










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Secondo un nuovo studio condotto dai ricercatori dell’Università dell’Arizona , il SARS-CoV-2, il virus che causa il COVID-19 , può alleviare il dolore.
La scoperta potrebbe spiegare perché quasi la metà delle persone che contraggono il COVID-19 manifesta pochi o nessun sintomo, anche se sono in grado di diffondere la malattia, secondo l’autore corrispondente dello studio Rajesh Khanna, Ph.D., professore presso il College of Medicina — Dipartimento di Farmacologia di Tucson.
“Per me aveva molto senso che forse la ragione per la diffusione inesorabile di COVID-19 è che nelle prime fasi, stai andando in giro tutto bene come se nulla fosse sbagliato perché il tuo dolore è stato soppresso”, ha detto il dott. Khanna.
“Hai il virus, ma non ti senti male perché il dolore è sparito. Se possiamo dimostrare che questo sollievo dal dolore è ciò che sta causando l’ulteriore diffusione di COVID-19, è di enorme valore “.
I centri statunitensi per il controllo e la prevenzione delle malattie hanno rilasciato dati aggiornati il 10 settembre secondo cui il 50% della trasmissione di COVID-19 si verifica prima della comparsa dei sintomi e il 40% delle infezioni da COVID-19 sono asintomatiche.
“Questa ricerca solleva la possibilità che il dolore, come un sintomo precoce di COVID-19, possa essere ridotto dalla proteina spike SARS-CoV-2 in quanto silenzia le vie di segnalazione del dolore del corpo”, ha detto il vicepresidente senior di UArizona Health Sciences Michael D. Dake, MD.
“I ricercatori dell’Università dell’Arizona di Scienze della salute presso il Comprehensive Pain and Addiction Center stanno sfruttando questa scoperta unica per esplorare una nuova classe di terapie per il dolore mentre continuiamo a cercare nuovi modi per affrontare l’epidemia di oppioidi”.
I virus infettano le cellule ospiti attraverso i recettori proteici sulle membrane cellulari.
All’inizio della pandemia, gli scienziati hanno stabilito che la proteina spike SARS-CoV-2 utilizza il recettore dell’enzima di conversione dell’angiotensina 2 (ACE2) per entrare nel corpo.
Ma a giugno, due documenti pubblicati sul server di preprint bioRxiv hanno indicato la neuropilina-1 come secondo recettore per SARS-CoV-2.
“Questo ha attirato la nostra attenzione perché negli ultimi 15 anni il mio laboratorio ha studiato un complesso di proteine e percorsi che si riferiscono all’elaborazione del dolore che sono a valle della neuropilina”, ha detto il dottor Khanna, affiliato alla UArizona Health Sciences Comprehensive Pain and Addiction Center ed è membro dell’UArizona BIO5 Institute.
“Così abbiamo fatto un passo indietro e ci siamo resi conto che questo potrebbe significare che forse la proteina spike è coinvolta in una sorta di elaborazione del dolore”.
Molti percorsi biologici segnalano al corpo di provare dolore. Uno è attraverso una proteina denominata fattore di crescita endoteliale vascolare-A (VEGF-A), che svolge un ruolo essenziale nella crescita dei vasi sanguigni ma è stata anche collegata a malattie come il cancro, l’artrite reumatoide e, più recentemente, COVID-19.
Come una chiave in una serratura, quando il VEGF-A si lega al recettore della neuropilina, dà inizio a una cascata di eventi con conseguente ipereccitabilità dei neuroni, che porta al dolore. Il dottor Khanna e il suo gruppo di ricerca hanno scoperto che la proteina spike SARS-CoV-2 si lega alla neuropilina esattamente nella stessa posizione del VEGF-A.
Con queste conoscenze, hanno eseguito una serie di esperimenti in laboratorio e su modelli di roditori per testare la loro ipotesi che la proteina spike SARS-CoV-2 agisca sulla via del dolore VEGF-A / neuropilina.
Hanno usato il VEGF-A come trigger per indurre l’eccitabilità dei neuroni, che crea dolore, quindi hanno aggiunto la proteina spike SARS-CoV-2.
“La proteina spike ha completamente invertito la segnalazione del dolore indotta da VEGF”, ha detto il dottor Khanna. “Non importava se usavamo dosi molto alte di picco o dosi estremamente basse: ha invertito completamente il dolore.”
Il dottor Khanna sta collaborando con immunologi e virologi di UArizona Health Sciences per continuare la ricerca sul ruolo della neuropilina nella diffusione del COVID-19.
Nel suo laboratorio, esaminerà la neuropilina come nuovo obiettivo per il sollievo dal dolore non oppioide. Durante lo studio, il dottor Khanna ha testato gli inibitori della neuropilina a piccole molecole esistenti sviluppati per sopprimere la crescita del tumore in alcuni tipi di cancro e ha scoperto che fornivano lo stesso sollievo dal dolore della proteina spike SARS-CoV-2 quando si legava alla neuropilina.
“Stiamo andando avanti con la progettazione di piccole molecole contro la neuropilina, in particolare composti naturali, che potrebbero essere importanti per alleviare il dolore”, ha detto il dottor Khanna. “Abbiamo una pandemia e un’epidemia di oppioidi. Si stanno scontrando. I nostri risultati hanno enormi implicazioni per entrambi. SARS-CoV-2 ci sta insegnando la diffusione virale, ma COVID-19 ci fa anche considerare la neuropilina come un nuovo metodo non oppioide per combattere l’epidemia di oppioidi “.
La sindrome respiratoria acuta grave coronavirus 2 (SARS-CoV-2) è l’agente eziologico del COVID-19, una malattia da coronavirus che, al 24 agosto, ha infettato più di 23,5 milioni di persone e ha causato quasi 810.000 decessi in tutto il mondo [15].
La maggior parte dei pazienti infettati da SARS-CoV-2 riporta una malattia respiratoria da lieve a grave con sintomi quali febbre, tosse e mancanza di respiro [30]. D’altra parte, un sottogruppo di pazienti a cui viene diagnosticato un test positivo per i nuclei acidi ma che sono asintomatici o minimamente sintomatici [30].
