppressivo non è ancora completamente stabilito [125] i neutrofili riconoscono e uccidono le cellule tumorali attraverso l’ADCC.
I neutrofili contribuiscono al controllo del tumore e all’eradicazione del tumore anche regolando l’attivazione e la funzione delle cellule immunitarie adattive come le cellule T CD4 o CD8 [124,131]. Infatti, i neutrofili funzionano anche come APC e quindi possono interagire direttamente con le cellule T, supportando una robusta attività antitumorale delle cellule T. [132-136].
Al contrario, i neutrofili possono reclutare e attivare le cellule Treg in seguito alla secrezione di citochine come CCL17 e promuovendo un ambiente immunosoppressivo (Figura 2) [137]. Tuttavia, nel microambiente tumorale i neutrofili, così come i macrofagi, esercitano generalmente un’attività tollerogena e, di conseguenza, ostacolano l’efficacia dell’immunoterapia basata sulle cellule T.
In questo contesto, TGFβ può polarizzare i neutrofili verso un fenotipo immunosoppressivo e ostacolare l’infiltrazione del tumore delle cellule T CD8 [133]. Il blocco del TGFβ nel tumore aumenta l’infiltrazione delle cellule T CD8 e la risposta antitumorale dipendente dalle cellule T CD8 [133].
Al contrario, è stato dimostrato che l’IFNβ promuove l’attività antitumorale dei neutrofili e il reclutamento nel microambiente tumorale [125,138].
Il blocco della funzione immunosoppressiva dei neutrofili infiltranti il tumore ha dimostrato di migliorare l’efficacia delle immunoterapie basate sulle cellule T, come ICB e ACT.
In un modello murino di melanoma, l’inibizione del recettore tirosina chinasi c-MET (HGF-R, recettore del fattore di crescita degli epatociti) riduce il reclutamento dei neutrofili nel microambiente tumorale e nei linfonodi drenanti in risposta all’immunoterapia (Figura 2). Di conseguenza, l’inibizione di c-MET migliora la risposta a ICB e ACT [139].
Nel frattempo, l’inibizione della proteina 2 trasportatrice di acidi grassi (FATP2) abroga l’attività immunosoppressiva dei neutrofili e la risposta all’ICB [140]. Inoltre, in determinate condizioni di stress, i neutrofili possono rilasciare strutture a base di DNA, chiamate trappole extracellulari di neutrofili (NET), che consentono ai neutrofili di catturare e uccidere agenti patogeni extracellulari [141].
Questo fenomeno di morte cellulare è oggi chiamato NETosis [142] e rappresenta un meccanismo adottato dai neutrofili per modulare l’immunità adottiva delle cellule T. I NET sono in grado di interagire con i DC migliorandone la maturazione [143.144], determinando la sovraregolazione di molecole costimolatorie come CD40, CD80 / CD86 e MHCII e la proliferazione delle cellule T CD4 [145].
Inoltre, i NET possono innescare direttamente le cellule T portando ad una maggiore espressione di marcatori di attivazione come CD69 e CD25 [146]. In particolare, le cellule T innescate dai NET sono state in grado di rispondere a specifici antigeni anche in assenza di segnali costimolatori.
Vale la pena notare che questo scenario è comune nel microambiente tumorale e, pertanto, NETosis può essere sfruttato per indurre o modulare l’attivazione di cellule T specifiche del tumore.
Tuttavia, la neutrofilia e la formazione di NET supportano la progressione della malattia e la formazione di metastasi nel carcinoma polmonare e del colon e il blocco della NETosi ha comportato una riduzione della diffusione delle cellule tumorali [147]).
Inoltre, è stato dimostrato che i NET possono proteggere le cellule tumorali dalle cellule T e dalla citotossicità mediata dalle cellule NK [148]. Le cellule tumorali sono in grado di reclutare neutrofili e di sostenere la NETosi attraverso la secrezione di chemioattraenti di neutrofili, come i ligandi dei recettori delle chemochine CXCR1 e CXCR2 [149].
