








")


 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



In un articolo pubblicato oggi sulla rivista Cell, gli scienziati descrivono risultati inaspettati e indesiderabili dopo aver modificato i geni negli embrioni umani con CRISPR, un sistema di editing genomico.
Lo studio, l’analisi più dettagliata fino ad oggi di CRISPR negli embrioni umani, mostra che l’applicazione di CRISPR per riparare un gene che causa la cecità nelle prime fasi dello sviluppo di un embrione umano spesso elimina un intero cromosoma o una larga parte di esso.
“Il nostro studio mostra che CRISPR / Cas9 non è ancora pronto per l’uso clinico per correggere le mutazioni in questa fase dello sviluppo umano”, afferma l’autore senior dello studio Dieter Egli, assistente professore di biologia cellulare dello sviluppo presso il Dipartimento di Pediatria presso la Columbia University Vagelos College di medici e chirurghi.
CRISPR in embrioni umani
Il sistema di modifica del genoma chiamato CRISPR / Cas9 ha rivoluzionato la biologia molecolare negli ultimi anni e i suoi scopritori sono stati insigniti del Premio Nobel per la chimica di quest’anno. Il sistema consente agli scienziati di apportare modifiche precise ai genomi di cellule coltivate, tessuti viventi ed embrioni animali.
Il primo utilizzo di CRISPR in embrioni umani è stato segnalato nel 2015. Solo pochi anni dopo, nel 2018, He Jiankui ha affermato di aver eseguito la procedura su una coppia di gemelli, provocando una tempesta di condanna da parte di scienziati e funzionari governativi di tutto il mondo.
CRISPR può portare ad anomalie cromosomiche
Nel nuovo articolo, Egli ei suoi colleghi hanno testato gli effetti dell’editing genomico di CRISPR / Cas9 su embrioni umani in fase iniziale portatori di una mutazione in un gene chiamato EYS (omologa occhi chiusi), che causa cecità ereditaria.
“Sappiamo da studi precedenti su cellule umane differenziate e su topi che una rottura del DNA si traduce principalmente in due risultati: riparazione precisa o piccoli cambiamenti locali.
Nel gene EYS, questi cambiamenti possono produrre un gene funzionale, sebbene non sia una riparazione perfetta “, afferma Michael Zuccaro, Ph.D. candidato alla Columbia University Vagelos College of Physicians and Surgeons e co-autore principale dell’articolo.
Quando Zuccaro, Egli ei loro colleghi hanno esaminato l’intero genoma degli embrioni modificati in questo studio, hanno identificato un altro risultato.
“Abbiamo imparato che nelle cellule embrionali umane , una singola rottura nel DNA può provocare un terzo esito, la perdita di un intero cromosoma o talvolta di un ampio segmento di quel cromosoma, e questa perdita del cromosoma è molto frequente”, dice Zuccaro.
Nel 2017, uno studio ha riportato la corretta correzione di una mutazione che causa malattie cardiache in embrioni umani normali utilizzando CRISPR.
I dati completi del nuovo studio offrono un’interpretazione diversa di questi risultati: invece di essere corretto, il cromosoma portatore della mutazione potrebbe essere stato perso del tutto.
“Se i nostri risultati fossero stati noti due anni fa, dubito che qualcuno sarebbe andato avanti con un tentativo di utilizzare CRISPR per modificare un gene in un embrione umano nella clinica”, dice Egli.
“La nostra speranza è che questi risultati cautelativi dovrebbero scoraggiare l’applicazione clinica prematura di questa importante tecnologia, ma possono anche guidare la ricerca responsabile per ottenere il suo uso sicuro ed efficace”.
Lo studio è stato pubblicato il 29 ottobre su Cell, in un documento intitolato “Rimozione del cromosoma specifico dell’allele dopo la scissione di Cas9 negli embrioni umani”.