Prove crescenti dimostrano che gli individui asintomatici possono diffondere il virus in modo efficiente, e l’emergere di questi diffusori silenziosi di SARS-CoV-2 ha un controllo limitato della pandemia [14; 41].
Il dolore è una preoccupazione crescente nei pazienti sintomatici, probabilmente derivante da un attacco diretto di SARS-CoV-2 sulle cellule e dalla “tempesta di citochine” scatenata dalle cellule colpite [51; 68]. Non è noto se gli individui asintomatici o minimamente sintomatici abbiano soglie del dolore ridotte o se il loro dolore sia tacitato, ma entrambi potrebbero contribuire ad aumentare le dinamiche di trasmissione della malattia.
L’enzima di conversione dell’angiotensina 2 (ACE2) espresso in superficie è stato considerato il principale recettore per l’assorbimento di SARS-CoV-2 [22; 60; 64]. Evidenze emergenti indicano un sottoinsieme di neuroni sensoriali che esprimono ACE2 [48] che sinapsi con i neuroni del SNC spinali e del tronco cerebrale per produrre effetti neurologici, tra cui mal di testa e dolore ai nervi [32; 34].
Curiosamente, l’ACE2 non è presente nella maggior parte dei neuroni [48], nonostante l’aumento delle segnalazioni di sintomi neurologici comuni nei pazienti COVID-19 [32]. Paradossalmente, sebbene i livelli di espressione di ACE2 diminuiscano con l’invecchiamento [49], è stata osservata una maggiore gravità del COVID-19 nelle popolazioni di pazienti più anziane, come quella italiana [2], a sostegno della tesi secondo cui l’ACE2 non è l’unica porta d’ingresso della SARS -CoV-2 [1].
Due rapporti recenti hanno dimostrato che la proteina SARS-CoV-2 Spike può legarsi al dominio b1b2 del recettore della neuropilina-1 (NRP-1). Questa interazione avviene attraverso una sequenza di amminoacidi polibasici ( 682 RRAR 685 ), non conservata in SARS e MERS, chiamata motivo della “regola C-end” (CendR), che potenzia significativamente il suo ingresso nelle cellule [6; 11].
È importante sottolineare che le analisi “omiche” hanno rivelato una significativa sovraregolazione di NRP-1 nei campioni biologici di pazienti COVID-19 rispetto ai controlli sani [6]. Usando il fattore di crescita endoteliale vascolare-A (VEGF-A), un ligando fisiologico per la tasca b1b2 in NRP-1, abbiamo interrogato se la proteina Spike, il principale antigene di superficie di SARS-CoV-2, potesse bloccare VEGF-A / NRP -1 segnalazione per influenzare i comportamenti del dolore.
Dati i parallelismi tra gli effetti pro-nocicettivi del VEGF-A nei roditori [4; 58] e gli esseri umani [23; 58] e risultati clinici che dimostrano un aumento dei livelli di VEGF-A nel liquido di lavaggio alveolare bronchiale da pazienti COVID-19 [47] accoppiato con livelli sostanzialmente più bassi nel siero di individui asintomatici rispetto ai pazienti sintomatici [30], una domanda secondaria era verificare se La proteina Spike potrebbe conferire analgesia. Abbiamo scoperto che il VEGF-A sensibilizza l’attività dei nocicettori, un segno distintivo del dolore neuropatico [59], che è stato bloccato dalla proteina Spike e dall’inibitore NRP-1 EG00229 [26]. Inoltre, identifichiamo un nuovo ruolo analgesico per la proteina Spike, che è rispecchiato dall’inibizione di NRP-1.
Risultati
L’impegno specifico del ligando della segnalazione NRP-1 induce attività e dolore nocicettori
Inizialmente, abbiamo valutato il coinvolgimento di Spike e NRP-1 nel percorso VEGF-A / NRP-1. Un’interazione tra Spike (dominio S1 aa 16-685, contenente il motivo CendR 682 RRAR 685 ) e la porzione extracellulare di NRP-1 è stata confermata dal saggio di immunoassorbimento enzimatico (ELISA) (Fig. 1A).
Abbiamo calcolato che una costante di dissociazione di equilibrio (Kd) per questa interazione sia ∼166,2 nM (Fig. 1A). Successivamente, abbiamo placcato i neuroni sensoriali su array di microelettrodi multipozzetto (MEA), un approccio che consente misurazioni multiplex di potenziali d’azione extracellulari spontanei e stimolati da grandi popolazioni di cellule [9].
VEGF-A ha aumentato l’attivazione spontanea dei neuroni del ganglio della radice dorsale (DRG), che è stata bloccata dal dominio S1 della proteina Spike e dall’inibitore NRP-1 EG00229 (Fig. 1A). Al contrario, ligandi VEGF-B (ligando per VEGFR1 – un co-recettore per NRP-1 [31]) e semaforina 3A (Sema3A, ligando per recettore plexina – anche un co-recettore per NRP-1) [13; 53]) non ha influenzato l’attivazione spontanea dei nocicettori (Fig. 1B, C).
La mancanza di effetto di VEGF-B e Sema3A esclude un ruolo per VEGF-R1 e plexina, rispettivamente, implicando così un nuovo ligando, VEGF-A e recettore, NRP-1, percorso specifico che guida l’attivazione dei nocicettori (Fig. 1D).

Poiché sia il VEGF-A che la proteina Spike condividono una sacca di legame comune su NRP-1 (Fig. 1C) [6; 11; 43], abbiamo chiesto se la proteina Spike potesse bloccare la segnalazione di VEGF-A / NRP-1 per influenzare i comportamenti del dolore. Coerentemente con le relazioni precedenti [4; 58], abbiamo confermato che VEGF-A è pro-nocicettivo in quanto l’iniezione intra-plantare di VEGF-A ha ridotto sia le soglie di ritiro della zampa (Fig. 2A, B e Tabella S1) che le latenze a uno stimolo termico (Fig. Tabella S1) nei ratti maschi.
