










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A mix of ultrapotent antibodies from recovered COVID-19 patients has been shown to recognize and lock down the infection machinery of the pandemic coronavirus and keep it from entering cells. Each of the antibody types performs these overlapping tasks slightly differently.
Low doses of these antibodies, individually or as a cocktail, were also shown to protect hamsters from infection when exposed to the coronavirus by preventing it from replicating in their lungs.
An advantage of such cocktails is that they might also prevent the natural mutant forms of the virus that arose during this pandemic to escape treatment.
As some variants in the infection machinery have already been discovered during the coronavirus pandemic, using a mix of antibodies allows for neutralization of a broad spectrum of such viral variants.
In addition to preventing virus entry into host cells, the presence of the antibodies also seems to set off the infection-fighting actions of other immune cells, which arrive to clear out the virus.
“We believe that leveraging multiple, distinct, complementary mechanisms of action could provide additional benefits for clinical applications,” the researchers noted.
The researchers determined how the antibodies worked on a molecular level through cryo-electron microscopy studies of the resulting changes in the configuration of the virus infection machinery.
Besides directly preventing interactions with the host receptor, one of the two discovered antibodies locks the infection machinery in an inactive conformation, meaning it could not fuse with the host membrane on the surface of the cell. If unable to fuse, the coronavirus cannot break in and deliver its RNA to commandeer the cell.
The findings of this research are reported Sept. 24 in a rapid release paper in Science. Here is the paper.
The senior authors were Dr. Katja Fink of Vir Biotechnology and Dr. David Veesler, associate professor of biochemistry at the University of Washington School of Medicine. Veesler has studied the molecular structure and infection mechanisms of a variety of coronaviruses and other viruses.
The lead authors were M. Alejandra Tortorici of the UW Department of Biochemistry and the Institut Pasteur in Paris, and Martina Beltramello of Humab BioMed, a subsidiary of Vir Biotechnology in Switzerland.
Researchers from Washington University in St Louis, Rega Institute in Belgium, the University of Milan, Italy, and the University of Texas in Dallas also collaborated on the research.
Efficient therapeutic options are needed to control the spread of SARS-CoV-2 that has caused more than 978,000 fatalities worldwide. While the world awaits approved vaccines, pharmaceuticals to prevent or treat infections from the pandemic coronavirus are being sought that might be quicker to develop and test. These might both address the gap until vaccines are widely distributed, and still be needed for use after vaccines are available.
“Our results pave the way to implement antibody cocktails for prophylaxis or therapy that might have the advantage of circumventing or limiting the emergence of viral escape mutants,” the researchers noted. The antibody cocktail in their study needs to undergo trials in humans to determine safety and effectiveness.
SARS-CoV-2-specific T cells in patients with COVID-19
SARS-CoV-2-specific T cells have just started to be characterized for patients with COVID-1913,14 and their potential protective role has been inferred from studies of patients who recovered from SARS15 and MERS16. To study SARS-CoV-2-specific T cells associated with viral clearance, we collected peripheral blood from 36 individuals after recovery from mild to severe COVID-19 (demographic, clinical and virological information is included in Extended Data Table 1) and studied the T cell response against selected structural (N) and non-structural proteins (NSP7 and NSP13 of ORF1) of the large SARS-CoV-2 proteome (Fig. 1a).
We selected the N protein as it is one of the more-abundant structural proteins produced17 and has a high degree of homology between different betacoranaviruses18 (Extended Data Fig. 1).