La modifica del genoma (chiamata anche modifica del gene) è un gruppo di tecnologie che danno agli scienziati la capacità di modificare il DNA di un organismo. Queste tecnologie consentono di aggiungere, rimuovere o alterare materiale genetico in particolari punti del genoma. Sono stati sviluppati diversi approcci all’editing del genoma.
Uno recente è noto come CRISPR-Cas9, che è l’abbreviazione di brevi ripetizioni palindromiche raggruppate regolarmente interspaziate e proteina associata a CRISPR 9. Il sistema CRISPR-Cas9 ha generato molta eccitazione nella comunità scientifica perché è più veloce, più economico, più accurato e più efficiente di altri metodi di modifica del genoma esistenti.
CRISPR-Cas9 è stato adattato da un sistema di modifica del genoma presente in natura nei batteri. I batteri catturano frammenti di DNA dai virus invasori e li usano per creare segmenti di DNA noti come array CRISPR.
Gli array CRISPR consentono ai batteri di “ricordare” i virus (o quelli strettamente correlati). Se i virus attaccano di nuovo, i batteri producono segmenti di RNA dagli array CRISPR per colpire il DNA dei virus. I batteri quindi usano Cas9 o un enzima simile per tagliare il DNA, che disabilita il virus.
Il sistema CRISPR-Cas9 funziona in modo simile in laboratorio. I ricercatori creano un piccolo pezzo di RNA con una breve sequenza “guida” che si lega (si lega) a una specifica sequenza bersaglio di DNA in un genoma. L’RNA si lega anche all’enzima Cas9.
Come nei batteri, l’RNA modificato viene utilizzato per riconoscere la sequenza del DNA e l’enzima Cas9 taglia il DNA nella posizione mirata. Sebbene Cas9 sia l’enzima più utilizzato, è possibile utilizzare anche altri enzimi (ad esempio Cpf1).
Una volta tagliato il DNA, i ricercatori utilizzano il meccanismo di riparazione del DNA della cellula per aggiungere o eliminare pezzi di materiale genetico o per apportare modifiche al DNA sostituendo un segmento esistente con una sequenza di DNA personalizzata.
L’editing genomico è di grande interesse nella prevenzione e nel trattamento delle malattie umane. Attualmente, la maggior parte delle ricerche sull’editing del genoma viene svolta per comprendere le malattie utilizzando cellule e modelli animali. Gli scienziati stanno ancora lavorando per determinare se questo approccio è sicuro ed efficace per l’uso nelle persone.
Viene esplorato nella ricerca su un’ampia varietà di malattie, comprese le malattie monogeniche come la fibrosi cistica, l’emofilia e l’anemia falciforme. È anche promettente per il trattamento e la prevenzione di malattie più complesse, come cancro, malattie cardiache, malattie mentali e infezione da virus dell’immunodeficienza umana (HIV).
Le preoccupazioni etiche sorgono quando l’editing del genoma, utilizzando tecnologie come CRISPR-Cas9, viene utilizzato per alterare i genomi umani. La maggior parte delle modifiche introdotte con l’editing del genoma sono limitate alle cellule somatiche, che sono cellule diverse dalle cellule uovo e spermatozoi.
Questi cambiamenti interessano solo alcuni tessuti e non vengono trasmessi da una generazione all’altra. Tuttavia, le modifiche apportate ai geni nelle cellule uovo o spermatozoo (cellule germinali) o nei geni di un embrione potrebbero essere trasmesse alle generazioni future.
La modifica del genoma delle cellule germinali e dell’embrione solleva una serie di sfide etiche, inclusa la possibilità di utilizzare questa tecnologia per migliorare i normali tratti umani (come l’altezza o l’intelligenza). Sulla base di preoccupazioni circa l’etica e la sicurezza, l’editing del genoma delle cellule germinali e dell’embrione è attualmente illegale in molti paesi.
