










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Le specie di zanzare originarie dell’Asia minacciano di esporre decine di milioni di abitanti delle città in Africa a un rischio maggiore di contrarre la malaria mentre l’insetto invasore si diffonde in tutto il continente, secondo uno studio.
La malaria – che ha ucciso 400.000 persone nel 2018, principalmente bambini in Africa – è causata da parassiti che circa 40 specie di zanzare si diffondono tra gli esseri umani quando si nutrono.
Il gruppo di specie di zanzare Anopheles gambiae è il principale motore della diffusione della malaria in Africa , ma questi insetti non amano le pozzanghere inquinate che si vedono nelle città e non hanno imparato a deporre le loro larve nelle cisterne urbane di acqua dolce.
Per questi motivi, la maggior parte della trasmissione della malaria in Africa avviene nelle zone rurali.
In un nuovo studio pubblicato in Proceedings on the National Academy of Sciences (PNAS), l’entomologa medica Marianne Sinka, dell’Università di Oxford, ha tracciato la diffusione di un’altra specie, Anopheles stephensi, che ha avuto origine in Asia.
Questa specie ha imparato a scivolare attraverso le fessure per accedere alle cisterne d’acqua, privilegiando quelle in mattoni e cemento.
“È l’unico veramente bravo a entrare nelle aree urbane centrali”.
Anopheles stephensi ha causato una grave epidemia nella città di Gibuti nel Corno d’Africa nel 2012, una città dove la malaria non esisteva quasi mai, e da allora è stata osservata in Etiopia, Sudan e altrove.
Sinka e colleghi hanno combinato i dati di localizzazione per la specie con modelli spaziali che hanno identificato le condizioni ambientali che caratterizzano il suo habitat preferito: aree urbane ad alta densità dove fa caldo e piove abbondanti.
Il loro studio ha rilevato che 44 città sono luoghi “altamente adatti” per l’insetto, mettendo 126 milioni di africani in più – principalmente nelle regioni equatoriali – a rischio di malaria, rispetto ad oggi.
“Ciò significa che l’Africa, che ha già il più alto carico di malaria, potrebbe avere un impatto ancora maggiore”, ha detto Sinka, con il 40% della popolazione del continente nelle aree urbane.
A differenza delle zanzare africane, a cui piace mordere gli esseri umani quando di notte quando fa freddo, Anopheles stephensi può nutrirsi la sera quando fa più caldo, rendendo le zanzariere da letto meno efficaci.
Quindi installare zanzariere alle finestre, immergere le pareti in insetticidi e coprire il corpo sono modi migliori per proteggersi da questa specie.
A lungo termine, la misura più efficace è mirare alle larve: eliminare l’acqua stagnante e sigillare ermeticamente i serbatoi dell’acqua dall’intrusione. Questi metodi si sono dimostrati efficaci in India, ha detto Sinka.
La malaria rimane una delle principali preoccupazioni sanitarie globali con oltre 250 milioni di casi segnalati ogni anno [1]. In Etiopia, anche se sono stati compiuti progressi costanti nella riduzione della malaria [2], nel 2018 sono stati segnalati 1,5 milioni di casi [1]. Lo sviluppo di strategie efficaci di controllo della malaria in Etiopia richiede la conoscenza delle specie locali di vettori di zanzare [3].
Una minaccia ai continui progressi contro la malaria è l’espansione dei vettori in nuove aree. Il vettore dell’Asia meridionale Anopheles stephensi è stato recentemente scoperto in Etiopia e sta sollevando preoccupazioni circa l’impatto sulla trasmissione della malaria nel paese e nel resto del Corno d’Africa.
Anopheles stephensi è un importante vettore della malaria nell’Asia meridionale e nel Medio Oriente, inclusa la penisola arabica [4], ed è noto per trasmettere sia le principali specie di parassiti della malaria Plasmodium falciparum che P. vivax [5, 6].
La prima relazione di An. stephensi nel Corno d’Africa proveniva da Gibuti nel 2013 [7] ed è stato recentemente confermato che persiste nel paese [8]. Anopheles stephensi è stato rilevato in Etiopia per la prima volta nel 2016 a Kebridehar (regione somala) ma non è chiaro quanto sia ampiamente distribuita la specie nel resto del paese [9].