Risultati simili sono stati ottenuti anche nelle femmine di ratto (Fig. 2E-H e Tabella S1). Impedendo al VEGF-A di legarsi a NRP-1 con l’inibitore NRP-1 EG00229 o Spike di attivare la segnalazione VEGF-A / NRP-1, attenuato l’allodinia meccanica e l’iperalgesia termica indotte dal solo VEGF-A (Fig.2 e Tabella S1 ). Né Spike né EG00229 da soli hanno avuto alcun effetto su questi comportamenti (Fig. 2 e Tabella S1) in entrambi i sessi. Insieme, questi dati forniscono prove funzionali che la segnalazione di VEGF-A / NRP-1 promuove un fenotipo simile al dolore sensibilizzando l’attività dei nocicettori (Fig. 1D).
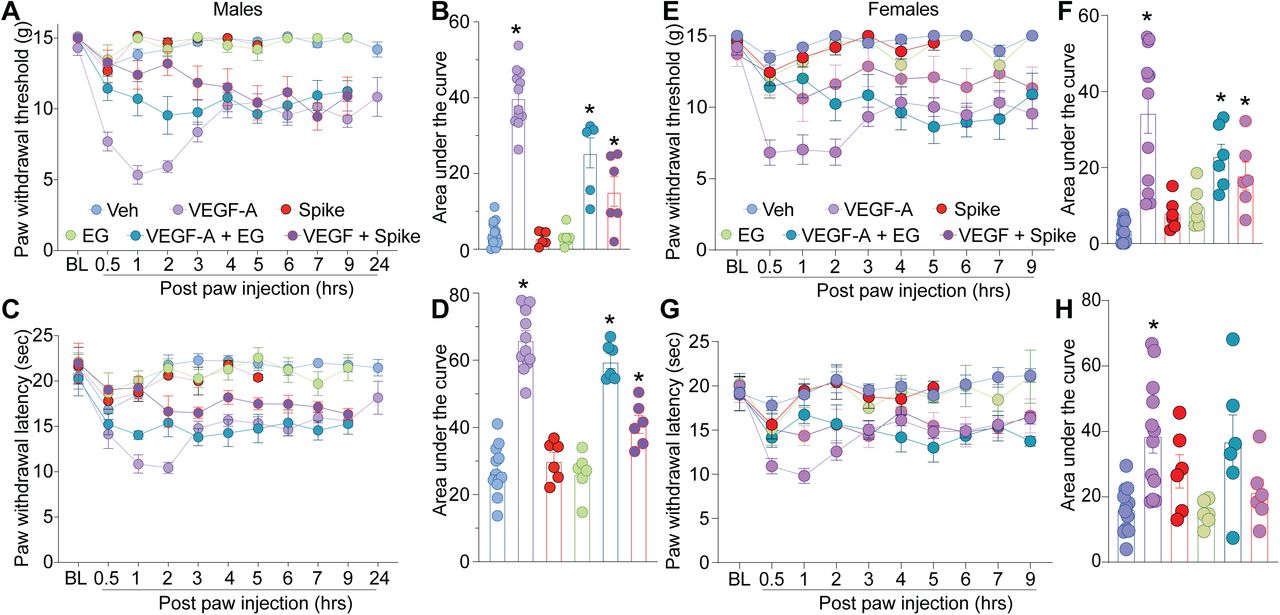
VEGF-A promuove un fenotipo simile al dolore che è bloccato dalla proteina Spike o dall’inibizione di NRP-1 nei ratti maschi e femmine.
Soglie di ritiro della zampa (A, B – maschio ed E, F – femmina) o latenze (C, D – maschio e G, H – femmina) per ratti naïve maschi e femmine a cui è stato iniettato VEGF-A (10 nM), Spike (100 nM), EG00229 (30 µM) o PBS (veicolo), da solo o in combinazione (50 µl / ratto; = 6-12). Per chiarezza, la significatività statistica non è presentata nei grafici dell’andamento temporale, ma è invece presentata nella Tabella S1. I pannelli B, F, D e H sono l’area sotto la curva per 0-24 ore. I dati sono mostrati come media ± sem e sono stati analizzati mediante analisi della varianza a due vie non parametrica in cui il tempo era il fattore entro il soggetto e il trattamento era il fattore tra i soggetti (post hoc: Sidak), * p <0,05. Le aree sotto la curva sono state confrontate mediante un’analisi della varianza unidirezionale con il test post-hoc di Kruskal-Wallis. Gli esperimenti sono stati analizzati da un investigatore cieco al trattamento.
Gli aumenti mediati da VEGF-A nelle correnti del canale ionico DRG sono normalizzati dalla rottura della segnalazione VEGF-A / NRP-1
Per ottenere informazioni sul meccanismo con cui il VEGF-A ha contribuito all’aumento dell’attività dei nocicettori, abbiamo postulato che i canali ionici nei DRG possono essere colpite, poiché contribuiscono alla plasticità nocicettiva [61].
Famiglie tipiche di correnti Na + da neuroni DRG di piccolo diametro sono mostrate nella Figura 3A. Il VEGF-A ha facilitato un aumento di 1,9 volte delle correnti totali di Na + rispetto ai DRG trattati con il veicolo (PBS), che erano completamente bloccati dalla proteina Spike (Fig. 3B, C). La proteina Spike da sola non ha influenzato le correnti di Na + (Fig. 3B, C e Tabella S1).
Poiché questa corrente diminuita potrebbe derivare da cambiamenti nel gate del canale, abbiamo determinato se la cinetica di attivazione e inattivazione delle correnti DRG Na + fosse influenzata. L’attivazione e l’inattivazione semimassimale (V1 / 2), così come i valori di pendenza (k) per l’attivazione e l’inattivazione, non erano diversi tra le condizioni testate (Fig. 3D, E e Tabelle S1, S2), ad eccezione di un ∼8 Spostamento iperpolarizzante del mV nell’inattivazione del canale del sodio indotta dal co-trattamento di VEGF-A e EG00229 (Tabella S2).