Modifica genica mediata da CRISPR / Cas9
Scoperte pionieristiche nella tecnologia CRISPR / Cas9
Il locus batterico CRISPR è stato descritto per la prima volta da Francisco Mojica (23) e successivamente identificato come un elemento chiave nel sistema immunitario adattativo dei procarioti (24). Il locus è costituito da frammenti di DNA virale o plasmidico che in precedenza infettavano il microbo (in seguito chiamati “spaziatori”), che sono stati trovati tra una serie di brevi sequenze ripetute palindromiche.
Successivamente, Alexander Bolotin scoprì la proteina Cas9 nello Streptococcus thermophilus, che a differenza di altri geni Cas noti, Cas9 era un gene di grandi dimensioni che codifica per una proteina a singolo effettore con attività nucleasica (25). Hanno inoltre notato una sequenza comune nel DNA bersaglio adiacente allo spaziatore, più tardi noto come motivo adiacente protospaziatore (PAM), la sequenza necessaria a Cas9 per riconoscere e legare il suo DNA bersaglio (25).
Studi successivi hanno riportato che gli spaziatori sono stati trascritti in CRISPR RNA (crRNA) che guidano le proteine Cas al sito bersaglio del DNA (26). Studi successivi hanno scoperto l’RNA CRISPR transattivante (tracrRNA), che forma un duplex con crRNA che insieme guidano Cas9 al suo DNA bersaglio (27).
Il potenziale utilizzo di questo sistema è stato semplificato introducendo un costrutto sintetico combinato di crRNA e tracrRNA chiamato RNA a guida singola (sgRNA) (28). Questo è stato seguito da studi che hanno dimostrato il successo dell’editing genomico da parte di CRISPR / Cas9 in cellule di mammifero, aprendo così la possibilità di implementare CRISPR / Cas9 nella terapia genica (29) (Figura 1).

Caratteristiche distintive della terapia genica CRISPR. Cronologia che evidenzia i principali eventi della terapia genica tradizionale, dello sviluppo di CRISPR e della terapia genica CRISPR. Il testo in rosso indica eventi di terapia genica che hanno sollevato significative preoccupazioni etiche.
Panoramica meccanicistica della modifica del genoma mediata da CRISPR / Cas9
CRISPR / Cas9 è un semplice sistema a due componenti utilizzato per un efficace editing genetico mirato. Il primo componente è la proteina Cas9 a effetto singolo, che contiene i domini endonucleasi RuvC e HNH.
RuvC scinde il filamento di DNA non complementare alla sequenza spaziatore e HNH scinde il filamento complementare. Insieme, questi domini generano rotture a doppio filamento (DSB) nel DNA bersaglio.
Il secondo componente di un efficace editing genetico mirato è un singolo RNA guida (sgRNA) che trasporta una sequenza di scaffold che consente il suo ancoraggio a Cas9 e una sequenza spaziatore di 20 coppie di basi complementare al gene target e adiacente alla sequenza PAM.
Questo sgRNA guida il complesso CRISPR / Cas9 alla sua posizione genomica prevista. Il sistema di editing si basa quindi su uno dei due percorsi di riparazione del DNA endogeno: non omologa end-join (NHEJ) o riparazione diretta dall’omologia (HDR) (Figura 2).
NHEJ si verifica molto più frequentemente nella maggior parte dei tipi di cellule e comporta l’inserimento e la cancellazione casuali di coppie di basi, o indel, nel sito di taglio.
Questo meccanismo soggetto a errori di solito si traduce in mutazioni frameshift, spesso creando un codone di stop prematuro e / o un polipeptide non funzionale. Questo percorso è stato particolarmente utile negli esperimenti di knock-out genetico e negli schermi CRISPR genomici funzionali, ma può anche essere utile in clinica nel contesto in cui la disgregazione genica fornisce un’opportunità terapeutica.
