











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Un gruppo di ricerca incentrato sulla professoressa Yamada Hideto della Facoltà di Medicina dell’Università di Kobe e la professoressa associata Tanimura Kenji (Dipartimento di Ostetricia e Ginecologia), e la professoressa Arase Hisashi et al. dell’Istituto di ricerca per le malattie microbiche (RIMD) dell’Università di Osaka ha rivelato per la prima volta al mondo l’alta frequenza di un nuovo autoanticorpo nelle donne che soffrono di ricorrenti perdite di gravidanza.
Una ricerca congiunta dell’Università di Kobe e dell’Università di Osaka nel 2015 ha precedentemente scoperto che questo autoanticorpo causa anche malattie come la trombosi.
Si prevede che questi risultati contribuiranno all’illuminazione dei meccanismi sottostanti alla base della perdita di gravidanza ricorrente, trombosi e complicanze della gravidanza come i disturbi ipertesi; portando allo sviluppo di nuovi metodi di trattamento.
I risultati sono stati pubblicati online il 25 giugno sulla rivista ” Arthritis and Rheumatology ” dell’American College of Rheumatology .
- Quelli con perdita di gravidanza ricorrente (RPL) sperimentano aborti ripetuti e nati morti, rendendoli incapaci di dare alla luce un bambino sano.
- In Giappone, si ritiene che circa 1.400.000 donne soffrano di RPL. In più della metà dei casi la causa non può essere determinata, rendendo difficile la prescrizione di un metodo di trattamento. Questo è un grosso problema, soprattutto alla luce del basso tasso di natalità e dell’invecchiamento della popolazione giapponese.
- Questo studio si è basato presso l’Università di Kobe, che sta facendo molti sforzi nella ricerca RPL e ha coperto 227 pazienti in cinque ospedali universitari in tutto il paese. Circa un quarto di questi pazienti è risultato positivo al nuovo autoanticorpo (neo-autoanticorpo). Inoltre, quasi un quinto è risultato positivo per il neo-autoanticorpo tra i pazienti per i quali test comuni non erano stati in grado di determinare la causa.
- Si prevede che questi risultati possano illuminare i meccanismi sottostanti alla base di RPL, trombosi e disturbi ipertesi della gravidanza, portando allo sviluppo di nuovi metodi di trattamento. Inoltre, si spera che questi risultati della ricerca possano essere la chiave per risolvere il basso tasso di natalità e l’invecchiamento della popolazione.
La perdita di gravidanza ricorrente (RPL) è una patologia in cui la vittima è in grado di rimanere incinta ma sperimenta ripetuti aborti spontanei e nati morti, rendendoli incapaci di dare alla luce un bambino sano.
Si ritiene che ci siano circa 1.400.000 donne con RPL in Giappone. Dato il basso tasso di natalità e l’invecchiamento della popolazione del Giappone, questo è un problema importante da superare.
Tuttavia, attualmente la causa rimane sconosciuta in oltre la metà dei pazienti, rendendo difficile sapere come curare la RPL in molti casi.
Tuttavia, una ricerca congiunta condotta dal professor Arase Hisashi del RIMD e dal professore associato dell’Università di Kobe Tanimura Kenji ha scoperto un autoanticorpo completamente nuovo che causa la sindrome antifosfolipidica.
Questa sindrome può portare al verificarsi di trombosi (in cui i coaguli di sangue possono bloccare i vasi sanguigni negli organi vitali, mettendo in pericolo la vita come causando un ictus), aborti spontanei e disturbi ipertesi della gravidanza, che possono mettere in pericolo la vita delle donne in gravidanza.
Questi risultati sono stati pubblicati come tesi sulla rivista “Blood” dell’American Society of Hematology nel 2015.
RPL che comporta aborti ripetuti è anche una manifestazione clinica della sindrome antifosfolipidica. Come accennato in precedenza, la causa dell’RPL è sconosciuta in oltre la metà dei casi, tuttavia l’associazione tra questo neo-autoanticorpo e l’RPL non è stata studiata.
Questo gruppo di ricerca ha ipotizzato che l’anticorpo neo-auto potrebbe essere collegato a RPL in casi con cause inspiegabili.
Con questo in mente, sono stati raccolti campioni di sangue da pazienti con RPL in cinque ospedali universitari (Kobe University, University of Toyama, Okayama University, University of Tokyo e Hyogo Medical University), e i campioni sono stati testati per gli anticorpi neo-self presso RIMD .