Risultati simili sono stati ottenuti per l’inibitore NRP-1 EG00229, che ha anche inibito l’aumento mediato da VEGF-A delle correnti totali di Na + (Fig. 3F-H e Tabella S1) ma non ha avuto effetto sulle proprietà biofisiche (Fig. 3I, J e Tabelle S1, S2).

Poiché i canali del calcio svolgono molteplici ruoli critici nella trasmissione e nell’elaborazione delle informazioni relative al dolore all’interno della via del dolore afferente primario [61], abbiamo valutato se fossero colpiti. Ci siamo concentrati sui canali di tipo N (CaV2.2) in quanto questi mediano il rilascio di neurotrasmettitori nelle sinapsi delle fibre afferenti nel corno dorsale e sono fondamentali nella matrice del dolore [50].
Il VEGF-A ha facilitato un aumento di 1,8 volte delle correnti totali di Ca2 + rispetto ai DRG trattati con il veicolo (PBS), che erano completamente bloccati dalla proteina Spike (Fig. 4A-C e Tabella S1). La sola proteina Spike non ha influenzato le correnti di Ca2 + (Fig. 4A-C). Inoltre, non abbiamo osservato alcun cambiamento nella cinetica di attivazione e inattivazione tra le condizioni testate (Fig. 4D, E e Tabelle S1, S2).
Risultati simili sono stati ottenuti per l’inibitore NRP-1 EG00229, che ha inibito l’aumento mediato da VEGF-A delle correnti di Ca2 + di tipo N (Fig. 4F-H e Tabella S1) ma non ha avuto effetto sulle proprietà biofisiche (Fig. 4I, J e tabelle S1, S2). Questi dati implicano la proteina Spike e NRP-1 nei canali Na + e Ca2 + (CaV2.2) nella segnalazione VEGF-A / NRP-1.

Il VEGF-A potenzia l’attività sinaptica nel corno dorsale lombare che è normalizzato dall’inibizione del segnale NRP-1 e della proteina Spike.
Il midollo spinale è un integratore della trasmissione sensoriale in cui i segnali nocicettivi in arrivo subiscono convergenza e modulazione [57]. La neurotrasmissione presinaptica spinale si basa sull’attivazione del potenziale d’azione dei neuroni DRG e sul rilascio dei neurotrasmettitori.
Da questi principi fisiologici fondamentali, oltre che dai risultati sopra descritti, siamo stati spinti a valutare se l’attività sinaptica fosse influenzata nel corno dorsale lombare.
Le ampiezze delle correnti spontanee eccitatorie postsinaptiche (sEPSC) dei neuroni nella regione della substantia gelatinosa del corno dorsale lombare non sono state influenzate dal VEGF-A (Fig. 5A, B e Tabella S1). Al contrario, l’applicazione di VEGF-A ha aumentato la frequenza di sEPSC di ∼3,6 volte, che è stata ridotta del ∼57% dall’inibizione di NRP-1 con EG00229 e ∼50% dalla proteina Spike (Fig. 5A, C e Tabella S1).
Le curve di distribuzione cumulativa dell’ampiezza e dell’intervallo tra eventi per le sEPSC sono mostrate nella Figura 5D, E. Rispetto ai controlli del veicolo, il VEGF-A, con o senza inibitore NRP-1 o proteina Spike, non ha avuto effetto sulla distribuzione dell’ampiezza cumulativa del EPSC spontanei (Fig. 5D e Tabella S1) ma ha cambiato la distribuzione della frequenza cumulativa di EPSC spontanei con intervalli tra eventi significativamente più lunghi (Fig. 5E e Tabella S1). Insieme, questi dati suggeriscono un meccanismo d’azione presinaptico della proteina Spike e NRP-1.

(A) Tracce rappresentative di correnti postsinaptiche eccitatorie spontanee (sEPSC) dai neuroni della substantia gelatinosa nel corno dorsale superficiale (lamina I / II) trattate per almeno 30 minuti con le condizioni indicate. Viene mostrato il riepilogo delle ampiezze (B) e delle frequenze (C) delle sEPSC per tutti i gruppi. Distribuzione cumulativa dell’ampiezza delle sEPSC (D) e dell’intervallo tra eventi (E) registrato dalle celle come indicato. La perfusione di 30 μM EG00229 ha ridotto la trasmissione sinaptica eccitatoria spontanea (AE) nei neuroni del corno dorsale lombare. I valori P dei confronti tra i trattamenti sono quelli indicati; per le analisi statistiche complete vedere la Tabella S1.
La proteina spike e l’inibizione di NRP-1 conferiscono anti-nocicezione nel modello di lesione del nervo risparmiato (SNI) del dolore neuropatico cronico
Abbiamo utilizzato il modello di lesione del nervo risparmiato (SNI) del dolore neuropatico, scelto perché produce un aumento affidabile e consistente della sensibilità al dolore [12], per valutare il potenziale di interruzione del percorso VEGF-A / NRP-1 per invertire la nocicezione. VEGF-A innesca l’autofosforilazione di VEGFR2 a Y1175 [54], servendo così come proxy per l’attivazione della segnalazione di VEGF-A. Nei ratti con SNI, l’applicazione intratecale di Spike, ha ridotto la fosforilazione di VEGFR2 (Y1175) sia sul lato controlaterale (non ferito) che omolaterale (ferito) (Fig. 6A, B). Ciò dimostra che Spike può inibire la segnalazione di VEGF-A in un modello di ratto di dolore neuropatico cronico. La lesione da SNI ha ridotto efficacemente le soglie di ritiro della zampa (PWT) (allodinia meccanica, Fig. 6C e Tabella S1) 10 giorni dopo l’infortunio. La somministrazione spinale della proteina Spike ha aumentato significativamente le PWT (Fig. 6C e Tabella S1), in modo dose-dipendente, per 5 ore. L’analisi dell’area sotto la curva (AUC) ha confermato l’inversione dose-dipendente dell’allodinia meccanica (Fig. 6D e Tabella S1) rispetto agli animali feriti trattati con veicolo. Risultati simili sono stati osservati con ratti femmina a cui è stato iniettato Spike (Fig. 6E, F). Infine, l’inibizione della segnalazione NRP-1 con EG00229 ha anche invertito le soglie di ritiro della zampa (Fig. 6G, H e Tabella S1).

