











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



species of mosquito originally from Asia threatens to put tens of millions of city-dwellers in Africa at higher risk of catching malaria as the invading insect spreads throughout the continent, a study said Monday.
Malaria – which killed 400,000 people in 2018, mainly children in Africa – is caused by parasites that roughly 40 mosquito species spread among humans when they feed.
The Anopheles gambiae group of mosquito species are the main drivers of malaria’s spread in Africa, but these insects dislike the polluted puddles seen in cities and haven’t learned to lay their larvae in urban fresh water tanks.
For these reasons, most malaria transmission in Africa occurs in rural areas.
In a new study published in the Proceedings on the National Academy of Sciences (PNAS), medical entomologist Marianne Sinka, of the University of Oxford, charted the spread of another species, Anopheles stephensi, which originated in Asia.
This species has learned to slip through cracks to access water tanks, favoring those made from brick and cement.
“It’s the only one that’s really good at getting into central urban areas,” .
Anopheles stephensi caused a major outbreak in Djibouti City in the Horn of Africa in 2012, a city where malaria hardly existed, and has since been observed in Ethiopia, Sudan and elsewhere.
Sinka and colleagues combined location data for the species with spatial models that identified the environmental conditions characterizing its preferred habitat: high-density urban areas where it is hot and rainfall is plentiful.
Their study found that 44 cities are “highly suitable” locations for the insect, putting 126 million more Africans – mainly around the equatorial regions – at risk of malaria, compared to today.
“That means that Africa, which has already got the highest burden of malaria, could have an even bigger impact,” said Sinka, with 40 percent of the continent’s population in urban areas.
Unlike African mosquitoes, which like to bite humans when at night when it is cool, Anopheles stephensi can feed in the evening when it is warmer, making bed nets less effective.
So installing mosquito nets on windows, soaking the walls in insecticides, and covering the body are better ways to protect against this species.
Longer term, the most effective measure is to target the larvae: eliminate stagnant water and tightly seal water tanks from intrusion. These methods proved effective in India, said Sinka.
Malaria remains a leading global health concern with over 250 million cases reported yearly [1]. In Ethiopia, even though there has been steady progress in the reduction of malaria [2], 1.5 million cases were reported in 2018 [1]. Developing effective malaria control strategies in Ethiopia require knowledge of local mosquito vector species [3].
One threat to continued progress against malaria is the expansion of vectors into new areas. The South Asian vector Anopheles stephensi was recently discovered in Ethiopia and is raising concerns about the impact on malaria transmission in the country and the rest of the Horn of Africa.
Anopheles stephensi is a major malaria vector in South Asia and the Middle East, including the Arabian Peninsula [4], and is known to transmit both the major malaria parasite species Plasmodium falciparum and P. vivax [5, 6].
The first report of An. stephensi in the Horn of Africa was from Djibouti in 2013 [7] and was recently confirmed to be persisting in the country [8]. Anopheles stephensi was detected in Ethiopia for the first time in 2016 in Kebridehar (Somali Region) but it remains unclear how broadly distributed the species is in the rest of the country [9].
Understanding the distribution of An. stephensi in Ethiopia is critical to evaluating the threat it poses to malaria control in Ethiopia and the rest of the Horn of Africa [9].
It is important during initial surveillance of a potential new vector to evaluate the accuracy of species identifications. Genetic analysis can be a useful complement to morphological identification to achieve optimal accuracy in species identification [10], particularly when identifying a recently detected species.
The objective of the study was to investigate the geographic distribution of An. stephensi in north eastern and eastern urban localities in Ethiopia using morphological and molecular identification of wild-caught Anopheles.
Methods
Survey sites
Anopheles stephensi surveys were conducted from August to November 2018 in ten selected urban sites situated in a climatic zone of either tropical, hot semi-arid or desert with an elevation range of 294 to 2055 meters above sea level. The localities included five in Somali region, three in Afar, one in Amhara region, and Dire Dawa city (Table 1, Fig. 1).