Per la prima volta al mondo, il gruppo di ricerca ha condotto questo studio clinico per illuminare l’associazione tra RPL e il neo-autoanticorpo.
Questo studio è stato condotto dalla Kobe University, che sta facendo molti sforzi nella ricerca RPL e ha coinvolto test per gli anticorpi neo-auto con il consenso delle coppie ambulatoriali colpite da RPL in cinque ospedali universitari a livello nazionale.
Allo stesso tempo, è stato effettuato un esame dettagliato della causa della RPL in ciascun caso. Ciò includeva test per i disturbi della tiroide, un’analisi cromosomica di ogni coppia e esami del sangue per studiare le condizioni che fanno facilmente formare i coaguli di sangue, come la presenza di anticorpi antifosfolipidi.
Inoltre, sono stati condotti test sulle coppie per i complessi genetici associati alla suscettibilità a varie malattie, come l’antigene dei leucociti umani (HLA) di classe II. (Ad esempio, è noto che quelli con HLA-DR4 sono inclini a RPL).
Il metodo di test degli anticorpi neo-auto utilizzato nello studio è una tecnologia brevettata sviluppata da questo gruppo di ricerca.
Questo metodo prevede la produzione di cellule in cui sulla cellula sono presenti i complessi prodotti dalla beta2-glicoproteina I (una proteina che si ritiene colpisca l’anticorpo che causa la sindrome antifosfolipidica) e l’HLA Classe II (il tipo HLA che aumenta la suscettibilità alla sindrome antifosfolipidica) superficie.
Viene quindi condotta una reazione tra i complessi e il sangue del paziente per rilevare gli anticorpi (neo-autoanticorpi), che si legano ai complessi sulla superficie cellulare.
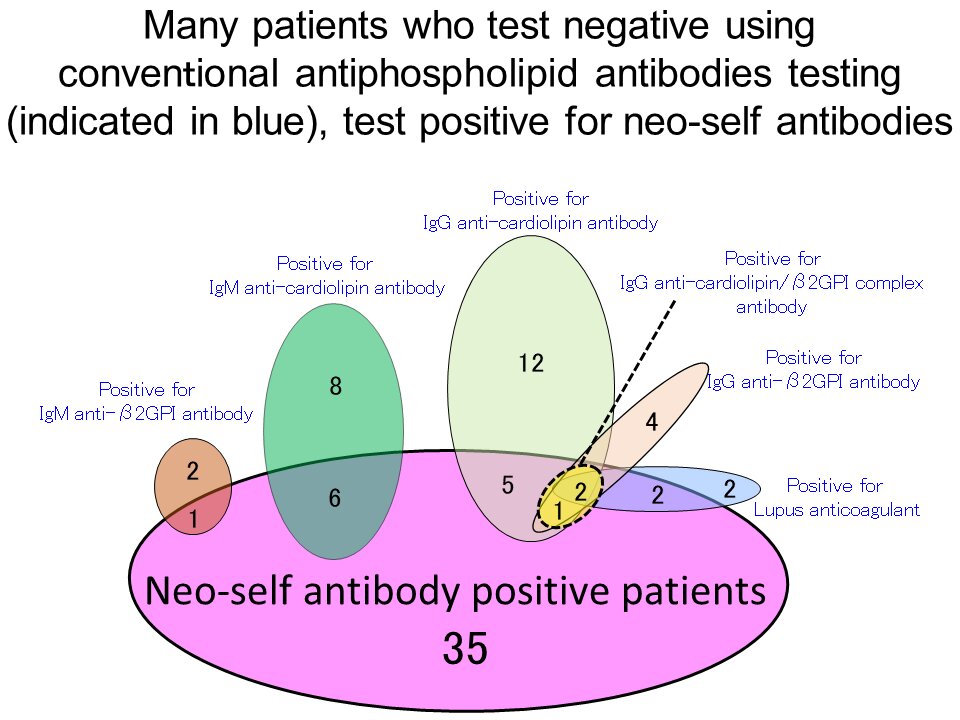
Prima di tutto, 208 donne che non avevano RPL e avevano precedentemente partorito bambini sani sono state testate per i neo-autoanticorpi per stabilire livelli normali.
Successivamente, è stato riscontrato che 52 (23%) su 227 pazienti con RPL sono risultati positivi per il neo-autoanticorpo.
Sono stati valutati i risultati dei test effettuati per determinare le cause di RPL. L’anticorpo neo-auto era il fattore con la più alta frequenza rispetto a tutti gli altri fattori che sono stati testati, inclusi problemi uterini (come malformazione e miomi), disfunzione tiroidea e anomalie cromosomiche in ciascun partner.