L’altro percorso, particolarmente interessante da sfruttare a fini clinici, è il percorso HDR privo di errori. Questo percorso prevede l’utilizzo della regione omologa del filamento di DNA non modificato come stampo per correggere il DNA danneggiato, con conseguente riparazione senza errori.
Sperimentalmente, questo percorso può essere sfruttato fornendo un modello di donatore esogeno con il meccanismo CRISPR / Cas9 per facilitare la modifica desiderata nel genoma (30).

Modifica genica mediata da CRISPR / Cas9. Cas9 in complesso con lo sgRNA prende di mira il rispettivo gene e crea DSB vicino alla regione PAM. La riparazione del danno al DNA procede attraverso il percorso NHEJ o HDR. Nel percorso NHEJ, inserimenti e delezioni casuali (indel) vengono introdotti sul lato tagliato e legati con conseguente riparazione soggetta a errori. Nel percorso HDR, il DNA cromosomico omologo funge da modello per il DNA danneggiato durante la riparazione, con conseguente riparazione senza errori.
Costruzione del modello sgRNA per l’editing genico di Cas9
Insieme alla nucleasi Cas9, gli esperimenti CRISPR richiedono l’introduzione di uno sgRNA contenente una sequenza di circa 20 basi specifica per il DNA bersaglio 5 ‘di una sequenza di scaffold non variabile. sgRNA può essere consegnato come RNA o trasformando con un plasmide con la sequenza codificante sgRNA sotto un promotore. Sono state sviluppate numerose strategie per sostituire rapidamente le 20 sequenze di base consentendo una comoda clonazione dello sgRNA utilizzando prodotti NEB.
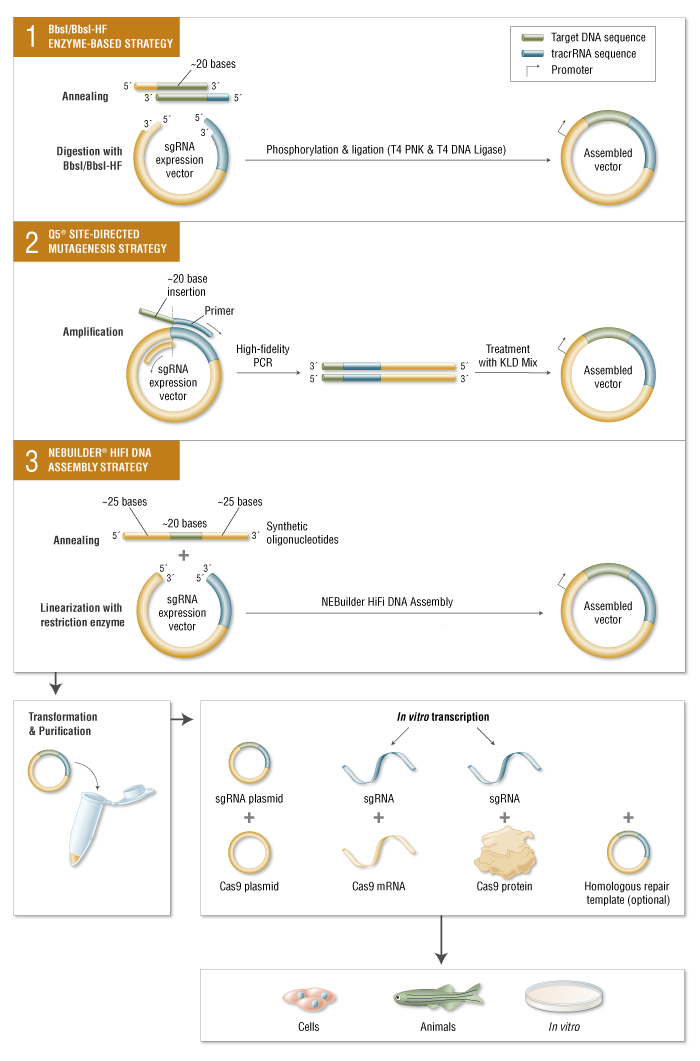
Limitazioni e avanzamenti di CRISPR / Cas9
Effetti fuori bersaglio
Una delle principali preoccupazioni per l’implementazione di CRISPR / Cas9 per la terapia genica è la frequenza relativamente elevata di effetti fuori bersaglio (OTE), che sono stati osservati con una frequenza ≥50% (31).