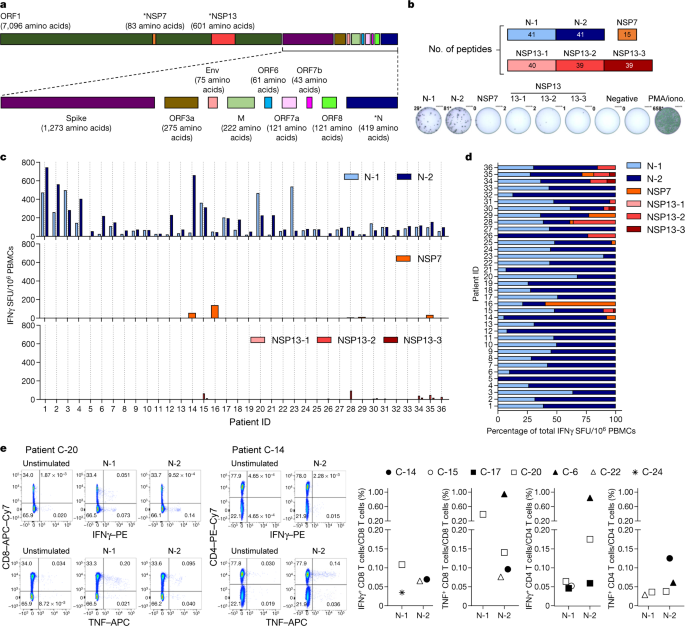
NSP7 and NSP13 were selected for their complete homology between SARS-CoV, SARS-CoV-2 and other animal coronaviruses that belong to the betacoranavirus genus12 (Extended Data Fig. 2), and because they are representative of the ORF1a/b polyprotein that encodes the replicase–transcriptase complex19.
This polyprotein is the first to be translated after infection with coronavirus and is essential for the subsequent transcription of the genomic and sub-genomic RNA species that encode the structural proteins19.
We synthesized 216 15-mer peptides that overlapped by 10 amino acids and that covered the whole length of NSP7 (83 amino acids), NSP13 (601 amino acids) and N (422 amino acids) and split these peptides into five pools of approximately 40 peptides each (N-1, N-2, NSP13-1, NSP13-2 and NSP13-3) and a single pool of 15 peptides that spanned NSP7 (Fig. 1b).
This unbiased method with overlapping peptides was used instead of bioinformatics selection of peptides, as the performance of such algorithms is often sub-optimal in Asian populations20.
Peripheral blood mononuclear cells (PBMCs) of 36 patients who recovered from COVID-19 were stimulated for 18 h with the different peptide pools and virus-specific responses were analysed by interferon-γ (IFNγ) ELISpot assay.
In all individuals tested (36 out of 36), we detected IFNγ spots after stimulation with the pools of synthetic peptides that covered the N protein (Fig. 1c, d).
In nearly all individuals, N-specific responses could be identified against multiple regions of the protein: 34 out of 36 individuals showed reactivity against the region that comprised amino acids 1–215 (N-1) and 36 out of 36 individuals showed reactivity against the region comprising amino acids 206–419 (N-2).
By contrast, responses to NSP7 and NSP13 peptide pools were detected at very low levels in 12 out of 36 COVID-19-convalescent individuals tested.
Direct ex vivo intracellular cytokine staining (ICS) was performed to confirm and define the N-specific IFNγ ELISpot response. Owing to their relative low frequency, N-specific T cells were more difficult to visualize by ICS than by ELISpot; however, a clear population of CD4 and/or CD8 T cells that produced IFNγ and/or TNF was detectable in seven out of nine analysed individuals (Fig. 1e and Extended Data Figs. 3, 4).
Moreover, despite the small sample size, we could compare the frequency of SARS-CoV-2-specific IFNγ spots with the presence of virus-neutralizing antibodies, the duration of infection and disease severity and found no correlations (Extended Data Fig. 5).
To confirm and further delineate the multi-specificity of the N-specific responses detected ex vivo in patients who recovered from COVID-19, we mapped the precise regions of the N protein that is able to activate IFNγ responses in nine individuals.
We organized the 82 overlapping peptides that covered the entire N protein into small peptide pools (of 7–8 peptides) that were used to stimulate PBMCs either directly ex vivo or after an in vitro expansion protocol that has previously been used for patients with hepatitis B virus21 or SARS22.
A schematic representation of the peptide pools is shown in Fig. 2a. We found that 8 out of 9 patients who recovered from COVID-19 had PBMCs that recognized multiple regions of the N protein of SARS-CoV-2 (Fig. 2a). Notably, we then defined single peptides that were able to activate T cells in seven patients. Using a peptide matrix strategy22, we first deconvolved the individual peptides that were responsible for the detected response by IFNγ ELISpot.