Ciò indica la possibilità che questo neo-autoanticorpo sia una delle principali cause di RPL.
Anche dopo aver testato le cause più comuni di RPL, la causa non è stata determinata in circa la metà dei pazienti (121 donne).
Tuttavia, 24 (20%) delle 121 donne con RPL inspiegabile sono risultate positive al solo anticorpo neo-auto (Figura 1).
In particolare, molti di coloro che sono risultati negativi per i criteri anticorpali antifosfolipidi sono risultati positivi per il neo-autoanticorpo (Figura 2).
Quando le donne positive agli anticorpi neo-auto-positivi con RPL sono state confrontate con quelle pazienti RPL che erano risultate negative, si è riscontrato che la frequenza di coloro che avevano il gene HLA-DR4, che predispone il paziente alla RPL, era più alta nei positivi dell’anticorpo neo-auto donne rispetto alle donne negative.
Attualmente, non è chiaro il motivo per cui quelli con HLA-DR4 sono inclini a RPL. Si spera che le scoperte di questo studio possano servire come chiave per comprenderne la ragione.
Studiando l’anticorpo neo-auto, i ricercatori sono stati in grado di illuminare il meccanismo di insorgenza di RPL, in particolare il meccanismo di insorgenza del disturbo in alcuni casi inspiegabili. Ciò potrebbe portare a una soluzione per i bassi tassi di natalità e l’invecchiamento della popolazione in Giappone.
Questo studio ha dimostrato che il romanzo autoanticorpo (neo-autoanticorpo) che è stato scoperto dal professore associato Tanimura e dal professor Arase et al. è una delle principali cause di RPL.
Successivamente, i ricercatori mirano a generare un farmaco che inibisce i neo-autoanticorpi o ne sopprime la produzione. Inoltre, si spera che la ricerca sugli anticorpi neo-auto illuminerà il meccanismo di insorgenza per i pazienti ostetrici con condizioni, come inspiegabili disturbi ipertesi della gravidanza e limitazione della crescita fetale, dove la causa è sconosciuta.
Ciò potrebbe portare allo sviluppo di metodi di trattamento per questi pazienti. Inoltre, è possibile che esista un anticorpo neo-auto che innesca molti disturbi autoimmuni comuni, come i reumatismi. Una simile scoperta potrebbe essere rivoluzionaria sia per la reumatologia che per l’immunologia.
La perdita di gravidanza ricorrente (RPL) è la perdita di due o più gravidanze prima delle 24 settimane di gestazione, secondo la Società europea di riproduzione umana e embriologia [1]. Il Comitato di pratica dell’American Society for Reproductive Medicine definisce RPL come due o più gravidanze cliniche fallite [2].
RPL è una sfida per la comunità clinica e scientifica. In effetti, solo nel 50% circa dei casi di RPL si possono trovare cause / fattori di rischio definiti, tra cui l’età materna avanzata, anomalie genetiche, autoanticorpi materni selezionati, disfunzioni endocrine e anomalie uterine [3]. Gli altri casi RPL attualmente non sono spiegati (uRPL) [3,4].
Si ritiene che una percentuale significativa, sebbene non esattamente quantificata, di RPL sia associata a eziologie immunitarie [5] e che in questi casi le perdite in gravidanza possano verificarsi attraverso disturbi persistenti in diverse vie immunitarie [6]. In questo contesto, l’endometrio potrebbe svolgere un ruolo rilevante.
L’endometrio ha un ruolo cruciale nella riproduzione. È il tessuto materno che entra in contatto diretto con l’embrione e consente il suo corretto impianto, sopravvivenza e sviluppo, i processi a cui partecipa attivamente.
Negli ultimi decenni sono state condotte approfondite ricerche per chiarire i meccanismi biomolecolari che rendono l’endometrio recettivo all’embrione, nonché i tipi cellulari specifici e le vie cellulari coinvolte nella ricettività endometriale. Tuttavia, la nostra conoscenza di questi meccanismi è ancora in gran parte incompleta.
Le informazioni attualmente disponibili indicano che l’endometrio è un tessuto unico in cui si verificano una serie di eventi – collettivamente chiamati decidualizzazione – al fine di garantire, in modo dinamico, l’ambiente corretto per il concetto in via di sviluppo.