Gli attuali tentativi di affrontare questa preoccupazione includono varianti Cas9 ingegnerizzate che mostrano OTE ridotto e ottimizzazione del design delle guide.
Una strategia che riduce al minimo le OTE utilizza la nickasi Cas9 (Cas9n), una variante che induce rotture a filamento singolo (SSB), in combinazione con una coppia di sgRNA che mira a entrambi i filamenti del DNA nella posizione prevista per produrre il DSB (32).
I ricercatori hanno anche sviluppato varianti Cas9 che sono specificamente progettate per ridurre le OTE mantenendo l’efficacia dell’editing (Tabella 1).
SpCas9-HF1 è una di queste varianti ad alta fedeltà che sfrutta il modello di “eccesso di energia” che propone che vi sia un’affinità in eccesso tra Cas9 e il DNA bersaglio che potrebbe consentire le OTE. Introducendo mutazioni a 4 residui coinvolti nel legame idrogeno diretto tra Cas9 e la spina dorsale fosfato del DNA bersaglio, è stato dimostrato che SpCas9-HF1 non possiede attività fuori bersaglio rilevabile rispetto a SpCas9 di tipo selvatico (35).
Altre varianti Cas9 che sono state sviluppate includono evoCas9 e HiFiCas9, che contengono entrambi residui di amminoacidi alterati nel dominio Rec3 che è coinvolto nel riconoscimento dei nucleotidi. Desensibilizzare il dominio Rec3 aumenta la dipendenza dalla specificità dell’eteroduplex DNA: RNA per indurre i DSB, riducendo così le OTE pur mantenendo l’efficacia dell’editing (38, 39).
Uno degli sviluppi più recenti è la variante Cas9_R63A / Q768A, in cui la mutazione R63A destabilizza la formazione del loop R in presenza di mismatch e la mutazione Q768A aumenta la sensibilità ai mismatch PAM-distali (49).
Nonostante le diverse strategie, il razionale per generare molte varianti Cas9 con OTE ridotti è stato quello di ridurre in definitiva le interazioni Cas9 e DNA generali e dare un ruolo più forte al DNA: RNA eteroduplex nel facilitare le modifiche.
Tabella 1 – Varianti Cas9.
| Origine batterica | Variante Cas9 | Vantaggio | Mutazione variante | PAM | Riferimenti |
|---|---|---|---|---|---|
| Streptococcus pyogenes | Cas9-D1135E | Riconoscimento PAM migliorato | D1135E | NGG | (33) |
| Cas9-VQR | PAM alterato | D1135V / R1335Q / T1337R | NGAN o NGNG | ||
| Cas9-EQR | PAM alterato | D1135E / R1335Q / T1337R | NGAG | ||
| Cas9-Venom | PAM alterato | D1135V / G1218R / R1335E / T1337R | NGCG | ||
| Cas9-VRQR | PAM alterato | M495V / Y515N / K526E / R661Q | A PARTIRE DAL | ||
| Cas9-QQR1 | PAM alterato | G1218R / N1286Q / I1331F / D1332K / R1333Q / R1335Q / T1337R | NAAG | (34) | |
| SpCas9-HF1 | OTE ridotta | N497A / R661A / Q695A / Q926A | NGG | (35) | |
| eSpCas9 | OTE ridotta | K846A / K1003A / R1060A | NGG | (36) | |
| HeFSpCas9 | OTE ridotta | N497A / R661A / Q695A / K846A / Q926A / K1003A / R1060A | NGG | (37) | |
| evoCas9 | OTE ridotta | M495V / Y515N / K526E / R661Q | NGG | (38) | |
| HiFiCas9 | OTE ridotta | R691A | NGG | (39) | |
| Cas9n / Cas9D10A | SSB invece di DSB, OTE ridotta | D10A | NGG | (40, 41) | |