Subsequently, we confirmed the identity of the single peptides by testing—using ICS—the ability of the peptides to activate CD4 or CD8 T cells (Table 1 and Fig. 2b). Table 1 summarizes the different T cell epitopes that were defined by both ELISpot and ICS for seven individuals who recovered from COVID-19.
Notably, we observed that COVID-19-convalescent individuals developed T cells that were specific to regions that were also targeted by T cells from individuals who recovered from SARS. For example, the region of amino acids 101–120 of the N protein, which is a previously described CD4 T cell epitope in SARS-CoV-exposed individuals11,22, also stimulated CD4 T cells in two COVID-19-convalescent individuals.
Similarly, the region of amino acids 321–340 of the N protein contained epitopes that triggered CD4 and CD8 T cells in patients who recovered from either COVID-19 or from SARS22. The finding that patients who recovered from COVID-19 and SARS can mount T cell responses against shared viral determinants suggests that previous SARS-CoV infection can induce T cells that are able to cross-react against SARS-CoV-2.
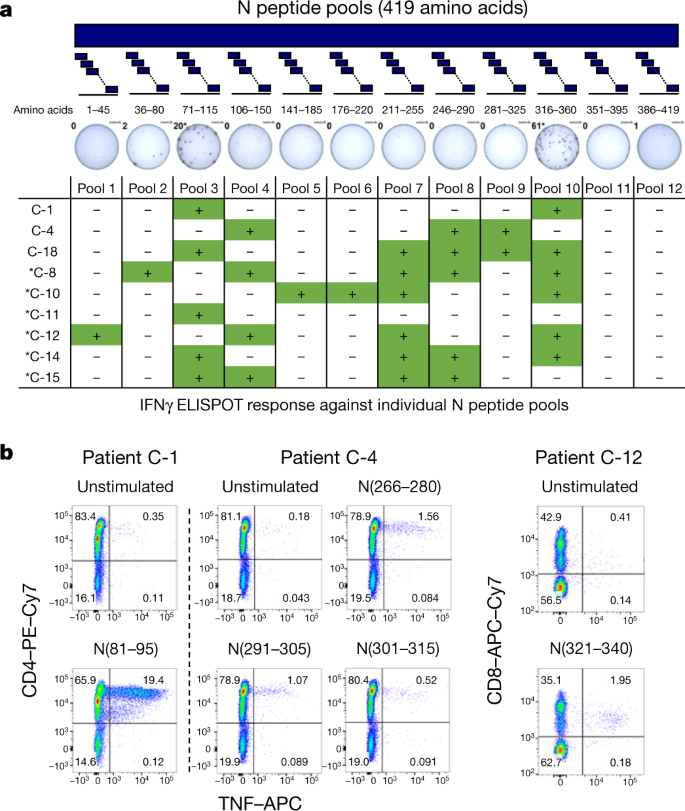
Table 1 SARS-CoV-2-specific T cell epitopes
From: SARS-CoV-2-specific T cell immunity in cases of COVID-19 and SARS, and uninfected controls
| Participants | T cell phenotype | Protein (amino acid residues) | SARS-CoV-2 amino acid sequence | SARS-CoV amino acid sequence |
|---|---|---|---|---|
| C-1 | CD4 | N (81–95) | DDQIGYYRRATRRIR | DDQIGYYRRATRRVR |
| CD8 | N (321–340) | GMEVTPSGTWLTYTGAIKLD | GMEVTPSGTWLTYHGAIKLD | |
| C-4 | CD4 | N (266–280) | KAYNVTQAFGRRGPE | KQYNVTQAFGRRGPE |
| CD4 | N (291–305) | LIRQGTDYKHWPQIA | LIRQGTDYKHWPQIA | |
| CD4 | N (301–315) | WPQIAQFAPSASAFF | WPQIAQFAPSASAFF | |
| C-8 | CD4 | N (51–65) | SWFTALTQHGKEDLK | SWFTALTQHGKEELR |
| CD4 | N (101–120) | MKDLSPRWYFYYLGTGPEAG | MKELSPRWYFYYLGTGPEAS | |
| C-10 | CD4 and CD8 | N (321–340) | GMEVTPSGTWLTYTGAIKLD | GMEVTPSGTWLTYHGAIKLD |
| C-12 | CD8 | N (321–340) | GMEVTPSGTWLTYTGAIKLD | GMEVTPSGTWLTYHGAIKLD |
| C-15 | CD4 | N (101–120) | MKDLSPRWYFYYLGTGPEAG | MKELSPRWYFYYLGTGPEAS |
| C-16 | CD4 | NSP7 (21–35) | RVESSSKLWAQCVQL | RVESSSKLWAQCVQL |
- T cells that react with distinct peptides were identified by IFNγ ELISpot and confirmed by ICS. Previously described T cell epitopes for SARS-CoV are highlighted in bold; non-conserved amino acid residues between SARS-CoV and SARS-CoV-2 are underlined.