Comprendere la distribuzione di An. stephensi in Etiopia è fondamentale per valutare la minaccia che rappresenta per il controllo della malaria in Etiopia e nel resto del Corno d’Africa [9].
È importante durante la sorveglianza iniziale di un potenziale nuovo vettore per valutare l’accuratezza delle identificazioni delle specie. L’analisi genetica può essere un utile complemento all’identificazione morfologica per ottenere un’accuratezza ottimale nell’identificazione delle specie [10], in particolare quando si identifica una specie rilevata di recente.
L’obiettivo dello studio era di indagare la distribuzione geografica di An. stephensi nelle località urbane nord-orientali e orientali in Etiopia utilizzando l’identificazione morfologica e molecolare di Anopheles catturati in natura.
Metodi
Siti di indagine
Le indagini Anopheles stephensi sono state condotte da agosto a novembre 2018 in dieci siti urbani selezionati situati in una zona climatica tropicale, calda semi-arida o desertica con un intervallo di altitudine da 294 a 2055 metri sul livello del mare. Le località includevano cinque nella regione somala, tre in Afar, una nella regione di Amhara e la città di Dire Dawa (Tabella 1, Fig. 1).
Tabella 1 Altitudine del sito di raccolta e coordinate geografiche
Da: Distribuzione geografica di Anopheles stephensi nell’Etiopia orientale
| Posto | Regione | Altitudine (mslm) | Coordinate |
|---|---|---|---|
| Jigjiga | Somalo | 1657 | 9°3′51″N, 42°7′93″E |
| Erer Drop | Somalo | 1090 | 9°5′56″N, 41°3′84″E |
| Kebridehar | Somalo | 532 | 6°7′38″N, 44°2′77″E |
| Godey | Somalo | 294 | 5°9′49″N, 43°5’53″E |
| Degehabur | Somalo | 1065 | 8°2′23″N, 43°5′58″E |
| Semera | Lontano | 431 | 11°7′94″N, 41°0′08″E |
| Gewane | Lontano | 617 | 10°1′66″N, 40°6′46″E |
| Awash Sebat Kilo | Lontano | 916 | 8.9′89″N, 40°1′64″E |
| Dire Dawa | Dire Dawa | 1178 | 9°5′96″N, 41°8′54″E |
| Bati | Amhara | 2055 | 11°1′92″N, 40°0′17″E |
- Abbreviazione : mslm, metri sul livello del mare
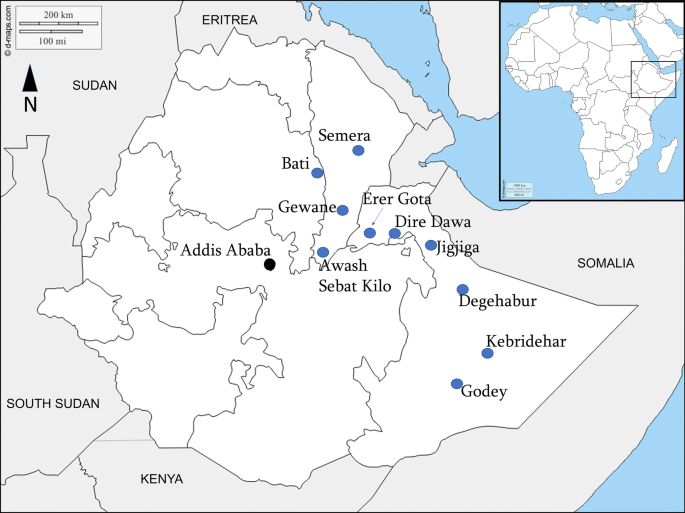
Le aree hanno temperature medie annuali di circa 20 ° C a 30 ° C e una piovosità media annua da 200 a 900 mm. C’è una stagione delle piogge più piccola tra marzo e maggio, seguita da un periodo più lungo tra luglio e ottobre [11].