Prove emergenti indicano che un ruolo chiave nel rimodellamento dell’endometrio e nella tolleranza materna nei confronti dell’embrione è svolto da diverse cellule del sistema immunitario innato e adattivo, che hanno o acquisiscono caratteristiche specifiche quando si trovano o raggiungono l’endometrio, insieme a un numero crescente di molecole immunoregolatorie.
Le evidenze sperimentali e cliniche suggeriscono anche che gli squilibri che si verificano nell’ambiente immunitario endometriale possono essere coinvolti in diverse importanti disfunzioni riproduttive, come l’insufficienza di impianto ricorrente (RIF) e la perdita di gravidanza ricorrente (RPL) di eziologia altrimenti inspiegabile.
Questa recensione riassume i principali contributi del sistema immunitario nella fisiologia della funzione endometriale e le attuali conoscenze sulle principali alterazioni conosciute del sistema immunitario endometriale con specifica applicazione all’RPL nelle donne.
Funzione endometriale fisiologica e sistema immunitario
Rimodellamento e decidualizzazione
dell’endometrio L’endometrio è il tessuto materno che entra in contatto diretto con l’embrione, che è immunologicamente diverso dalla madre, di solito considerato come un semi-allotrapianto o anche come un completo allotrapianto nel caso di donazione di ovociti [7].
La funzione principale dell’endometrio è quella di creare e garantire un ambiente endocrino / paracrino, immunitario e molecolare ottimale che consenta la corretta apposizione, attaccamento, impianto, invasione e sviluppo e la piena maturazione dell’embrione.
Per compiere questa missione, l’endometrio deve sviluppare una serie di cambiamenti adattativi unici e sorprendenti definiti collettivamente decidualizzazione, con conseguente riprogrammazione morfologica e funzionale profonda delle cellule stromali endometriali che si differenziano in cellule altamente specializzate con capacità secretorie.
Tra i mammiferi eutheriani – placentare – questi cambiamenti sono particolarmente significativi nelle specie con un tipo invasivo di placenta, in particolare negli esseri umani che hanno il tipo più invasivo di placenta.
In effetti, l’estensione della decidualizzazione è proporzionale al grado di invasività dell’embrione [8]. Mentre la trasformazione endometriale nella maggior parte delle specie animali avviene in risposta alla presenza dell’embrione, in alcune specie – compresi gli esseri umani – ha luogo con largo anticipo rispetto alla presenza dell’embrione e, quindi, è – almeno inizialmente – sotto esclusiva materna controllo ormonale.
Se non si verifica il concepimento, in risposta al calo dei livelli circolanti di progesterone, l’endometrio decidualizzato in queste specie viene eliminato e ne conseguono le mestruazioni. Questo avvia l’inizio di un nuovo ciclo.
La maggior parte delle informazioni recenti sul ruolo dell’endometrio e sui cambiamenti funzionali che si verificano nell’endometrio normale deriva dalle indagini istologiche condotte su biopsie endometriali umane, nonché da studi condotti in vivo su specie animali (in particolare babbuini e knockout genico topi) e in vitro in colture cellulari primarie di cellule stromali endometriali umane (HESC).
Nonostante le notevoli differenze tra le specie, il quadro generale emergente è che la decidualizzazione dell’endometrio è un processo che comporta una riprogrammazione cellulare profonda, rimodellamento dei tessuti, cambiamenti nell’espressione genica e regolazione post-traduzione e alterazioni nei percorsi di segnalazione cellulare.
Si verificano anche notevoli cambiamenti e modulazione dell’attività delle cellule immunitarie a livello locale. Questi eventi coinvolgono un gran numero di mediatori ed effettori molecolari. Un riepilogo dei principali cambiamenti endometriali caratteristici della decidualizzazione nelle donne è descritto nella Tabella 1.
Tabella 1
Riepilogo semplificato delle principali modifiche che caratterizzano la decidualizzazione dell’endometrio.