Le cellule tumorali rivestite con NET sono state fisicamente protette contro le cellule immunitarie adattive, compromettendo la motilità delle cellule T e delle cellule NK e impedendo loro di raggiungere il tumore. In particolare, l’inibizione farmacologica della NETosi sensibilizza i tumori alla terapia ICB [149].
Questa evidenza evidenzia le potenziali opportunità terapeutiche nel prendere di mira i neutrofili associati alle immunoterapie. Tuttavia, il contributo dei neutrofili allo sviluppo e alla progressione del tumore deve ancora essere completamente chiarito [124] e le terapie mirate ai neutrofili in grado di potenziare l’immunoterapia basata sulle cellule T non hanno ancora raggiunto applicazioni cliniche.
Gli eosinofili partecipano ai processi di riparazione dei tessuti e alla risposta immunitaria contro i parassiti. Inoltre, gli eosinofili svolgono un ruolo in diverse malattie come l’asma allergico, i disturbi autoimmuni e il cancro [150,151].
Sebbene gli eosinofili siano stati descritti nei campioni di tumore alla fine del XIX secolo e siano noti per infiltrarsi in diversi tipi di tumore [152], la loro funzione nella progressione del tumore e nel microambiente tumorale è ancora oggetto di indagine [153].
Gli studi hanno dimostrato un aumento della conta degli eosinofili sulla terapia con ICB nei pazienti con melanoma, carcinoma prostatico e polmonare che si correlava positivamente con la prognosi della malattia [154-157]. Si propone che il rilascio di DAMPS da parte delle cellule necrotiche nello stroma tumorale sia responsabile del reclutamento di eosinofili (eosinofilia) [158].
In effetti, l’infiltrazione di eosinofili nel microambiente tumorale di pazienti con melanoma trattati con ICB è correlata positivamente con un aumento dell’infiltrazione di TIL e un migliore risultato del paziente [159].
Gli eosinofili attivati sono in grado di rimodellare e normalizzare la vascolarizzazione del tumore, promuovendo l’infiltrazione delle cellule T e aumentando l’efficacia dell’ACT [160].
Inoltre, gli eosinofili possono supportare l’immunità antitumorale modulando la polarizzazione dei macrofagi. In particolare, le citochine proinfiammatorie come INFγ e TNFα sono prodotte da eosinofili attivati.
Pertanto, quando trasferiti insieme alle cellule T, gli eosinofili attivati promuovono la polarizzazione della TAM verso il fenotipo simil-M1, contribuendo alla creazione di un microambiente tumorale proinfiammatorio [160].
A causa delle loro proprietà di ricerca del tumore, gli eosinofili stanno diventando nuovi potenziali bersagli per migliorare le immunoterapie basate sulle cellule T. Nel modello preclinico murino di carcinoma epatico, prostatico e mammario, l’inibizione della dipeptidil-peptidasi 4 (DPP4) con il farmaco approvato dalla FDA sitagliptin migliora l’attività antitumorale mediata dagli eosinofili dell’ospite e contribuisce all’efficacia dell’ICB [161].
Questi studi promettenti possono spianare la strada a nuove strategie terapeutiche volte a migliorare l’immunoterapia basata sulle cellule T attraverso l’inibizione della DPP4 (Figura 2) [162].

Riferimento:
123. Mestas, J .; Hughes, CC di topi e non uomini: differenze tra topo e immunologia umana.
J. Immunol. 2004, 172, 2731–2738.
124. Mantovani, A .; Cassatella, MA; Costantini, C .; Jaillon, S. Neutrophils nell’attivazione e regolazione dell’immunità innata e adattiva. Nat. Rev. Immunol. 2011, 11, 519-531.
125. Granot, Z. Neutrofili come bersaglio terapeutico. nel cancro. Davanti. Immunol. 2019, 10, 1710.
126. Sagiv, JY; Michaeli, J .; Assi, S .; Mishalian, I .; Kisos, H .; Levy, L .; Damti, P .; Lumbroso, D .; Polyansky, L .; Sionov, RV; et al. Diversità fenotipica e plasticità nelle sottopopolazioni circolanti di neutrofili nel cancro. Cell Rep.2015, 10, 562-573.