Table 1 Collection site altitude and geographical coordinates
From: Geographical distribution of Anopheles stephensi in eastern Ethiopia
| Site | Region | Altitude (masl) | Coordinates |
|---|---|---|---|
| Jigjiga | Somali | 1657 | 9°3′51″N, 42°7′93″E |
| Erer Gota | Somali | 1090 | 9°5′56″N, 41°3′84″E |
| Kebridehar | Somali | 532 | 6°7′38″N, 44°2′77″E |
| Godey | Somali | 294 | 5°9′49″N, 43°5’53″E |
| Degehabur | Somali | 1065 | 8°2′23″N, 43°5′58″E |
| Semera | Afar | 431 | 11°7′94″N, 41°0′08″E |
| Gewane | Afar | 617 | 10°1′66″N, 40°6′46″E |
| Awash Sebat Kilo | Afar | 916 | 8.9′89″N, 40°1′64″E |
| Dire Dawa | Dire Dawa | 1178 | 9°5′96″N, 41°8′54″E |
| Bati | Amhara | 2055 | 11°1′92″N, 40°0′17″E |
- Abbreviation: masl, meters above sea level
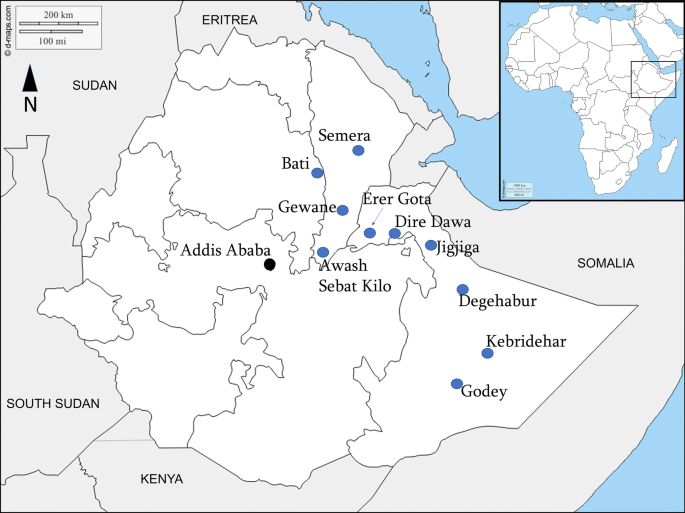
The areas have mean annual temperatures of about 20 °C to 30 °C and a mean annual rainfall from 200 to 900 mm. There is a smaller rainy season between March and May, followed by a longer period between July and October [11].
Results
A total of 82 adult An. stephensi from 300 PSCs and 200 CDC light traps were collected in 7 of the 10 sites. The sites with no adult collections were Jigjiga, Awash Sebat Kilo and Bati (Table 1). Of the 82 adults 81.7% (n = 67) were from PSC and the remaining 18.3% (n = 15) were from CDC light traps. The majority of An. stephensi sampled using PSC were from Semera and Erer and that of CDC were from Degehabur. Larval and pupal collections yielded 2149 adult An. stephensi from all the sites confirming the presence of immature stages (Table 2).
Table 2 Number of adult An. stephensi collections from PSC, CDC traps, and larval and pupal collections
From: Geographical distribution of Anopheles stephensi in eastern Ethiopia
| Region | Site | Method of collections and An. stephensi | |||
|---|---|---|---|---|---|
| PSC | CDC | Adults raised from larvae and pupae | Total | ||
| n (%) | n (%) | n (%) | |||
| Somali | Jigjiga | 0 (0) | 0 (0) | 18 (100) | 18 |
| Erer Gota | 14 (9.8) | 0 (0) | 129 (90.2) | 143 | |
| Kebridehar | 3 (0.4) | 0 (0) | 700 (99.6) | 703 | |
| Godey | 2 (0.6) | 0 (0) | 340 (99.4) | 342 | |
| Degehabur | 1 (0.6) | 13 (7.2) | 166 (92.2) | 180 | |
| Afar | Semera | 38 (12.7) | 1 (0.3) | 260 (87.0) | 299 |
| Gewane | 6 (5.5) | 1 (0.9) | 102 (93.6) | 109 | |
| Awash Sebat Kilo | 0 (0) | 0 (0) | 26 (100) | 26 | |
| Dire Dawa | Dire Dawa | 3 (1.1) | 0 (0) | 277 (98.9) | 280 |
| Amhara | Bati | 0 (0) | 0 (0) | 135 (100) | 135 |
| Total | 67 | 15 | 2149 | 2231 |
- Abbreviation: n, number collected
Larval breeding habitats included concrete water cisterns, discarded tires, water tanks, steel drums, plastic sheet water storage at construction sites, discarded buckets, abandoned car wash and discarded vehicle part (Fig. 2). The potential breeding containers in each of the sites are presented in Table 3. However, since this was a onetime cross-sectional survey, the likelihood of missing other breeding sites is inevitable. Moreover, the containers found without larvae at the time of the survey might be positive another time because of the seasonality of the population of An. stephensi. Larvae and pupae of An. stephensi and Aedes