| Cambiamenti morfologici (tissutali e cellulari) |
| Trasformazione secretoria delle ghiandole endometriali, compattazione delle cellule epiteliali superficiali, edema stromale, proliferazione delle cellule stromali, differenziazione delle cellule stromali simili ai fibroblasti in cellule epiteliali, massicce infiltrazioni di leucociti (principalmente cellule killer uterine naturali e mastociti), rimodellamento delle arterie a spirale |
| Modifiche alla matrice extracellulare |
| Aumento della produzione da parte delle cellule stromali decidue di collagene IV, fibronectina, laminina, decorina, proteoglicani eparan-solfato, IGFBP-1, PRL, LEFTY-2, integrina αvβ3, osteopontin |
| Cambiamenti nell’espressione dei geni coinvolti nelle seguenti funzioni cellulari |
| Regolazione del ciclo cellulare, rimodellamento del citoscheletro, risposta allo stress ossidativo, trasporto di ioni e acqua, risposta all’ormone steroideo, deposizione di matrice extracellulare (ECM), modulazione della trascrizione, pattern epigenetico, modifica post-traduzione, fattore di crescita, angiogenesi, citochine e segnalazione di chemiochine , modulazione immunitaria |
| Segnalazione cellulare e percorsi coinvolti nella decidualizzazione |
| via cAMP / PKA, via di segnalazione progesterone, via nodale, via di segnalazione Notch, via ERK1 / 2, via WNT / β-catenina, via cSRC, via JAK-STAT, via lipidica (sistema endocannabinoide), via TGFβ, BMP2-WNT4 cascata di segnalazione, fosfatidilinositolo 3-chinasi / via AKT, via Ras / Raf1 / MAPK, segnalazione EPAC1 ed EPAC2 |
LEFTY-2: fattore di determinazione sinistra-destra 2; IGFBP-1: insulina come fattore di crescita che lega la proteina-1; PRL: prolattina; CAMPIONE: adenosina monofosfato ciclico; PKA: protein chinasi A; ERK1 / 2: chinasi 1/2 regolata dal segnale extracellulare; JAK: Janus chinase; STAT: trasduttore di segnale e attivatore di trascrizione; TGFβ: trasformazione del fattore di crescita-β; BMP-2: proteina morfogenetica ossea 2; AKT: protein chinasi B; MAPK: protein chinasi attivata dal mitogeno; EPAC1 e 2: scambiano proteine attivate direttamente dai cAMP 1 e 2.
La descrizione dettagliata di tutti i cambiamenti e dei mediatori coinvolti nella decidualizzazione va oltre lo scopo di questo articolo ed è stata discussa di recente in diverse eccellenti recensioni [9,10,11,12,13]. Qui, l’obiettivo principale è quello di valutare il significato dei cambiamenti endometriali, in particolare la decidualizzazione, in relazione al ruolo svolto dal sistema immunitario.
La decidualizzazione può essere evolutivamente considerata avere due funzioni, non reciprocamente esclusive. Da un lato, può essere considerata la soluzione biologica di un conflitto materno-fetale in cui l’invasività dell’embrione, volta a garantire al massimo l’espansione dei geni fetali (parzialmente diversi da quelli materni), è contrastata dalla necessità di geni materni per garantire la propria espansione nelle gravidanze presenti e future [14].
D’altro canto, può essere visto come la risposta adattativa alla necessità materna di controllare il suo investimento rilevante in una gravidanza che porta un singolo feto di alta qualità per un periodo di tempo prolungato [15].
Questo concetto è supportato da prove solide che indicano che l’endometrio decidualizzato agisce come biosensore della qualità dell’embrione [16,17], che l’impianto è caratterizzato da una cooperazione tra l’embrione e l’endometrio decidualizzato [18] e che la perdita di plasticità endometriale può essere implicato in RPL inspiegabile [19].
Molti eventi chiave che si verificano nell’endometrio durante l’impianto di decidualizzazione e in decidua durante la gravidanza sono realizzati con il contributo rilevante delle cellule immunitarie e sono mediati da molte molecole immunoregolatorie.
Prove emergenti suggeriscono che possono verificarsi alterazioni della normale funzione immunitaria in questi tessuti in sottogruppi di donne con RPL.
Cellule immunitarie e loro funzioni
nell’endometrio normale e in Decidua È stato svolto un grande lavoro sperimentale per studiare le popolazioni specifiche di cellule immunitarie nell’endometrio durante il ciclo mestruale, durante l’impianto e all’inizio della gravidanza; i loro ruoli specifici nelle prime fasi della gravidanza insieme alle molecole regolatorie che si esprimono vengono caratterizzati.
Queste informazioni sono riassunte nella Tabella 2 e Tabella 3. Allo stesso modo, viene proposto un numero crescente di meccanismi per spiegare lo sviluppo della tolleranza materna nei confronti del concetto immunologicamente diverso.
Questi meccanismi proposti – riassunti schematicamente nella Tabella 4 e illustrati nella Figura 1a, b – potrebbero essere la base per spiegare, comprendere e, possibilmente, trattare la RPL immuno-mediata, che probabilmente rappresentano una parte sostanziale di tutta la RPL inspiegabile. Tuttavia, in realtà è estremamente difficile districare i meccanismi più importanti da quelli accessori.