| Dimeric dCas9-FokI | OTE ridotta | dCas9 fuso al dominio endonucleasi FokI | NGG | (42) | |
| xCas9-3.7 | Ampia specificità PAM | A262T / R324L / S409I / E480K / E543D / M694I / E1219V | NG, GAA o GAT | (43) | |
| SpCas9-NG | PAM minimo | R1335V / L1111R / D1135V / G1218R / E1219F / A1322R / T1337R | NGN | (44) | |
| HypaCas9 | OTE ridotta | N692A / M694A / Q695A / H698A | NGG | (45) | |
| Sniper-Cas9 | OTE ridotta | F539S / M763I / K890N | NGG | (46) | |
| SpG Cas9 | PAM minimo | D1135L / S1136W / G1218K / E1219Q / R1335Q / T1337R | NGN | (47) | |
| SpRY Cas9 | PAM minimo | D1135L / S1136W / G1218K / E1219Q / R1335Q / T1337R / L1111R / A1322R / A61R / N1317R / R1333P | RRN> NYN | ||
| SpCas9-HF1 più | OTE ridotta | N497A / Q695A / Q926A; amminoacidi 1005-1013 sostituiti con due glicina | NGG | (48) | |
| eSpCas9 altro | OTE ridotta | K848A / R1060A; amminoacidi 1005-1013 sostituiti con due glicina | NGG | ||
| Cas9_R63A / Q768A | OTE ridotta | R63A / Q768A | NGG | (49) | |
| Staphylococcus aureus | KKH SaCas9 | PAM rilassato | E782K / N968K / R1015H | NNNRRT | (33) |
| SaCas9-HF | OTE ridotta | R245A / N413A / N419A / R654A | NNGRRT | (50) | |
| SaCas9-NR | PAM rilassato | N986R | NNGRR | (51) | |
| SaCas9-RL | PAM rilassato | N986R / R991L | NNGRR | ||
| Streptococcus canis | ScCas9 | PAM minimo | N / A (tipo jolly) |
L’ottimizzazione della progettazione delle guide può anche ridurre la frequenza delle OTE (31). Molte caratteristiche in uno sgRNA determinano la specificità inclusa la sequenza seme (una regione di 10-12 bp prossimale a PAM su 3 ‘della sequenza spaziatore) (29, 53), il contenuto GC (54, 55) e modifiche come il troncamento di 5’ lo sgRNA (56).
Sono state progettate anche diverse piattaforme per fornire sequenze guida ottimizzate contro i geni bersaglio, tra cui E-Crisp (31, 57), CRISPR-design, CasOFFinder e altri (31). Tuttavia, molti di questi strumenti sono progettati sulla base di algoritmi computazionali con parametri variabili o si basano su schermi fenotipici che possono essere specifici per tipi di cellule e genomi, generando rumore apprezzabile e mancanza di generalizzabilità attraverso diverse configurazioni sperimentali (58, 59).
Recentemente, è stato sviluppato un ulteriore strumento per la progettazione di guide denominato sgDesigner che ha affrontato queste limitazioni utilizzando una nuova libreria di plasmidi in silico che conteneva sia lo sgRNA che il sito target all’interno dello stesso costrutto.
Ciò ha consentito di raccogliere i dati sull’efficienza di modifica di Cas9 in modo intrinseco e di stabilire un nuovo set di dati di addestramento che evita i pregiudizi introdotti da altri modelli. Inoltre, una valutazione comparativa delle prestazioni per prevedere l’efficienza dello sgRNA di sgDesigner con altri 3 strumenti comunemente usati (Doench Rule Set 2, Sequence Scan for CRISPR e DeepCRISPR) ha rivelato che sgDesigner ha superato tutti e 3 gli strumenti di designer in 6 set di dati indipendenti, suggerendo che sgDesigner potrebbe essere un piattaforma più robusta e generalizzabile (60).