Risultati
Un totale di 82 adulti An. stephensi da 300 PSC e 200 trappole di luce CDC sono stati raccolti in 7 dei 10 siti. I siti senza collezioni per adulti erano Jigjiga, Awash Sebat Kilo e Bati (Tabella 1). Degli 82 adulti, l’81,7% (n = 67) proveniva da PSC e il restante 18,3% (n = 15) proveniva da trappole luminose CDC. La maggior parte di An. Gli stephensi campionati usando PSC provenivano da Semera ed Erer e quello di CDC proveniva da Degehabur. Le raccolte larvali e pupali hanno prodotto 2149 An adulti. stephensi da tutti i siti confermando la presenza di stadi immaturi (Tabella 2).
Tabella 2 Numero di adulti An. raccolte stephensi da PSC, trappole CDC e raccolte larvali e pupali
Da: distribuzione geografica di Anopheles stephensi nell’Etiopia orientale
| Regione | Posto | Metodo di raccolta e An. stephensi | |||
|---|---|---|---|---|---|
| PSC | CDC | Adulti allevati da larve e pupe | Totale | ||
| n (%) | n (%) | n (%) | |||
| Somalo | Jigjiga | 0 (0) | 0 (0) | 18 (100) | 18 |
| Erer Drop | 14 (9.8) | 0 (0) | 129 (90.2) | 143 | |
| Kebridehar | 3 (0.4) | 0 (0) | 700 (99.6) | 703 | |
| Godey | 2 (0.6) | 0 (0) | 340 (99.4) | 342 | |
| Degehabur | 1 (0.6) | 13 (7.2) | 166 (92.2) | 180 | |
| Lontano | Semera | 38 (12.7) | 1 (0.3) | 260 (87.0) | 299 |
| Gewane | 6 (5.5) | 1 (0.9) | 102 (93.6) | 109 | |
| Awash Sebat Kilo | 0 (0) | 0 (0) | 26 (100) | 26 | |
| Dire Dawa | Dire Dawa | 3 (1.1) | 0 (0) | 277 (98.9) | 280 |
| Amhara | Bati | 0 (0) | 0 (0) | 135 (100) | 135 |
| Totale | 67 | 15 | 2149 | 2231 |
- Abbreviazione : n, numero raccolto
Gli habitat di riproduzione delle larve includevano cisterne d’acqua in cemento, pneumatici scartati, serbatoi d’acqua, fusti in acciaio, stoccaggio dell’acqua in fogli di plastica nei cantieri, secchi scartati, autolavaggi abbandonati e parti di veicoli scartate (Fig. 2). I potenziali contenitori di riproduzione in ciascuno dei siti sono presentati nella Tabella 3.
Tuttavia, poiché questa era un’indagine trasversale precedente, la probabilità di perdere altri siti di riproduzione è inevitabile. Inoltre, i contenitori trovati senza larve al momento dell’indagine potrebbero essere positivi un’altra volta a causa della stagionalità della popolazione di An. stephensi.
Larve e pupe di An. stephensi e Aedes aegypti sono stati rilevati visivamente ma non registrati. È stato eseguito il test dell’endpoint PCR e sono stati ottenuti prodotti PCR di successo per 130 dei 133 campioni di Anopheles. Con il test endpoint PCR, 119 esemplari sono stati identificati come An. stephensi e 11 esemplari sono stati identificati come non-An. stephensi.
È stato inoltre completato il sequenziamento di porzioni dei loci ITS2 e cox1 ed è stato completato con successo il sequenziamento per 118 campioni di Anopheles. L’analisi BLAST delle sequenze di Anopheles ha confermato il rilevamento positivo di An. stephensi in tutti e dieci i siti.
L’identificazione delle specie basata sulla sequenza era per lo più coerente con i risultati del test dell’endpoint ITS2, ad eccezione di un singolo campione identificato morfologicamente come An. gambiae (sl), identificato come non-An. stephensi con il test endpoint, ma il sequenziamento ha rilevato An. stephensi. L’analisi BLAST delle sequenze ITS2 ha ulteriormente identificato tutti i non-An. esemplari stephensi come An. arabiensis.
È stato completato anche il sequenziamento di porzioni dei loci ITS2 e cox1 ed è stato completato con successo il sequenziamento per 118 campioni di Anopheles. L’analisi BLAST delle sequenze di Anopheles ha confermato il rilevamento positivo di An. stephensi in tutti e dieci i siti.