127. Wculek, SK; Malanchi, I. I neutrofili supportano la colonizzazione polmonare delle cellule del cancro al seno che iniziano la metastasi. Natura 2015, 528, 413–417.
128. Scapini, P .; Morini, M .; Tecchio, C .; Minghelli, S .; Di Carlo, E .; Tanghetti, E .; Albini, A .; Lowell, C .; Berton, G .; Noonan, DM; et al. L’angiogenesi indotta da CXCL1 / macrofagi indotta dalla proteina-2 in vivo è mediata dall’immunolo AJ del fattore di crescita endoteliale vascolare derivato dai neutrofili. 2004, 172, 5034-5040.
129. Nozawa, H .; Chiu, C .; Hanahan, D. I neutrofili infiltranti mediano l’interruttore angiogenico iniziale in un modello murino di carcinogenesi a più stadi. Proc. Natl. Acad. Sci. Stati Uniti d’America 2006, 103, 12493-12498.
130. Schmielau, J .; Finn, granulociti attivati da OJ e perossido di idrogeno derivato dai granulociti sono il meccanismo alla base della soppressione della funzione delle cellule T nei pazienti con carcinoma avanzato. Cancer Res. 2001, 61, 4756–4760.
131. Nicolas-Avila, JA; Adrover, JM; Hidalgo, A. Neutrophils in Omeostasi, Immunità e Cancro. Immunità 2017, 46, 15–28.
132. Ashtekar, AR; Saha, motivo di B. Poly: iscrizione al club degli APC. Trends Immunol. 2003, 24, 485–490.
133. Fridlender, ZG; Sun, J .; Kim, S .; Kapoor, V .; Cheng, G .; Ling, L .; Worthen, GS; Albelda, SM Polarizzazione del fenotipo neutrofilo associato al tumore mediante TGF-beta: “N1” contro “N2” TAN. Cellula tumorale 2009, 16, 183–194.
134. Eruslanov, EB; Bhojnagarwala, PS; Quatromoni, JG; Stephen, TL; Ranganathan, A .; Deshpande, C .; Akimova, T .; Vachani, A .; Litzky, L .; Hancock, WW; et al. I neutrofili associati al tumore stimolano le risposte delle cellule T nel carcinoma polmonare in stadio precoce. J. Clin. Investig. 2014, 124, 5466–5480.
135. Radsak, M .; Iking-Konert, C .; Stegmaier, S .; Andrassy, K .; Neutrofili polimorfonucleati di Hänsch, GM come cellule accessorie per l’attivazione delle cellule T: complesso di istocompatibilità maggiore induzione dipendente dall’antigene di proliferazione delle cellule T di classe II. Immunologia 2000, 101, 521-530.
136. Stoppacciaro, A .; Melani, C .; Parenza, M .; Mastracchio, A .; Bassi, C .; Baroni, C .; Parmiani, G .; Colombo, regressione MP di un tumore accertato geneticamente modificato per rilasciare il fattore stimolante le colonie di granulociti richiede la cooperazione tra cellule di granulociti-T e gamma di interferone prodotta dalle cellule T. J. Exp. Med. 1993, 178, 151–161.
137. Mishalian, I .; Bayuh, R .; Eruslanov, E .; Michaeli, J .; Levy, L .; Zolotarov, L .; Singhal, S .; Albelda, SM; Granot, Z .; Fridlender, ZG Neutrophils reclutano le cellule T regolatorie nei tumori attraverso la secrezione di CCL17, un nuovo meccanismo di ridotta immunità antitumorale. Int. J. Cancer 2014, 135, 1178-1186.
138. Eash, KJ; Greenbaum, AM; Gopalan, PK; Link, DC CXCR2 e CXCR4 regolano antagonisticamente il traffico di neutrofili dal midollo osseo murino. J. Clin. Investig. 2010, 120, 2423-2431.
139. Glodde, N .; Calvo, T .; van den Boorn-Konijnenberg, D .; Nakamura, K .; O’Donnell, JS; Szczepanski, S .; Brandes, M .; Eickhoff, S .; Das, I .; Shridhar, N .; et al. Reazioni reattive di neutrofili dipendenti dal recettore Tirosina chinasi c-MET Limite Immunoterapia. Immunità 2017, 47, 789–802.e9.