Il quadro generale emergente suggerisce fortemente che nella normale gravidanza, il sistema immunitario materno subisce una modulazione considerevole in almeno molti (se non tutti) componenti chiave al fine di sviluppare tolleranza verso gli antigeni paterni estranei del feto e un ambiente immunologicamente favorevole per il feto mantenendo allo stesso tempo la piena capacità di risposta contro altri antigeni stranieri [20,21,22,23].
I cambiamenti correlati alla gravidanza nelle cellule immunitarie e nel sistema che si verificano nell’endometrio e nella decidua sono determinati solo parzialmente. Tuttavia, per quanto riguarda il successo dell’impianto, i cambiamenti più importanti finora noti nelle cellule immunitarie endometriali coinvolgono macrofagi, cellule killer uterine naturali (NK), cellule dendritiche (DC), cellule T, in particolare cellule T citolitiche (CTL), i cui ruoli specifici sono stati, almeno in parte, determinati.

Rappresentazione schematica dei cambiamenti che si verificano nell’endometrio umano e nel traffico locale di cellule immunitarie nello stato normale e nella perdita di gravidanza ricorrente (RPL). a) endometrio nella fase secretoria del ciclo mestruale in assenza dell’embrione; (b) endometrio in presenza di un embrione normalmente impiantato; (c) disordini immunitari endometriali in RPL. M: macrofagi; Regno Unito: cellula killer naturale uterina; iDC: cellula dendritica uterina immatura; mDC: cellula dendritica uterina matura; N: granulociti neutrofili; Treg: cellula T regolatoria; MC: mastocita; Teff: cellule T effettrici; ESC: cellula stromale endometriale; BV: vaso sanguigno.
Tavolo 2
Panoramica delle cellule rilevanti del sistema immunitario innato nell’endometrio e nella decidua con le loro principali funzioni riproduttive.
| Tipo di cella | fenotipo | Densità (%) della popolazione di leucociti nell’endometrio | Cambiamenti durante il ciclo mestruale e all’inizio della gravidanza | Posizione preferenziale nell’endometrio | Molecole rilevanti secrete | Funzioni suggerite | Riferimenti |
|---|---|---|---|---|---|---|---|
| Cellule killer naturali uterine (Regno Unito) | CD3 – CD 56bright CD16 – (fenotipo predominante) | 30% -40% delle cellule stromali 70% dei leucociti endometriali nella fase luteale tardiva (LP) e all’inizio della gravidanza | Aumentare progressivamente dalla fase follicolare (FP) all’LP. Massima densità nel tardo LP e in decidua gestazionale | Circonda le arterie e le ghiandole | IFN-γ, VEGF, PlGF, TGF-β, TNF-α, IL-10, GM-CSF, IL-1β, LIF, CSF-1, AP-2 | Rimodellamento dei tessuti (arterie a spirale), potenziamento dell’angiogenesi, controllo dell’invasione dei trofoblasti | [10,20,24,25,26,27,28,29] |
| Macrofagi (Mφ) | CD68 + | 20% -25% dei leucociti totali nella decidua | Aumentare progressivamente dal PQ al LP. Massima densità prima delle mestruazioni e in gravidanza. Acquisire fenotipo tollerogenico | Sparsi in tutto l’endometrio; preferibilmente trovato intorno alle ghiandole e nel sito di impianto | TGF-β, IL-10, IDO, PGE2 | Coinvolto nel mantenimento del corpo luteo, impianto di blastocisti, rimodellamento delle arterie a spirale, controllo dell’invasione del trofoblasto, protezione del feto contro l’infezione intrauterina | [20,29,30,31,32,33,34,35] |
| Mast Cells (MCs) | MC T ; MC TC ; MC C (MC endometriale) | 3% -5% delle cellule endometriali totali; | Invariato durante il ciclo mestruale; cambiamenti nel fenotipo durante il ciclo mestruale; attivato nella fase iniziale e mediana | Più prominente nel compartimento endometriale basale | VEGF | Avvio delle mestruazioni Migliora il rimodellamento dell’arteria a spirale e dei tessuti, supporta l’impianto e l’angiogenesi | [20,36,37] |
Cellule dendritiche uterine (DC) | CD1a + (DC immaturi e tollerogenici); CD83 + (DC maturi) | La densità delle DC immature nell’endometrio è superiore a quella delle DC mature; DC 1% –2% delle cellule immunitarie nella decidua | DC immature aumentano da FP a LP; picco nella fase mestruale (accertamento controverso); Nessun cambiamento nelle DC mature con il ciclo mestruale | Sia le DC mature che quelle immature si trovano principalmente nello strato basale dell’endometrio nell’LP; Sparsi attraverso la decidua gestazionale Nei topi raggruppati in strutture a grappolo | TGF-β, IL-10, IDO | Coinvolto nell’accettazione materna dell’embrione, invasione e differenziazione dei trofoblasti; Rimodellamento uterino, angiogenesi; Determinare la differenziazione dei progenitori delle cellule T in Treg e l’espansione e l’attivazione di Treg | [29,33,38,39,40,41,42,43] |
| Granulociti neutrofili (N) | CD11b +; CD16b +; CD66c + | Dall’1% al 6% –15% delle cellule endometriali | Aumento considerevole del ritardo (premestruale) LP | Stroma endometriale | CCL2, CXCL8, TNF-α, IL-6, VEGF | Mestruazioni, rottura e riparazione dei tessuti, proangiogeniche e tollerogeniche nelle decidua in gravidanza |
Tabella 3
Panoramica delle cellule del sistema immunitario adattativo nell’endometrio e nella decidua con le loro principali funzioni riproduttive.