L’identificazione delle specie basata sulla sequenza era per lo più coerente con i risultati del test dell’endpoint ITS2, ad eccezione di un singolo campione identificato morfologicamente come An. gambiae (sl), identificato come non-An. stephensi con il test endpoint, ma il sequenziamento ha rilevato An. stephensi. L’analisi BLAST delle sequenze ITS2 ha ulteriormente identificato tutti i non-An. esemplari stephensi come An. arabiensis.
È stato completato anche il sequenziamento di porzioni dei loci ITS2 e cox1 ed è stato completato con successo il sequenziamento per 118 campioni di Anopheles. L’analisi BLAST delle sequenze di Anopheles ha confermato il rilevamento positivo di An. stephensi in tutti e dieci i siti.
L’identificazione delle specie basata sulla sequenza era per lo più coerente con i risultati del test dell’endpoint ITS2, ad eccezione di un singolo campione identificato morfologicamente come An. gambiae (sl), identificato come non-An. stephensi con il test endpoint, ma il sequenziamento ha rilevato An. stephensi. L’analisi BLAST delle sequenze ITS2 ha ulteriormente identificato tutti i non-An. stephensi esemplari come An. arabiensis.
L’identificazione delle specie basata sulla sequenza era per lo più coerente con i risultati del test dell’endpoint ITS2, ad eccezione di un singolo campione identificato morfologicamente come An. gambiae (sl), identificato come non-An. stephensi con il test endpoint, ma il sequenziamento ha rilevato An. stephensi. L’analisi BLAST delle sequenze ITS2 ha ulteriormente identificato tutti i non-An. esemplari stephensi come An. arabiensis.
L’identificazione delle specie basata sulla sequenza era per lo più coerente con i risultati del test dell’endpoint ITS2, ad eccezione di un singolo campione identificato morfologicamente come An. gambiae (sl), identificato come non-An. stephensi con il test endpoint, ma il sequenziamento ha rilevato An. stephensi. L’analisi BLAST delle sequenze ITS2 ha ulteriormente identificato tutti i non-An. stephensi esemplari come An. arabiensis.

Tabella 3 Distribuzione di An. stephensi basato su habitat riproduttivi larvali
Da: distribuzione geografica di Anopheles stephensi nell’Etiopia orientale
| Regione | Posto | Habitat riproduttivo larvale | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Serbatoi di stoccaggio dell’acqua da costruzione ( n ) | Pneumatici scartati ( n ) | Fusti in acciaio ( n ) | Serbatoi d’acqua ( n ) | Serbatoi di stoccaggio temporaneo dell’acqua ( n ) | Tag ( n ) | Autolavaggio abbandonato ( n ) | Parte del veicolo scartata ( n ) | Secchio rotto ( n ) | Totale ( n ) | ||
| Somalo | Jigjiga | nf | 0 | 0 | 0 | 15 | 3 | nf | nf | nf | 18 |
| Erer | nf | 0 | nf | 5 | 100 | nf | 24 | nf | nf | 129 | |
| Kebridehar | nf | 0 | 0 | 0 | 700 | nf | nf | nf | 700 | ||
| Godey | nf | 0 | 0 | 0 | 116 | 224 | nf | nf | nf | 340 | |
| Degehabur | nf | 0 | 0 | 0 | 34 | 132 | nf | nf | nf | 166 | |
| Lontano | Semera | 225 | 5 | 0 | 17 | 13 | nf | nf | nf | 260 | |
| Gewane | 12 | 16 | 39 | 11 | nf | nf | 5 | 19 | 102 | ||
| Awash Sebat Kilo | 26 | nf | nf | 0 | nf | nf | nf | nf | 26 | ||
| Dire Dawa | Dire Dawa | 100 | 0 | 24 | 53 | 100 | nf | nf | nf | nf | 277 |
| Amhara | Bati | 11 | 7 | 2 | 5 | 110 | nf | 0 | nf | nf | 135 |
- Abbreviazioni : nf, non trovato durante l’indagine; n, numero di An. stephensi
Abbiamo confrontato l’identificazione morfologica con i risultati del test dell’endpoint PCR ITS2. Dei 130 Anopheles, 124 sono stati classificati come An. stephensi e sei come An. gambiae (sl) in base alla morfologia.