RIFERIMENTI
[1].AlGhatrif M, Cingolani O, Lakatta EG. The Dilemma of Coronavirus Disease 2019, Aging, and Cardiovascular Disease: Insights From Cardiovascular Aging Science. JAMA Cardiology 2020.Google Scholar
[2].Asselta R, Paraboschi EM, Mantovani A, Duga S. ACE2 and TMPRSS2 variants and expression as candidates to sex and country differences in COVID-19 severity in Italy. Aging (Albany NY) 2020;12(11):10087–10098.Google Scholar
[3].Bates DO, Cui TG, Doughty JM, Winkler M, Sugiono M, Shields JD, Peat D, Gillatt D, Harper SJ. VEGF165b, an inhibitory splice variant of vascular endothelial growth factor, is down-regulated in renal cell carcinoma. Cancer research 2002;62(14):4123–4131.Abstract/FREE Full TextGoogle Scholar
[4].Beazley-Long N, Hua J, Jehle T, Hulse RP, Dersch R, Lehrling C, Bevan H, Qiu Y, Lagreze WA, Wynick D, Churchill AJ, Kehoe P, Harper SJ, Bates DO, Donaldson LF. VEGF-A165b is an endogenous neuroprotective splice isoform of vascular endothelial growth factor A in vivo and in vitro. The American journal of pathology 2013;183(3):918–929.CrossRefPubMedGoogle Scholar
[5].Brittain JM, Duarte DB, Wilson SM, Zhu W, Ballard C, Johnson PL, Liu N, Xiong W, Ripsch MS, Wang Y, Fehrenbacher JC, Fitz SD, Khanna M, Park CK, Schmutzler BS, Cheon BM, Due MR, Brustovetsky T, Ashpole NM, Hudmon A, Meroueh SO, Hingtgen CM, Brustovetsky N, Ji RR, Hurley JH, Jin X, Shekhar A, Xu XM, Oxford GS, Vasko MR, White FA, Khanna R. Suppression of inflammatory and neuropathic pain by uncoupling CRMP-2 from the presynaptic Ca(2)(+) channel complex. Nature medicine 2011;17(7):822–829.CrossRefPubMedGoogle Scholar
[6].Cantuti-Castelvetri L, Ojha R, Pedro LD, Djannatian M, Franz J, Kuivanen S, Kallio K, Kaya T, Anastasina M, Smura T, Levanov L, Szirovicza L, Tobi A, Kallio-Kokko H, Österlund P, Joensuu M, Meunier FA, Butcher S, Winkler MS, Mollenhauer B, Helenius A, Gokce O, Teesalu T, Hepojoki J, Vapalahti O, Stadelmann C, Balistreri G, Simons M. Neuropilin-1 facilitates SARS-CoV-2 cell entry and provides a possible pathway into the central nervous system. bioRxiv 2020:2020.2006.2007.137802.Google Scholar
[7].Cebe-Suarez S, Grunewald FS, Jaussi R, Li X, Claesson-Welsh L, Spillmann D, Mercer AA, Prota AE, Ballmer-Hofer K. Orf virus VEGF-E NZ2 promotes paracellular NRP-1/VEGFR-2 coreceptor assembly via the peptide RPPR. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 2008;22(8):3078–3086.Google Scholar
[8].Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. Journal of neuroscience methods 1994;53(1):55–63.CrossRefPubMedWeb of ScienceGoogle Scholar
[9].Chen Y, Moutal A, Navratilova E, Kopruszinski C, Yue X, Ikegami M, Chow M, Kanazawa I, Bellampalli SS, Xie J, Patwardhan A, Rice K, Fields H, Akopian A, Neugebauer V, Dodick D, Khanna R, Porreca F. The prolactin receptor long isoform regulates nociceptor sensitization and opioid-induced hyperalgesia selectively in females. Science translational medicine 2020;12(529).Google Scholar
[10].Choe W, Messinger RB, Leach E, Eckle VS, Obradovic A, Salajegheh R, Jevtovic-Todorovic V, Todorovic SM. TTA-P2 is a potent and selective blocker of T-type calcium channels in rat sensory neurons and a novel antinociceptive agent. MolPharmacol 2011;80(5):900–910.Abstract/FREE Full TextGoogle Scholar
[11].Daly JL, Simonetti B, Antón-Plágaro C, Kavanagh Williamson M, Shoemark DK, Simón-Gracia L, Klein K, Bauer M, Hollandi R, Greber UF, Horvath P, Sessions RB, Helenius A, Hiscox JA, Teesalu T, Matthews DA, Davidson AD, Cullen PJ, Yamauchi Y. Neuropilin-1 is a host factor for SARS-CoV-2 infection. bioRxiv 2020:2020.2006.2005.134114.Google Scholar
[12].Decosterd I, Woolf CJ. Spared nerve injury: an animal model of persistent peripheral neuropathic pain. Pain 2000;87(2):149–158.CrossRefPubMedWeb of ScienceGoogle Scholar
[13].Deo RC, Schmidt EF, Elhabazi A, Togashi H, Burley SK, Strittmatter SM. Structural bases for CRMP function in plexin-dependent semaphorin3A signaling. The EMBO journal 2004;23(1):9–22.Abstract/FREE Full TextGoogle Scholar
[14].Dhama K, Khan S, Tiwari R, Sircar S, Bhat S, Malik YS, Singh KP, Chaicumpa W, Bonilla-Aldana DK, Rodriguez-Morales AJ. Coronavirus Disease 2019-COVID-19. Clin Microbiol Rev 2020;33(4).Google Scholar
[15].Dong E, Du H, Gardner L. An interactive web-based dashboard to track COVID-19 in real time. The Lancet Infectious Diseases 2020;20(5):533–534.CrossRefPubMedGoogle Scholar
[16].Dustrude ET, Moutal A, Yang X, Wang Y, Khanna M, Khanna R. Hierarchical CRMP2 posttranslational modifications control NaV1.7 function. Proceedings of the National Academy of Sciences of the United States of America 2016;113(52):E8443–E8452.Abstract/FREE Full TextGoogle Scholar