140. Veglia, F .; Tyurin, VA; Blasi, M .; De Leo, A .; Kossenkov, AV; Donthireddy, L .; A, TK; Schug, Z .; Basu, S .; Wang, F .; et al. La proteina di trasporto degli acidi grassi 2 riprogramma i neutrofili nel cancro. Natura 2019, 569, 73–78.
141. Brinkmann, V .; Reichard, U .; Goosmann, C .; Fauler, B .; Uhlemann, Y .; Weiss, DS; Weinrauch, Y .; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532-1535.
142. Fuchs, TA; Abed, U .; Goosmann, C .; Hurwitz, R .; Schulze, I .; Wahn, V .; Weinrauch, Y .; Brinkmann, V .; Zychlinsky, A. Un nuovo programma di morte cellulare porta a trappole extracellulari di neutrofili. J. Cell Biol. 2007, 176, 231–241.
143. Sangaletti, S.; Treppiede, C.; Chiodoni, C.; Guarnotta, C.; Cappetti, B.; Casalini, P; Piconese, S .; Parenza, M .; Guiducci, C.; Vitali, C.; et al. Trappole extracellulari di neutrofili Trasferimento mediatico di antigeni neutrofili citoplasmatici a cellule dendritiche mieloidi verso l’induzione ANCA e l’autoimmunità associata. Sangue 2012, 120, 3007–3018.
144. Papadaki, G.; Kambas, K .; Choulaki, C.; Vlachou, K .; Drakos, E.; Bertsias, G.; Ritis, K .; Boumpas, DT; Thompson, PR; Verginis, P.; et al. Trappole extracellulari di neutrofili esacerbano le risposte autoimmuni mediate da Th1 nell’artrite reumatoide promuovendo la maturazione DC. Euro. J. Immunol. 2016, 46, 2542–2554.
145. Scozzi, D .; Wang, X .; Liao, F .; Liu, Z .; Zhu, J .; Pugh, K .; Ibrahim, M .; Hsiao, H.-M .; Miller, MJ; Yizhan, G .; et al. Frammenti di trappole extracellulari neutrofili stimolano risposte immunitarie innate che impediscono la tolleranza al trapianto di polmone. Am. J. Transpl. 2019, 19, 1011-1023.
146. Tillack, K .; Breiden, P .; Martin, R .; Sospedra, Primer per linfociti M. T di trappole extracellulari di neutrofili collega risposte immunitarie innate e adattive. J. Immunol. 2012, 188, 3150–3159.
147. Rayes, RF; Mouhanna, JG; Nicolau, I .; Bourdeau, F .; Giannias, B .; Rousseau, S .; Quaglia, D .; Walsh, L .; Sangwan, V .; Bertos, N .; et al. I tumori primari inducono trappole extracellulari di neutrofili con metastasi mirabili che promuovono effetti. JCI Insight 2019, 4, e128008.
148. Flemming, A. I tumori usano le reti come scudi fisici. Nat. Rev. Immunol. 2020, 20, 352–353.
149. Teijeira, A .; Garasa, S .; Gato, M .; Alfaro, C .; Migueliz, I .; Cirella, A .; de Andrea, C .; Ochoa, MC; Otano, io .; Etxeberria, I .; et al. Gli agonisti dei recettori delle chemochine CXCR1 e CXCR2 prodotti dai tumori inducono trappole extracellulari neutrofili che interferiscono con la citotossicità immunitaria. Immunità 2020, 52, 856–871.e8.
150. Melo, RCN; Liu, L .; Xenakis, JJ; Spencer, LA Citochine derivate dagli eosinofili in salute e malattia: svelare nuovi meccanismi di secrezione selettiva. Allergy 2013, 68, 274–284.
151. Rosenberg, HF; Dyer, KD; Foster, PS Eosinophils: cambiare prospettiva in salute e malattia. Nat. Rev. Immunol. 2013, 13, 9–22.