Cinque dei 124 (4,0%) identificati morfologicamente An. stephensi non è stato confermato per essere An. stephensi con il test endpoint PCR. Tutti identificati morfologicamente An. gambiae (sl) (An. arabiensis confermata dalla sequenza) che sono state amplificate con successo sono state identificate come non-An. stephensi con il test dell’endpoint ITS2 PCR.
Tutti gli 82 adulti catturati in natura An. stephensi sono stati sottoposti a screening per l’infezione da P. falciparum o P. vivax. Nessun DNA di Plasmodium è stato rilevato in nessuno dei campioni.
Discussione
Questo sondaggio conferma che An. stephensi è distribuito ampiamente nell’Etiopia orientale. Questi dati, presi con precedenti rapporti di An. stephensi a Kebridehar nel 2016 [9], confermano che An. stephensi è stabilito in questa regione. A nostra conoscenza, questa è la prima prova della presenza di un adulto An. stephensi in più regioni dell’Etiopia, dove potrebbe trasmettere la malaria.
La presenza capillare di An. stephensi in Etiopia insieme a Gibuti suggerisce che anche i paesi vicini, come il Sudan, il Sud Sudan, l’Eritrea, la Somalia e il Kenya, dovrebbero rafforzare la sorveglianza.
Dato che An. stephensi a Gibuti è stato trovato per trasportare sia P. falciparum che P. vivax [7, 8], c’è la possibilità che questi parassiti vengano osservati in Etiopia; quindi, le strategie di controllo della malaria dovrebbero ora considerare il potenziale dell’An. stephensi per trasmettere la malaria.
La presenza di An. stephensi è stato confermato utilizzando metodi morfologici e molecolari. Mentre la morfologia era per lo più coerente con l’approccio molecolare (119/124 identificato correttamente An. Stephensi), ci sono stati alcuni casi di identificazione errata basata sulla morfologia che evidenziava il rischio di errata identificazione dei campioni.
Poiché più programmi di sorveglianza vettoriale in Africa incorporano An. stephensi nelle loro chiavi morfologiche, i dati molecolari possono essere utili per la valutazione della formazione di successo in An. identificazione morfologica stephensi.
Ciò è particolarmente importante in questa fase iniziale di An. stephensi la sorveglianza mentre i tecnici sul campo si adatteranno alla rilevazione di An. stephensi.
Abbiamo incorporato due approcci molecolari, uno dei quali, il saggio end-point utilizzando primer progettati da Djadid et al. [13] è più fattibile in contesti con risorse limitate. Abbiamo scoperto che questo saggio era per lo più coerente con i dati della sequenza e deve essere potenzialmente integrato negli attuali saggi basati sulla PCR che si concentrano sul rilevamento dei membri dell’An. gambiae complex, i più comuni vettori della malaria in Africa.
Sebbene abbiamo confermato l’ampia distribuzione di An. stephensi nell’Etiopia orientale, la distribuzione nella parte occidentale del paese è ancora da determinare. L’Etiopia occidentale ha avuto una sorveglianza più consistente dei vettori della malaria rispetto a quella orientale, a causa del peso della malattia lì; tuttavia, i metodi di trapping utilizzati in precedenza possono limitare la capacità di rilevare An. stephensi.
Le attuali tecniche di trapping che fanno molto affidamento sulle trappole di luce CDC possono limitare la capacità di rilevare An. stephensi, dato il basso numero di An. stephensi catturato con trappole luminose CDC nella maggior parte dei siti in questo studio.
Ulteriori studi sull’allevamento, l’alimentazione e il comportamento a riposo di An. stephensi può fornire informazioni cruciali che possono essere applicate per migliorare i futuri sforzi di sorveglianza nell’Etiopia occidentale e orientale.
Diverse aree di interrogazione aggiuntive devono essere ulteriormente perseguite per informare meglio gli sforzi di controllo dei vettori. Nessuno studio è stato pubblicato per confermare che l’etiope An. stephensi può trasmettere o trasmette Plasmodium. Entrambi i campi di conferma di An. stephensi e infezioni di laboratorio sono approcci utili per valutare queste informazioni.
Nel presente studio, gli 82 esemplari di An. stephensi sono stati sottoposti a screening sia per P. falciparum che per P. vivax mediante PCR e Plasmodium non è stato rilevato. Ciò non è inaspettato poiché le regioni incluse in questo studio riportano una bassa trasmissione della malaria, quindi sarebbe necessaria una dimensione del campione molto più grande per rilevare l’infezione da Plasmodium in An. stephensi.