[17].Dustrude ET, Wilson SM, Ju W, Xiao Y, Khanna R. CRMP2 protein SUMOylation modulates NaV1.7 channel trafficking. The Journal of biological chemistry 2013;288(34):24316–24331.Abstract/FREE Full TextGoogle Scholar
[18].Francois-Moutal L, Wang Y, Moutal A, Cottier KE, Melemedjian OK, Yang X, Wang Y, Ju W, Largent-Milnes TM, Khanna M, Vanderah TW, Khanna R. A membrane-delimited N-myristoylated CRMP2 peptide aptamer inhibits CaV2.2 trafficking and reverses inflammatory and postoperative pain behaviors. Pain 2015;156(7):1247–1264.CrossRefPubMedGoogle Scholar
[19].Friesner RA, Banks JL, Murphy RB, Halgren TA, Klicic JJ, Mainz DT, Repasky MP, Knoll EH, Shelley M, Perry JK, Shaw DE, Francis P, Shenkin PS. Glide: a new approach for rapid, accurate docking and scoring. 1. Method and assessment of docking accuracy. Journal of medicinal chemistry 2004;47(7):1739–1749.CrossRefPubMedWeb of ScienceGoogle Scholar
[20].Garcia J, Hurwitz HI, Sandler AB, Miles D, Coleman RL, Deurloo R, Chinot OL. Bevacizumab (Avastin®) in cancer treatment: A review of 15 years of clinical experience and future outlook. Cancer Treat Rev 2020;86:102017.Google Scholar
[21].Guo HF, Vander Kooi CW. Neuropilin Functions as an Essential Cell Surface Receptor. The Journal of biological chemistry 2015;290(49):29120–29126.Abstract/FREE Full TextGoogle Scholar
[22].Hoffmann M, Kleine-Weber H, Schroeder S, Kruger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH, Nitsche A, Muller MA, Drosten C, Pohlmann S. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020;181(2):271–280 e278.CrossRefPubMedGoogle Scholar
[23].Hulse RP. Role of VEGF-A in chronic pain. Oncotarget 2017;8(7):10775–10776.Google Scholar
[24].Hulse RP, Beazley-Long N, Hua J, Kennedy H, Prager J, Bevan H, Qiu Y, Fernandes ES, Gammons MV, Ballmer-Hofer K, Gittenberger de Groot AC, Churchill AJ, Harper SJ, Brain SD, Bates DO, Donaldson LF. Regulation of alternative VEGF-A mRNA splicing is a therapeutic target for analgesia. Neurobiology of disease 2014;71:245–259.CrossRefPubMedGoogle Scholar
[25].Ibrahim MM, Patwardhan A, Gilbraith KB, Moutal A, Yang X, Chew LA, Largent-Milnes T, Malan TP, Vanderah TW, Porreca F, Khanna R. Long-lasting antinociceptive effects of green light in acute and chronic pain in rats. Pain 2017;158(2):347–360.Google Scholar
[26].Jarvis A, Allerston CK, Jia H, Herzog B, Garza-Garcia A, Winfield N, Ellard K, Aqil R, Lynch R, Chapman C, Hartzoulakis B, Nally J, Stewart M, Cheng L, Menon M, Tickner M, Djordjevic S, Driscoll PC, Zachary I, Selwood DL. Small molecule inhibitors of the neuropilin-1 vascular endothelial growth factor A (VEGF-A) interaction. Journal of medicinal chemistry 2010;53(5):2215–2226.CrossRefPubMedGoogle Scholar
[27].Joseph EK, Green PG, Bogen O, Alvarez P, Levine JD. Vascular endothelial cells mediate mechanical stimulation-induced enhancement of endothelin hyperalgesia via activation of P2×2/3 receptors on nociceptors. The Journal of neuroscience : the official journal of the Society for Neuroscience 2013;33(7):2849–2859.Abstract/FREE Full TextGoogle Scholar
[28].Khanna R, Yu J, Yang X, Moutal A, Chefdeville A, Gokhale V, Shuja Z, Chew LA, Bellampalli SS, Luo S, Francois-Moutal L, Serafini MJ, Ha T, Perez-Miller S, Park KD, Patwardhan A, Streicher JM, Colecraft HM, Khanna M. Targeting the CaVα-β interaction yields an antagonist of the N-type CaV2.2 channel with broad antinociceptive efficacy. Pain 2019.Google Scholar
[29].Lindholm T, Risling M, Carlstedt T, Hammarberg H, Wallquist W, Cullheim S, Sköld MK. Expression of Semaphorins, Neuropilins, VEGF, and Tenascins in Rat and Human Primary Sensory Neurons after a Dorsal Root Injury. Frontiers in Neurology 2017;8(49).Google Scholar
[30].Long QX, Tang XJ, Shi QL, Li Q, Deng HJ, Yuan J, Hu JL, Xu W, Zhang Y, Lv FJ, Su K, Zhang F, Gong J, Wu B, Liu XM, Li JJ, Qiu JF, Chen J, Huang AL. Clinical and immunological assessment of asymptomatic SARS-CoV-2 infections. Nature medicine 2020.Google Scholar
[31].Makinen T, Olofsson B, Karpanen T, Hellman U, Soker S, Klagsbrun M, Eriksson U, Alitalo K. Differential binding of vascular endothelial growth factor B splice and proteolytic isoforms to neuropilin-1. The Journal of biological chemistry 1999;274(30):21217–21222.Abstract/FREE Full TextGoogle Scholar
[32].Mao L, Jin H, Wang M, Hu Y, Chen S, He Q, Chang J, Hong C, Zhou Y, Wang D, Miao X, Li Y, Hu B. Neurologic Manifestations of Hospitalized Patients With Coronavirus Disease 2019 in Wuhan, China. JAMA Neurol 2020.Google Scholar
[33].Mintz IM, Venema VJ, Swiderek KM, Lee TD, Bean BP, Adams ME. P-type calcium channels blocked by the spider toxin omega-Aga-IVA. Nature 1992;355(6363):827–829.CrossRefPubMedWeb of ScienceGoogle Scholar