aegypti were visually detected but not recorded. PCR endpoint assay was performed, and successful PCR products were obtained for 130 out of 133 Anopheles specimens. With the PCR endpoint assay, 119 specimens were identified as An. stephensi and 11 specimens were identified as non-An. stephensi. Sequencing of portions of the ITS2 and cox1 loci was also completed and successful sequencing was completed for 118 Anopheles specimens. BLAST analysis of Anopheles sequences confirmed the positive detection of An. stephensi at all ten sites. Sequence-based species identification was mostly consistent with ITS2 endpoint assay results, except for a single specimen that was morphologically identified as An. gambiae (s.l.), identified as non-An. stephensi with the endpoint assay, but sequencing detected An. stephensi. BLAST analysis of ITS2 sequences further identified all the non-An. stephensi specimens as An. arabiensis.

Table 3 Distribution of An. stephensi based on larval breeding habitats
From: Geographical distribution of Anopheles stephensi in eastern Ethiopia
| Region | Site | Larval breeding habitat | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Construction water storage reservoirs (n) | Discarded tires (n) | Steel drums (n) | Water tanks (n) | Temporary water storage reservoirs (n) | Birkas (n) | Abandoned car wash (n) | Discarded vehicle part (n) | Broken bucket (n) | Total (n) | ||
| Somali | Jigjiga | nf | 0 | 0 | 0 | 15 | 3 | nf | nf | nf | 18 |
| Erer | nf | 0 | nf | 5 | 100 | nf | 24 | nf | nf | 129 | |
| Kebridehar | nf | 0 | 0 | 0 | 700 | nf | nf | nf | 700 | ||
| Godey | nf | 0 | 0 | 0 | 116 | 224 | nf | nf | nf | 340 | |
| Degehabur | nf | 0 | 0 | 0 | 34 | 132 | nf | nf | nf | 166 | |
| Afar | Semera | 225 | 5 | 0 | 17 | 13 | nf | nf | nf | 260 | |
| Gewane | 12 | 16 | 39 | 11 | nf | nf | 5 | 19 | 102 | ||
| Awash Sebat Kilo | 26 | nf | nf | 0 | nf | nf | nf | nf | 26 | ||
| Dire Dawa | Dire Dawa | 100 | 0 | 24 | 53 | 100 | nf | nf | nf | nf | 277 |
| Amhara | Bati | 11 | 7 | 2 | 5 | 110 | nf | 0 | nf | nf | 135 |
- Abbreviations: nf, not found during the survey; n, number of An. stephensi
We compared the morphological identification to the ITS2 PCR endpoint assay results. Of the 130 Anopheles, 124 were classified as An. stephensi and six as An. gambiae (s.l.) based on morphology. Five of the 124 (4.0%) morphologically identified An. stephensi were not confirmed to be An. stephensi with the PCR endpoint-assay. All morphologically identified An. gambiae (s.l.) (sequence-confirmed An. arabiensis) that were successfully amplified were also identified as non-An. stephensi with the ITS2 PCR endpoint assay.
All 82 wild-caught adult An. stephensi were screened for P. falciparum or P. vivax infection. No Plasmodium DNA was detected in any of the specimens.
Discussion
This survey confirms that An. stephensi is distributed broadly in eastern Ethiopia. These data, taken with previous reports of An. stephensi in Kebridehar in 2016 [9], confirm that An. stephensi is established in this region. To our knowledge, this is the first evidence for the presence of adult An. stephensi in multiple regions in Ethiopia, where it might transmit malaria.