La sorveglianza futura continuerà a eseguire lo screening del Plasmodium utilizzando un saggio di immunoassorbimento enzimatico (ELISA) basato su PCR e circumsporozoite.
La sorveglianza di An. stephensi fino a questo punto è stato condotto in brevi intervalli di tempo, con limitata capacità di valutare i cambiamenti in An. dimensione della popolazione stephensi nel tempo.
Ripeteremo le raccolte in più siti nell’Etiopia orientale per fornire informazioni cruciali su come la popolazione cambia di anno in anno. Queste informazioni saranno particolarmente importanti quando verranno implementati nuovi interventi di controllo dei vettori per valutarne l’efficacia.
La resistenza agli insetticidi è stata segnalata nel vettore di malaria dominante An. arabiensis in Etiopia [17], ma lo stato di resistenza agli insetticidi in An. stephensi è sconosciuto. Sono in corso indagini sulla resistenza agli insetticidi, sui meccanismi molecolari alla base della resistenza e sui potenziali marcatori genetici che possono essere utilizzati per la sorveglianza.
Una domanda che rimane senza risposta riguarda l’origine di An. stephensi in Etiopia. La precedente analisi filogeografica ha rivelato la somiglianza della sequenza cox1 più vicina all’Etiopia An. stephensi trovato a Kebridehar da un esemplare proveniente dal Pakistan [9].
Analisi filogeografica incluso il sequenziamento dal recente An globale. Le raccolte stephensi che utilizzano più loci o sequenze dell’intero genoma possono aiutare a identificare l’origine esatta di An. stephensi nel Corno d’Africa e come si è diffuso in tutta la regione. Queste informazioni sosterranno gli sforzi per prevenire l’ulteriore introduzione e diffusione di An. stephensi.
Un’osservazione ausiliaria durante la sorveglianza è stata la rilevazione di larve del vettore dengue Ae. aegypti insieme ad An. stephensi larve, suggerendo che questi due vettori condividono habitat larvali. La dengue è una preoccupazione crescente per la salute pubblica in Etiopia, in particolare nell’Etiopia orientale, dove sono stati segnalati focolai gravi nel 2013 [18, 19] e nel 2015 [20].
Con il ritrovamento di Ae. aegypti larve con An. stephensi, possiamo considerare il controllo vettoriale integrato per indirizzare sia An. stephensi e Ae. aegypti. Questo sarebbe un approccio conveniente per ridurre la trasmissione del virus della malaria e della dengue. La futura sorveglianza nell’Etiopia orientale lavorerà per determinare l’abbondanza relativa di Ae. aegypti larve presso An. siti di riproduzione stephensi.
link di riferimento: https://parasitesandvectors.biomedcentral.com/articles/10.1186/s13071-020-3904-y
Bionomics
An . stephensi è una specie di laboratorio comune (come conseguenza della sua capacità di riprodursi in piccoli contenitori), tuttavia ci sono pochi dati quantificati che descrivono la bionomica di An. stephensi in natura. Nessun tasso di inoculazione entomologico riportato (EIR – il numero di punture di zanzara infette ricevute per unità di tempo) e solo un singolo valore di tasso di punture umane (HBR – il numero di punture di zanzara per unità di tempo) è stato trovato in letteratura (42).
In un breve riassunto della bionomica delle specie asiatiche, Sinka et al. (9) indicare An. stephensi preferisce principalmente nutrirsi di animali (zoofili) con esempi di morso umano (antropofagia) all’interno (endofagico) con comportamento a riposo all’aperto (endofilo), ma con esempi di comportamento morso all’aperto (esofagico), con una preferenza per mordere durante il crepuscolo (crepuscolare ) e la notte.
Simile a molte delle specie vettoriali dominanti asiatiche, An. stephensi sembra avere un certo livello di plasticità nel suo comportamento, in particolare nella scelta dell’ospite, con An rurale . Gli stephensi tendono ad essere più zoofili delle popolazioni urbane (42, 43). Il comportamento pungente della zanzara è determinato dalla disponibilità dell’ospite, inclusi modelli di attività umana e pratiche di ricovero del bestiame.