[34].Montalvan V, Lee J, Bueso T, De Toledo J, Rivas K. Neurological manifestations of COVID-19 and other coronavirus infections: A systematic review. Clin Neurol Neurosurg 2020;194:105921.PubMedGoogle Scholar
[35].Moutal A, Cai S, Yu J, Stratton HJ, Chefdeville A, Gomez K, Ran D, Madura CL, Boinon L, Soto M, Zhou Y, Shan Z, Chew LA, Rodgers KA, Khanna R. Studies on CRMP2 SUMOylation-deficient transgenic mice identify sex-specific NaV1.7 regulation in the pathogenesis of chronic neuropathic pain. bioRxiv 2020:2020.2004.2020.049106.Google Scholar
[36].Moutal A, Chew LA, Yang X, Wang Y, Yeon SK, Telemi E, Meroueh S, Park KD, Shrinivasan R, Gilbraith KB, Qu C, Xie JY, Patwardhan A, Vanderah TW, Khanna M, Porreca F, Khanna R. (S)-lacosamide inhibition of CRMP2 phosphorylation reduces postoperative and neuropathic pain behaviors through distinct classes of sensory neurons identified by constellation pharmacology. Pain 2016;157(7):1448–1463.Google Scholar
[37].Moutal A, Li W, Wang Y, Ju W, Luo S, Cai S, Francois-Moutal L, Perez-Miller S, Hu J, Dustrude ET, Vanderah TW, Gokhale V, Khanna M, Khanna R. Homology-guided mutational analysis reveals the functional requirements for antinociceptive specificity of collapsin response mediator protein 2-derived peptides. British journal of pharmacology 2017.Google Scholar
[38].Moutal A, Wang Y, Yang X, Ji Y, Luo S, Dorame A, Bellampalli SS, Chew LA, Cai S, Dustrude ET, Keener JE, Marty MT, Vanderah TW, Khanna R. Dissecting the role of the CRMP2-neurofibromin complex on pain behaviors. Pain 2017;158(11):2203–2221.CrossRefPubMedGoogle Scholar
[39].Newcomb R, Szoke B, Palma A, Wang G, Chen X, Hopkins W, Cong R, Miller J, Urge L, Tarczy-Hornoch K, Loo JA, Dooley DJ, Nadasdi L, Tsien RW, Lemos J, Miljanich G. Selective peptide antagonist of the class E calcium channel from the venom of the tarantula Hysterocrates gigas. Biochemistry 1998;37(44):15353–15362.CrossRefPubMedWeb of ScienceGoogle Scholar
[40].Nowak DG, Woolard J, Amin EM, Konopatskaya O, Saleem MA, Churchill AJ, Ladomery MR, Harper SJ, Bates DO. Expression of pro-and anti-angiogenic isoforms of VEGF is differentially regulated by splicing and growth factors. Journal of cell science 2008;121(Pt 20):3487–3495.Abstract/FREE Full TextGoogle Scholar
[41].Oran DP, Topol EJ. Prevalence of Asymptomatic SARS-CoV-2 Infection: A Narrative Review. Annals of internal medicine 2020:M20–3012.Google Scholar
[42].Pacchioni AM, Vallone J, Worley PF, Kalivas PW. Neuronal pentraxins modulate cocaine-induced neuroadaptations. The Journal of pharmacology and experimental therapeutics 2009;328(1):183–192.Abstract/FREE Full TextGoogle Scholar
[43].Parker MW, Xu P, Li X, Vander Kooi CW. Structural basis for selective vascular endothelial growth factor-A (VEGF-A) binding to neuropilin-1. The Journal of biological chemistry 2012;287(14):11082–11089.Abstract/FREE Full TextGoogle Scholar
[44].Peach CJ, Mignone VW, Arruda MA, Alcobia DC, Hill SJ, Kilpatrick LE, Woolard J. Molecular Pharmacology of VEGF-A Isoforms: Binding and Signalling at VEGFR2. Int J Mol Sci 2018;19(4).Google Scholar
[45].Plein A, Fantin A, Ruhrberg C. Neuropilin regulation of angiogenesis, arteriogenesis, and vascular permeability. Microcirculation 2014;21(4):315–323.CrossRefPubMedWeb of ScienceGoogle Scholar
[46].Powell J, Mota F, Steadman D, Soudy C, Miyauchi JT, Crosby S, Jarvis A, Reisinger T, Winfield N, Evans G, Finniear A, Yelland T, Chou YT, Chan AWE, O’Leary A, Cheng L, Liu D, Fotinou C, Milagre C, Martin JF, Jia H, Frankel P, Djordjevic S, Tsirka SE, Zachary IC, Selwood DL. Small Molecule Neuropilin-1 Antagonists Combine Antiangiogenic and Antitumor Activity with Immune Modulation through Reduction of Transforming Growth Factor Beta (TGFbeta) Production in Regulatory T-Cells. Journal of medicinal chemistry 2018;61(9):4135–4154.PubMedGoogle Scholar
[47].Ray PR, Wangzhou A, Ghneim N, Yousuf MS, Paige C, Tavares-Ferreira D, Mwirigi JM, Shiers S, Sankaranarayanan I, McFarland AJ, Neerukonda SV, Davidson S, Dussor G, Burton MD, Price TJ. A pharmacological interactome between COVID-19 patient samples and human sensory neurons reveals potential drivers of neurogenic pulmonary dysfunction. Brain, behavior, and immunity 2020.Google Scholar
[48].Shiers S, Ray PR, Wangzhou A, Tatsui CE, Rhines L, Li Y, Uhelski ML, Dougherty PM, Price TJ. ACE2 expression in human dorsal root ganglion sensory neurons: implications for SARS-CoV-2 virus-induced neurological effects. bioRxiv 2020:2020.2005.2028.122374.Google Scholar
[49].Singh KK, Chaubey G, Chen JY, Suravajhala P. Decoding SARS-CoV-2 Hijacking of Host Mitochondria in Pathogenesis of COVID-19. American journal of physiology Cell physiology 2020.Google Scholar