| Tipo di cella | fenotipo | Densità (%) della popolazione di leucociti (CD45 +) nell’endometrio | Cambiamenti durante il ciclo mestruale e all’inizio della gravidanza | Posizione preferenziale nell’endometrio | Molecole rilevanti secrete | Funzioni suggerite | Riferimenti |
|---|---|---|---|---|---|---|---|
| BLymphocytes | CD45 + CD19 + | 0.2%–4.5% | Lievissimo aumento nel tardo LP | Cluster tra le cellule stromali nel periodo perimensionale | IL-10 | Ancora indeterminato; Potenzialmente implicato nelle prime fasi della gravidanza | [20,47,48] |
| Linfociti T. | CD45 + CD3 + | 1% –2% al 28% | Secondo quanto riferito ridotto o invariato dalla fase follicolare (FP) alla fase luteale (LP) | Aggregati linfoidi; sparsi in tutto l’epitelio e lo stroma | Variabile in base al sottoinsieme di celle specifico | Protettivo o dannoso per l’embrione in base al sottoinsieme di cellule specifico | [24,29,48,49] |
| Linfociti T Helper (Th1, Th2, Th17, Tregs) | CD45 + CD3 + CD4 + | 3.8%–21.4% | Nessuna chiara variazione segnalata | Presente nella mucosa uterina come aggregati unici che circondano un nucleo di cellule B. | TNF-α e IFN-γ (di Th1); IL-4 (di Th2); IL-8 (di Th17) | Th1 produce citochine infiammatorie; Th2 produce citochine antinfiammatorie; Th17 ha effetti pro-infiammatori (per Treg vedere la sezione dedicata nella tabella) | [33,48,50] |
| Linfociti citotossici | CD45 + CD3 + CD8 + | 4,4% –34,5% al 66% | Riduzione significativa da FP a LP | Aggregati linfoidi | Rilasciare sostanze citotossiche (granzimi, perforina) | Potenzialmente dannoso per l’embrione. Bloccato in caso di gravidanza riuscita | [47,48,49,51] |
| Tregs | CD4 + CD25 + FOXP3 + | Non chiaramente definito nell’uomo | Espansione dell’endometrio preimpianto; Aumento della decidua nel sito di impianto e all’inizio della gravidanza fino alla media | Dati insufficienti nell’endometrio umano preimpianto | Galectin-1, TGF-β, IL-10, HO-1 | Indispensabile nel controllo di un’eccessiva risposta infiammatoria materna nel sito di impianto; Coinvolto nella tolleranza immunitaria materna all’innesto fetale, in particolare all’inizio della gravidanza; Blocco delle cellule T effettrici materne Coinvolto nella regolazione del rimodellamento vascolare materno | [20,33,49,50,52,53,54,55,56,57] |
CD: cluster di differenziazione; FP: fase follicolare; LP: fase luteale; IFN-γ: gamma di interferone; VEGF: fattore di crescita endoteliale vascolare; PlGF: fattore di crescita placentare; TNF-α: fattore di necrosi tumorale alfa; IL-10: interleuchina-10; GM-CSF: fattore stimolante le colonie di macrofagi dei granulociti; IL-1β: interleuchina-1β; LIF: fattore inibitorio della leucemia; CSF-1: fattore 1 stimolante le colonie; AP-2: proteina 2 dell’adattatore endocitico; IDO: indoleamina 2,3-diossigenasi; PGE2: prostaglandina E2; CCL2: ligando di motivo CC chemiochine 2; CXCL8: ligando motivo CXC 8; IL-6: interleuchina-6; IL-4: interleuchina-4; IL-8: interleuchina-8; HO-1: eme ossigenasi-1.