SARS-CoV-2-specific T cells in patients with SARS
For the management of the current pandemic and for vaccine development against SARS-CoV-2, it is important to understand whether acquired immunity will be long-lasting. We have previously demonstrated that patients who recovered from SARS have T cells that are specific to epitopes within different SARS-CoV proteins that persist for 11 years after infection11.
Here, we collected PBMCs 17 years after SARS-CoV infection and tested whether they still contained cells that were reactive against SARS-CoV and whether these had cross-reactive potential against SARS-CoV-2 peptides. PBMCs from individuals who had resolved a SARS-CoV infection (n = 15) were stimulated directly ex vivo with peptide pools that covered the N protein of SARS-CoV (N-1 and N-2), NSP7 and NSP13 (Fig. 3a).
This revealed that 17 years after infection, IFNγ responses to SARS-CoV peptides were still present and were almost exclusively focused on the N protein rather than the NSP peptide pools (Fig. 3b). Subsequently, we tested whether the N peptides of SARS-CoV-2 (amino acid identity, 94%) induced IFNγ responses in PBMCs from individuals who resolved a SARS-CoV infection.
Indeed, PBMCs from all 23 individuals tested reacted to N peptides from SARS-CoV-2 (Fig. 3c, d). To test whether these low-frequency responses in individuals who had recovered from SARS could expand after encountering the N protein of SARS-CoV-2, the quantity of IFNγ-producing cells that responded to the N, NSP7 and NSP13 proteins of SARS-CoV-2 was analysed after 10 days of cell culture in the presence of the relevant peptides.
Seven out of eight individuals tested showed clear, robust expansion of N-reactive cells (Fig. 3e) and ICS confirmed that individuals who recovered from SARS had SARS-CoV N-reactive CD4 and CD8 memory T cells11 (Extended Data Fig. 6). In contrast to the response to the N peptides, we could not detect any cells that reacted to the peptide pools that covered NSP13 and only cells from one out of eight individuals reacted to NSP7 (Fig. 3e).
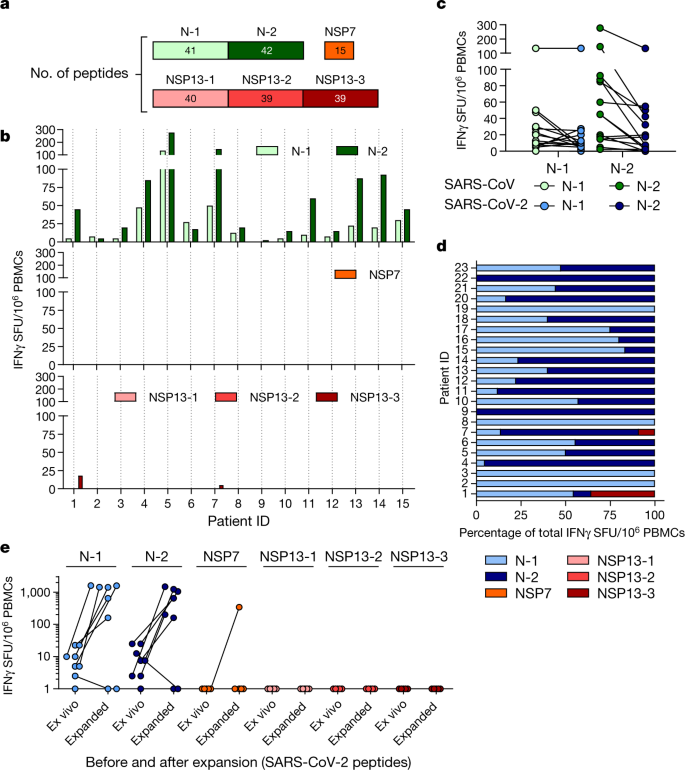
Thus, SARS-CoV-2 N-specific T cells are part of the T cell repertoire of individuals with a history of SARS-CoV infection and these T cells are able to robustly expand after encountering N peptides of SARS-CoV-2.