[50].Snutch TP. Targeting chronic and neuropathic pain: the N-type calcium channel comes of age. NeuroRx 2005;2(4):662–670.Abstract/FREE Full TextGoogle Scholar
[51].Su S, Cui H, Wang T, Shen X, Ma C. Pain: A potential new label of COVID-19. Brain, behavior, and immunity 2020;87:159–160.Google Scholar
[52].Taiana MM, Lombardi R, Porretta-Serapiglia C, Ciusani E, Oggioni N, Sassone J, Bianchi R, Lauria G. Neutralization of schwann cell-secreted VEGF is protective to in vitro and in vivo experimental diabetic neuropathy. PloS one 2014;9(9):e108403.CrossRefPubMedGoogle Scholar
[53].Takahashi T, Fournier A, Nakamura F, Wang LH, Murakami Y, Kalb RG, Fujisawa H, Strittmatter SM. Plexin-neuropilin-1 complexes form functional semaphorin-3A receptors. Cell 1999;99(1):59–69.CrossRefPubMedWeb of ScienceGoogle Scholar
[54].Takahashi T, Yamaguchi S, Chida K, Shibuya M. A single autophosphorylation site on KDR/Flk-1 is essential for VEGF-A-dependent activation of PLC-gamma and DNA synthesis in vascular endothelial cells. The EMBO journal 2001;20(11):2768–2778.Abstract/FREE Full TextGoogle Scholar
[55].Takano S, Uchida K, Inoue G, Matsumoto T, Aikawa J, Iwase D, Mukai M, Miyagi M, Takaso M. Vascular endothelial growth factor expression and their action in the synovial membranes of patients with painful knee osteoarthritis. BMC Musculoskelet Disord 2018;19(1):204.Google Scholar
[56].Teesalu T, Sugahara KN, Kotamraju VR, Ruoslahti E. C-end rule peptides mediate neuropilin-1-dependent cell, vascular, and tissue penetration. Proceedings of the National Academy of Sciences of the United States of America 2009;106(38):16157–16162.Abstract/FREE Full TextGoogle Scholar
[57].Todd AJ. Neuronal circuitry for pain processing in the dorsal horn. Nat Rev Neurosci 2010;11(12):823–836.CrossRefPubMedGoogle Scholar
[58].Verheyen A, Peeraer E, Nuydens R, Dhondt J, Poesen K, Pintelon I, Daniels A, Timmermans JP, Meert T, Carmeliet P, Lambrechts D. Systemic anti-vascular endothelial growth factor therapies induce a painful sensory neuropathy. Brain : a journal of neurology 2012;135(Pt 9):2629–2641.CrossRefPubMedGoogle Scholar
[59].Walters ET. Adaptive mechanisms driving maladaptive pain: how chronic ongoing activity in primary nociceptors can enhance evolutionary fitness after severe injury. Philosophical transactions of the Royal Society of London Series B, Biological sciences 2019;374(1785):20190277.Google Scholar
[60].Wang Q, Zhang Y, Wu L, Niu S, Song C, Zhang Z, Lu G, Qiao C, Hu Y, Yuen KY, Wang Q, Zhou H, Yan J, Qi J. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020;181(4):894–904 e899.CrossRefPubMedGoogle Scholar
[61].Waxman SG, Zamponi GW. Regulating excitability of peripheral afferents: emerging ion channel targets. Nature neuroscience 2014;17(2):153–163.CrossRefPubMedGoogle Scholar
[62].Weekes CD, Beeram M, Tolcher AW, Papadopoulos KP, Gore L, Hegde P, Xin Y, Yu R, Shih LM, Xiang H, Brachmann RK, Patnaik A. A phase I study of the human monoclonal anti-NRP1 antibody MNRP1685A in patients with advanced solid tumors. Invest New Drugs 2014;32(4):653–660.CrossRefPubMedGoogle Scholar
[63].Yaksh TL, Rudy TA. Chronic catheterization of the spinal subarachnoid space. Physiology & behavior 1976;17(6):1031–1036.CrossRefPubMedWeb of ScienceGoogle Scholar
[64].Yan R, Zhang Y, Li Y, Xia L, Guo Y, Zhou Q. Structural basis for the recognition of SARS-CoV-2 by full-length human ACE2. Science 2020;367(6485):1444–1448.Abstract/FREE Full TextGoogle Scholar
[65].Yu J, Moutal A, Dorame A, Bellampalli SS, Chefdeville A, Kanazawa I, Pham NYN, Park KD, Weimer JM, Khanna R. Phosphorylated CRMP2 Regulates Spinal Nociceptive Neurotransmission. Molecular neurobiology 2018.Google Scholar
[66].Yu J, Moutal A, Dorame A, Bellampalli SS, Chefdeville A, Kanazawa I, Pham NYN, Park KD, Weimer JM, Khanna R. Phosphorylated CRMP2 Regulates Spinal Nociceptive Neurotransmission. Molecular neurobiology 2019;56(7):5241–5255.CrossRefPubMedGoogle Scholar
[67].Zeng Y, Han H, Tang B, Chen J, Mao D, Xiong M. Transplantation of Recombinant Vascular Endothelial Growth Factor (VEGF)189-Neural Stem Cells Downregulates Transient Receptor Potential Vanilloid 1 (TRPV1) and Improves Motor Outcome in Spinal Cord Injury. Med Sci Monit 2018;24:1089–1096.CrossRefPubMedGoogle Scholar
[68].Zhang X, Tan Y, Ling Y, Lu G, Liu F, Yi Z, Jia X, Wu M, Shi B, Xu S, Chen J, Wang W, Chen B, Jiang L, Yu S, Lu J, Wang J, Xu M, Yuan Z, Zhang Q, Zhang X, Zhao G, Wang S, Chen S, Lu H. Viral and host factors related to the clinical outcome of COVID-19. Nature 2020.Google Scholar
Source: University of Arizona



{kind=link}