Tabella 4
Principali meccanismi proposti alla base della tolleranza immunitaria materna nei confronti dell’embrione in gravidanza normale nell’endometrio e nella decidua.
| Meccanismo | Effetto | Riferimenti |
|---|---|---|
| Aumento della secrezione di LIF e IL-1β da parte dei macrofagi uterini | Valorizzazione dell’attaccamento embrionale all’epitelio endometriale | [58] |
| Produzione di TGFβ da macrofagi decidui materni | Soppressione del rigetto EVT mediata dalle cellule NK | [59] |
| Produzione di macrofagi di TGFβ, IL-10, IDO e PGE2 | Azioni immunoinibitive e favorevoli alla tolleranza | [33] |
| Caratteristiche specifiche delle cellule del Regno Unito nell’endometrio e del DNA in gravidanza | Acquisizione di un ruolo regolatore piuttosto che la classica capacità di uccidere le cellule | [21,27,60] |
| Le cellule dNK smorzano le cellule Th17 attraverso la produzione di IFN | Sopprimere la risposta infiammatoria indotta da Th17 | [61] |
| Produzione di galectina-1 da parte di cellule dNK | Induce l’apoptosi delle cellule T CD8 + attivate | [62,63,64] |
| Reclutamento endometriale di cellule immunitarie innate (Mφ, DC e granulociti) innescate dal liquido seminale prima dell’impianto | Attivazione ed espansione di Tregs che, a sua volta, crea un microambiente uterino favorevole per l’impianto dell’embrione e migliora la tolleranza materna verso gli antigeni MHC paterni | [21,54,65,66,67] |
| Induzione ed espansione del fenotipo di DC tollerogenici | Coinvolto nel controllo e nell’attivazione di Tregs | [33,54,68,69] |
| Produzione ridotta di IL-12 da parte dei DC | Priming di cellule decidue CD4 + in un fenotipo Th2 | [70,71] |
| Le cellule EVT esprimono HLA-C, HLA-E e HLA-G ma non HLA-A e HLA-B Mancanza di espressione delle molecole di MHC da parte dei sinciziotrofoblasti | Inibizione dell’attività citolitica delle cellule dNK contro i trofoblasti Induzione dell’apoptosi potenziata dalla sensibilità NK delle cellule CD8 + attivate | [70,72,73,74,75] |
| Produzione di trofoblasti di esosomi | Sottoregolazione dell’immunità materna nei confronti del trofoblasto | [76] |
| Sviluppo di una specifica rete di citochine e chemochine nell’endometrio e nella decidua | Raggiungimento di un corretto reclutamento e dialogo delle cellule immunitarie a favore dell’impianto dell’embrione e della corretta evoluzione della gravidanza | [77] |
| Presenza di addominali asimmetrici materni contro antigeni paterni | Lega il trofoblasto ma non è in grado di innescare una risposta immunitaria distruttiva | [78] |
| Aumento della produzione di PIBF | Upregulation della produzione di citochine Th2. Downregulation dell’attività di dNK. Aumentare la produzione di glicodelina A che induce l’apoptosi nelle cellule T. Questi effetti possono contribuire alla protezione immunitaria del trofoblasto | [66,79] |
| Aumento della produzione di progesterone in LP e in gravidanza | Espansione delle Treg e valorizzazione delle loro azioni immunosoppressive | [33,78] |
| produzione di hCG da parte di syncytiotrophoblast | Reclutamento di Tregs all’interfaccia materno-fetale | [80] |
EVT: trofoblasto stravagante; HLA: antigene leucocitario umano; MHC: grande complesso di istocompatibilità; Mφ: macrofagi; DC: cellule dendritiche; PIBF: fattore di blocco indotto dal progesterone; LP: fase luteale; hCG: gonadotropina corionica umana; Abs: anticorpi asimmetrici.
riferimento: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6862690/
Ulteriori informazioni: Kenji Tanimura et al., Il complesso β2-glicoproteina I / HLA ‐ DR è il principale bersaglio autoanticorpale nella sindrome antifosfolipidica ostetrica, artrite e reumatologia (2020). DOI: 10.1002 / art. 41410



{kind=link}