These findings demonstrate that virus-specific T cells induced by infection with betacoronaviruses are long-lasting, supporting the notion that patients with COVID-19 will develop long-term T cell immunity. Our findings also raise the possibility that long-lasting T cells generated after infection with related viruses may be able to protect against, or modify the pathology caused by, infection with SARS-CoV-2.
SARS-CoV-2-specific T cells in unexposed donors
To explore this possibility, we tested N-, NSP7- and NSP13-peptide-reactive IFNγ responses in 37 donors who were not exposed to SARS-CoV and SARS-CoV-2. Donors were either sampled before July 2019 (n = 26) or were serologically negative for both SARS-CoV-2 neutralizing antibodies and SARS-CoV-2 N antibodies23 (n = 11).
Different coronaviruses known to cause common colds in humans such as OC43, HKU1, NL63 and 229E present different degrees of amino acid homology with SARS-CoV-2 (Extended Data Fig. 1 and 2) and recent data have shown the presence of SARS-CoV-2 cross-reactive CD4 T cells (mainly specific to the spike protein) in donors who were not exposed to SARS-CoV-214.
Notably, we detected SARS-CoV-2-specific IFNγ responses in 19 out of 37 unexposed donors (Fig. 4a, b). The cumulative proportion of all studied individuals who responded to peptides covering the N protein and the ORF1-encoded NSP7 and NSP13 proteins is shown in Fig. 4b.
Unexposed donors showed a distinct pattern of reactivity; whereas individuals who recovered from COVID-19 and SARS reacted preferentially to N peptide pools (66% of individuals who recovered from COVID-19 and 91% of individuals who recovered from SARS responded to only the N peptide pools), the unexposed group showed a mixed response to the N protein or to NSP7 and NSP13 (Fig. 4a–c).
In addition, whereas NSP peptides stimulated a dominant response in only 1 out of 59 individuals who had resolved COVID-19 or SARS, these peptides triggered dominant reactivity in 9 out of 19 unexposed donors with SARS-CoV-2-reactive cells (Fig. 4c and Extended Data Fig. 7).
These SARS-CoV-2-reactive cells from unexposed donors had the capacity to expand after stimulation with SARS-CoV-2-specific peptides (Fig. 4d). We next delineated the SARS-CoV-2-specific response detected in unexposed donors in more detail.
Characterization of the N-specific response in one donor (H-2) identified CD4 T cells that were reactive to an epitope within the region of amino acids 101–120 of the N protein. This epitope was also detected in patients who recovered from COVID-19 and SARS8,22 (Fig. 2b).
This region has a high degree of homology to the sequences of the N protein of MERS-CoV, OC43 and HKU1 (Fig. 4e). In the same donor, we analysed PBMCs collected at multiple time points, demonstrating the persistence of the response to the 101–120 amino acid region of the N protein over 1 year (Extended Data Fig. 8a).
In three other donors who were not exposed to SARS-CoV or SARS-CoV-2, we identified CD4 T cells specific to the region of amino acids 26–40 of NSP7 (SKLWAQCVQLHNDIL; donor H-7) and CD8 T cells specific to an epitope comprising the region of amino acids 36–50 of NSP7 (HNDILLAKDTTEAFE; H-3, H-21; Fig. 4e, Extended Data Fig. 8b).