The widespread presence of An. stephensi in Ethiopia along with Djibouti suggests that neighboring countries, such as Sudan, South Sudan, Eritrea, Somalia and Kenya, should also enhance surveillance.
Given that An. stephensi in Djibouti was found to carry both P. falciparum and P. vivax [7, 8], there is potential for these parasites to be observed in Ethiopia; thus, malaria control strategies should now consider the potential for the established An. stephensi to transmit malaria.
The presence of An. stephensi was confirmed using both morphological and molecular methods. While morphology was mostly consistent with molecular approach (119/124 correctly identified An. stephensi), there were a few instances of incorrect identification based on morphology highlighting the risk for the misidentification of specimens.
As more vector surveillance programmes in Africa incorporate An. stephensi into their morphological keys, molecular data can be helpful with the evaluation of the successful training in An. stephensi morphological identification.
This is particularly important at this beginning phase of An. stephensi surveillance as field technicians will be adapting to detecting An. stephensi.
We incorporated two molecular approaches, one of which, the end-point assay using primers designed by Djadid et al. [13] is more feasible in resource-limited settings. We found that this assay was mostly consistent with the sequence data and has to potential be integrated into current PCR based assays that focus on the detection of members of the An. gambiae complex, the most common malaria vectors in Africa.
While we have confirmed the broad distribution of An. stephensi in eastern Ethiopia, the distribution in the western part of the country is yet to be determined. West Ethiopia has had more consistent surveillance of malaria vectors than the eastern, due to the burden of the disease there; however, previously used trapping methods may limit the ability to detect An. stephensi.
The current trapping techniques that rely heavily on CDC light traps may limit the ability to detect An. stephensi, given the low number of An. stephensi caught with CDC light traps at most sites in this study.
Additional studies on the breeding, feeding, and resting behavior of An. stephensi can provide crucial information that can be applied to enhance future surveillance efforts in west and eastern Ethiopia.
Several additional areas of query need to be pursued further to better inform vector control efforts. No study has been published to confirm that the Ethiopian An. stephensi can or does transmit Plasmodium. Both field confirmation of infected An. stephensi and laboratory infections are helpful approaches evaluating this information.
In the present study, the 82 wild-caught An. stephensi were screened for both P. falciparum and P. vivax using PCR, and Plasmodium was not detected. This is not unexpected as the regions included in this study report low malaria transmission, so a much larger sample size would be required to detect Plasmodium infection in An. stephensi.
Future surveillance will continue to screen for Plasmodium using both a PCR-based and circumsporozoite protein enzyme-linked immunosorbent assay (ELISA).
The surveillance of An. stephensi to this point has been conducted in short time spans, with limited ability to assess changes in An. stephensi population size over time.
We will be repeating collections in multiple sites in eastern Ethiopia to provide crucial information about how the population is changing year to year. This information will be particularly important as new vector control interventions are rolled out to evaluate their effectiveness.
Insecticide resistance has been reported in the dominant malaria vector An. arabiensis in Ethiopia [17], but insecticide resistance status in An. stephensi is unknown. Investigations into insecticide resistance, the molecular mechanisms behind resistance, and potential genetic markers that can be used for surveillance are ongoing.
One question that remains unanswered relates to the origin of An. stephensi in Ethiopia. Previous phylogeographic analysis revealed the closest cox1 sequence similarity of the Ethiopia An. stephensi found in Kebridehar to a specimen from Pakistan [9].
Phylogeographic analysis including sequencing from recent global An. stephensi collections using multiple loci or whole genome sequences can help identify the exact origin of the An. stephensi in the Horn of Africa and how it has spread throughout the region. This information will support efforts to prevent further introduction and spread of An. stephensi.
An ancillary observation during the surveillance was the detection of larvae of the dengue vector Ae. aegypti together with An. stephensi larvae, suggesting that these two vectors share larval habitats. Dengue is a growing public health concern in Ethiopia, particularly in eastern Ethiopia, where major outbreaks were reported in 2013 [18, 19] and 2015 [20].