Si dice che le zanzare femmine nutrite preferiscano riposare in edifici mal costruiti o capannoni per animali (42⇓⇓-45). Tali strutture sono caratteristiche dei siti urbani non pianificati e possono essere trovate intorno e all’interno di molte città in rapida espansione in Africa (46, 47).
Molte indagini sui vettori della malaria includono catture di zanzare ogni ora tra le 18:00 e le 6:00, quando i vettori di anofelina sono tipicamente più attivi. Utilizzando i dati del database bionomico del Malaria Atlas Project (48), l’attività notturna per le tre specie di vettori africani primari ( An. Gambiae, An. Arabiensis e An. Funestus ) viene confrontata con An. stephensi (Fig.5).
Questa semplice trama illustra che An. stephensi ha il picco di attività (cioè, sta attivamente cercando un pasto di sangue) la sera prima di qualsiasi specie di vettore dominante africano (DVS) con oltre la metà dei morsi totali di questa specie che si verificano entro il primo quarto della notte (18:00 alle 21:00). L’attività può essere osservata in seguito, ma a un tasso molto inferiore (42).
È noto che le specie di vettori africani preferiscono mordere quando gli esseri umani sono addormentati e vulnerabili e, come previsto, An. gambiae e An. funestus hanno entrambi attività di punta fino a tarda notte (terzo trimestre: da mezzanotte alle 3 del mattino). Sia che si stabilisca di recente African An. Le popolazioni stephensi mantengono una preferenza per mordere la sera, resta da vedere, e ci sono alcune indicazioni che potrebbe cambiare la sua attività di mordere all’aperto a seconda della stagione (42).
Tuttavia, la sua preferenza per il morso crepuscolare consentirebbe ad An. stephensi per bypassare le reti da letto trattate con insetticida; gli interventi contro le zanzare più utilizzati (e attualmente più efficaci) utilizzati in tutta l’Africa (49, 50). An . Le larve stephensi sono state trovate coesistere in contenitori di stoccaggio dell’acqua con Aedes aegypti (un vettore di febbre gialla e dengue tra gli altri) e Culex quinquefasciatus (la zanzara della casa meridionale – vettore di filariosi linfatica e febbre del Nilo occidentale) quindi gli interventi già in atto per colpire questi le zanzare, così come una migliore gestione delle infrastrutture urbane, comprese l’acqua e gli spazi verdi, potrebbero aiutare a controllare An. stephensi (51⇓ – 53)

Un. stephensi è noto per avere un picco stagionale in abbondanza, normalmente nella tarda primavera / inizio estate (42, 43, 54), e mostra una certa regolazione dalle condizioni climatiche; qui abbiamo visto che la temperatura annuale è la covariata più influente nella descrizione della nicchia di questa specie. Nelle aree urbane, An. stephensi è stato trovato in grondaie, pozzi domestici, unità di condizionamento d’aria e cisterne (9, 52, 55).
Non dipende dalla pioggia per riempire i suoi siti larvali come molte altre specie (p. Es., An. Gambiae che fa uso di pozzanghere temporanee piene di pioggia), ma si avvale di strutture di stoccaggio dell’acqua costruite e mantenute dall’uomo, in particolare strutture in cemento o mattoni ( 19, 20, 42, 43, 52, 55, 56); una necessità per le popolazioni urbane umane durante i mesi estivi asciutti.
In effetti, è An. la capacità di stephensi di utilizzare tali serbatoi di stoccaggio dell’acqua costruiti dall’uomo che potrebbero aver facilitato la sua capacità di espandere la sua portata (15). All’interno del nostro modello, anche la densità della popolazione umana era una variabile molto influente e la nostra mappa finale dell’insieme suggerisce che molte grandi distese urbane sono altamente adatte se questa specie continua a invadere l’Africa, mentre la maggior parte delle aree rurali non lo è.
link di riferimento: https://www.pnas.org/content/early/2020/09/08/2003976117
Ulteriori informazioni: ME Sinka el al., “Un nuovo vettore della malaria in Africa: prevedere la gamma di espansione di Anopheles stephensi e identificare le popolazioni urbane a rischio”, PNAS (2020). www.pnas.org/cgi/doi/10.1073/pnas.2003976117



{kind=link}