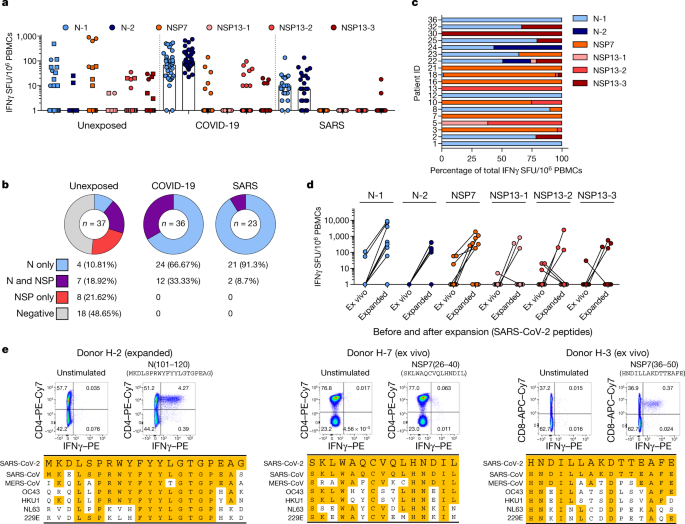
These latter two T cell specificities were of particular interest as the homology between the two protein regions of SARS-CoV, SARS-CoV-2 and other common cold coronaviruses (OC43, HKU1 NL63 and 229E) was minimal (Fig. 4e), especially for the CD8 T cell epitope. Indeed, the low-homology peptides that covered the sequences of the common cold coronaviruses failed to stimulate PBMCs from individuals with T cells responsive to amino acids 36–50 of NSP7 (Extended Data Fig. 8c).
Even though we cannot exclude that some SARS-CoV-2-reactive T cells might be naive or induced by completely unrelated pathogens5, this finding suggests that unknown coronaviruses, possibly of animal origin, might induce cross-reactive SARS-CoV-2 T cells in the general population.
We further characterized the NSP7-specific CD4 and CD8 T cells that were present in the three unexposed individuals. The reactive T cells expanded efficiently in vitro and mainly produced either both IFNγ and TNF (CD8 T cells) or only IFNγ (CD4 T cells) (Extended Data Fig. 9a).
We also determined that the CD8 T cells that were specific to amino acids 36–50 of NSP7 were HLA-B35-restricted and had an effector memory/terminal differentiated phenotype (CCR7−CD45RA+/−) (Extended Data Fig. 9b, c).
Conclusions
It is unclear why NSP7- and NSP13-specific T cells are detected and often dominant in unexposed donors, while representing a minor population in individuals who have recovered from SARS or COVID-19.
It is, however, consistent with the findings of a previous study11, in which ORF1-specific T cells were preferentially detected in some donors who were not exposed to SARS-CoV-2 whereas T cells from individuals who had recovered from COVID-19 preferentially recognized structural proteins.
Induction of virus-specific T cells in individuals who were exposed but uninfected has been demonstrated in other viral infections24,25,26. Theoretically, individuals exposed to coronaviruses might just prime ORF1-specific T cells, as the ORF1-encoded proteins are produced first in coronavirus-infected cells and are necessary for the formation of the viral replicase–transcriptase complex that is essential for the subsequent transcription of the viral genome, which then leads to the expression of various RNA species18.
Therefore, ORF1-specific T cells could hypothetically abort viral production by lysing SARS-CoV-2-infected cells before the formation of mature virions. By contrast, in patients with COVID-19 and SARS, the N protein—which is abundantly produced in cells that secrete mature virions17—would be expected to preferentially boost N-specific T cells.
Notably, the ORF1 region contains domains that are highly conserved among many different coronaviruses9. The distribution of these viruses in different animal species might result in periodic human contact that induces ORF1-specific T cells with cross-reactive abilities against SARS-CoV-2.
Understanding the distribution, frequency and protective capacity of pre-existing structural or non-structural protein-associated SARS-CoV-2 cross-reactive T cells could be important for the explanation of some of the differences in infection rates or pathology observed during this pandemic.
T cells that are specific to viral proteins are protective in animal models of airway infections27,28, but the possible effects of pre-existing N- and/or ORF1-specific T cells onthe differential modulation of SARS-CoV-2 infection will have to be carefully evaluated.
reference link : https://www.nature.com/articles/s41586-020-2550-z#MOESM4
More information: M. Alejandra Tortorici et al. Ultrapotent human antibodies protect against SARS-CoV-2 challenge via multiple mechanisms, Science (2020). DOI: 10.1126/science.abe3354



{kind=link}