152. Davis, BP; Rothenberg, ME Eosinofili e cancro. Cancer Immunol. Res. 2014, 2, 1–8.
153. Reichman, H .; Karo-Atar, D .; Munitz, A. Ruoli emergenti per gli eosinofili nel microambiente tumorale. Trends Cancer 2016, 2, 664–675.
154. Delyon, J .; Mateus, C .; Lefeuvre, D .; Lanoy, E .; Zitvogel, L .; Chaput, N .; Roy, S .; Eggermont, AM; Routier, E .; Robert, C. Esperienza nella pratica quotidiana con Ipilimumab per il trattamento dei pazienti con melanoma metastatico: un aumento precoce dei conteggi dei linfociti e degli eosinofili è associato a una migliore sopravvivenza. Ann. Oncol. 2013, 24, 1697–1703.
155. Gebhardt, C .; Sevko, A .; Jiang, H .; Lichtenberger, R .; Reith, M .; Tarnanidis, K .; Holland-Letz, T .; Umansky, L .; Beckhove, P .; Sucker, A .; et al. Cellule mieloidi e relativi fattori infiammatori cronici come nuovi marker predittivi nel trattamento del melanoma con Ipilimumab. Clin. Cancer Res. 2015, 21, 5453-5459.
156. McNeel, DG; Gardner, TA; Higano, CS; Kantoff, PW; Piccolo, EJ; Wener, MH; Sims, RB; DeVries, T.; Sheikh, NA; Dreicer, R. Un aumento transitorio degli eosinofili è associato alla sopravvivenza prolungata negli uomini con carcinoma della prostata resistente alla castrazione metastatica che ricevono Sipuleucel-T. Cancer Immunol. Res. 2014, 2, 988-999.
157. Tanizaki, J .; Haratani, K .; Hayashi, H .; Chiba, Y .; Nakamura, Y .; Yonesaka, K .; Kudo, K .; Kaneda, H .; Hasegawa, Y .; Tanaka, K .; et al. Biomarcatori del sangue periferico associati a risultati clinici in pazienti con carcinoma polmonare non a piccole cellule trattati con Nivolumab. J. Thorac. Oncol. 2018, 13, 97-105.
158. Lotfi, R .; Herzog, GI; DeMarco, RA; Beer-Stolz, D .; Lee, JJ; Rubartelli, A .; Schrezenmeier, H .; Lotze, MT Eosinophils Oxidize Molecole di pattern molecolari associati a danni derivati da cellule stressate. J. Immunol. 2009, 183, 5023–5031.
159. Simon, SCS; Hu, X .; Panten, J .; Grees, M .; Renders, S .; Thomas, D .; Weber, R .; Schulze, TJ; Utikal, J .; Umansky, V. L’accumulo di eosinofilo prevede la risposta al trattamento del melanoma con inibitori del checkpoint immunitario. Oncoimmunology 2020, 9, 1727116.
160. Carretero, R .; Sektioglu, IM; Garbi, N .; Salgado, OC; Beckhove, P .; Hämmerling, GJ Eosinophils Orchestrate Rifiuto del cancro normalizzando i vasi tumorali e migliorando l’infiltrazione di cellule T CD8 (+). Nat. Immunol. 2015, 16, 609–617.
161. Hollande, C .; Boussier, J .; Ziai, J .; Nozawa, T .; Bondet, V .; Phung, W .; Lu, B .; Duffy, D .; Paradis, V .; Mallet, V .; et al. L’inibizione della dipeptidil peptidasi DPP4 (CD26) rivela il controllo della crescita tumorale indotto da eosinofili dipendente dall’IL-33. Nat. Immunol. 2019, 20, 257–264.
162. Munitz, A .; Hogan, SP Eosinofili allarmanti per combattere i tumori. Nat. Immunol. 2019, 20, 250–252.
Ulteriori informazioni: Brigger, D., Riether, C., van Brummelen, R. et al. Gli eosinofili regolano l’infiammazione del tessuto adiposo e sostengono la forma fisica e immunologica nella vecchiaia. Nat Metab (2020). doi.org/10.1038/s42255-020-0228-3



{kind=link}