With the finding of Ae. aegypti larvae with An. stephensi, we can consider integrated vector control to target both An. stephensi and Ae. aegypti. This would be a cost-effective approach to reducing both malaria and dengue virus transmission. Future surveillance in eastern Ethiopia will work towards determining relative abundance of Ae. aegypti larvae at An. stephensi breeding sites.
reference link : https://parasitesandvectors.biomedcentral.com/articles/10.1186/s13071-020-3904-y
Bionomics
An. stephensi is a common laboratory species (as a consequence of its ability to breed in small containers), yet there are little quantified data describing the bionomics of An. stephensi in the wild. No reported entomological inoculation rate (EIR—the number of infected mosquito bites received per unit time) and only a single human biting rate value (HBR—the number of mosquito bites per unit time) could be found in the literature (42).
In a brief summary of Asian species bionomics, Sinka et al. (9) indicate An. stephensi primarily prefers to feed on animals (zoophilic) with examples of human biting (anthropophagy) indoors (endophagic) with outdoor (endophilic) resting behavior, but with examples of outdoor biting (exophagic) behavior, with a preference for biting during dusk (crepuscular) and the night.
Similar to many of the Asian dominant vector species, An. stephensi appears to have some level of plasticity in its behavior, specifically in host choice, with rural An. stephensi tending to be more zoophilic than urban populations (42, 43). The mosquito’s biting behavior is driven by host availability including human activity patterns and livestock sheltering practices.
Fed female mosquitoes are reported as preferring to rest in poorly constructed buildings or animal sheds (42⇓⇓–45). Such structures are characteristic of unplanned urban sites and can be found around and within many rapidly expanding cities in Africa (46, 47).
Many malaria vector surveys include hourly mosquito catches that run between 6 PM and 6 AM, when anopheline vectors are typically most active. Using data from the Malaria Atlas Project’s bionomics database (48), nightly activity for the three primary African vector species (An. gambiae, An. arabiensis, and An. funestus) are compared with An. stephensi (Fig. 5).
This simple plot illustrates that An. stephensi has peak activity (i.e., is actively seeking a blood meal) earlier in the evening than any of the African dominant vector species (DVS) with over half of the total bites from this species occurring within the first quarter of the night (6 PM to 9 PM). Activity can be observed later, but at a much lower rate (42).
African vector species are known to prefer to bite when humans are asleep and vulnerable, and as expected, An. gambiae and An. funestus both have peak activity well into the night (third quarter: midnight to 3 AM). Whether newly establishing African An. stephensi populations maintain a preference for biting in the evening remains to be seen, and there is some indication that it may change its outdoor biting activity depending on the season (42).
However, its crepuscular biting preference would allow An. stephensi to bypass insecticide treated bednets; the most widely used (and currently most effective) mosquito interventions used across Africa (49, 50). An. stephensi larvae have been found coexisting in water storage containers with Aedes aegypti (a vector of yellow fever and dengue among others) and Culex quinquefasciatus (the southern house mosquito—vector of lymphatic filariasis and West Nile fever) so interventions already in place to target these mosquitoes, as well as better management of urban infrastructure including water and green spaces, could help control An. stephensi (51⇓–53)

An. stephensi is known to have a seasonal peak in abundance, normally in the late spring/early summer months (42, 43, 54), and shows some regulation by climatic conditions; here we have seen annual temperature is the most influential covariate in describing this species’ niche. In urban areas, An. stephensi has been found in roof gutters, domestic wells, air conditioning units, and cisterns (9, 52, 55).
It is not reliant on rainfall to fill its larval sites like many other species (e.g., An. gambiae that makes use of temporary rain-filled puddles), but takes advantage of human built and maintained water storage facilities, particularly cement or brick structures (19, 20, 42, 43, 52, 55, 56); a necessity to the human urban populations during the dry summer months.
Indeed, it is An. stephensi’s ability to utilize such human built water storage tanks that may have facilitated its ability to expand its range (15). Within our model, human population density was also a highly influential variable and our final ensemble map suggests that many large urban expanses are highly suitable if this species continues to invade Africa, whereas most rural areas are not.
reference link: https://www.pnas.org/content/early/2020/09/08/2003976117
More information: M. E. Sinka el al., “A new malaria vector in Africa: Predicting the expansion range of Anopheles stephensi and identifying the urban populations at risk,” PNAS (2020). www.pnas.org/cgi/doi/10.1073/pnas.2003976117



{kind=link}
{kind=link}