Requisiti del motivo adiacente Protospacer
Un’ulteriore limitazione della tecnologia è il requisito per un PAM vicino al sito di destinazione. Cas9 del batterio Streptococcus pyogenes (SpCas9) è uno dei Cas9 più ampiamente utilizzati con un sito di riconoscimento PAM canonico relativamente breve: 5′NGG3 ′, dove N è qualsiasi nucleotide.
Tuttavia, SpCas9 è relativamente grande e difficile da impacchettare in vettori AAV (61, 62), il veicolo di rilascio più comune per la terapia genica.
Staphylococcus aureus Cas9 (SaCas9) è un ortologo più piccolo che può essere impacchettato più facilmente in vettori AAV ma ha una sequenza PAM più lunga: 5′NNGRRT3 ′ o 5′NNGRR (N) 3 ′, dove R è qualsiasi purina, che restringe ulteriormente il finestra di siti di targeting terapeutico.
Sono state realizzate varianti SaCas9 ingegnerizzate, come KKH SaCas9, che riconosce un PAM 5′NNNRRT3 ′, ampliando i siti di targeting umano da 2 a 4 volte. Le OTE, tuttavia, sono osservate con frequenze simili a SaCas9 di tipo selvatico e devono essere considerate nella progettazione di qualsiasi applicazione terapeutica (33).
Diverse altre varianti di SpCas9 sono state progettate per ampliare la finestra del gene target tra cui SpCas9-NG, che riconosce un NG PAM minimo (44) e xCas9, che riconosce un’ampia gamma di PAM tra cui NG, GAA e GAT (43). Un confronto fianco a fianco di entrambe le varianti ha rivelato che mentre SpCas9-NG aveva un riconoscimento PAM più ampio, xCas9 aveva l’OTE più bassa nelle cellule umane (63).
Un altro ortholog Cas9 del batterio Streptococcus canis, ScCas9, è stato recentemente caratterizzato con una specificità PAM minima di 5′NNG3 ′ e un’omologia di sequenza dell’89,2% a SpCas9 ed efficienza di editing comparabile a SpCas9 sia nelle cellule batteriche che in quelle umane (52).
Lo sviluppo più recente è una variante di SpCas9 denominata SpRY che è stata progettata per essere quasi priva di PAM, riconoscendo PAM NRN> NYN minimi. Questa nuova variante può potenzialmente modificare qualsiasi gene indipendente da un requisito di PAM e quindi può essere utilizzata terapeuticamente contro diverse malattie genetiche (47).
In alternativa, sono state sviluppate varianti Cas9 mirate all’RNA che ampliano anche lo spettro del targeting genetico mitigando le restrizioni dei requisiti di PAM. S. pyogenese Cas9 (SpyCas9) può essere manipolato per mirare all’RNA fornendo un breve oligonucleotide con una sequenza PAM, nota come PAMmer (64, 65), eliminando così la necessità di un sito PAM all’interno della regione bersaglio.
Sono stati scoperti anche altri sottoinsiemi di enzimi Cas che mirano naturalmente all’RNA indipendentemente da un PAM, come Cas13d. Dopo l’ulteriore progettazione di questo effettore, CasRx è stato sviluppato per un efficiente targeting dell’RNA guidato da RNA nelle cellule umane (66, 67). Sebbene i progressi del CRISPR mirati all’RNA forniscano un’opportunità terapeutica senza il rischio di tossicità da danno al DNA, escludono la possibilità di modificare una correzione permanente nel genoma.
Tossicità per danni al DNA
I DSB indotti da CRISPR spesso innescano l’apoptosi piuttosto che la modifica genetica prevista (68). Ulteriori problemi di sicurezza sono stati rivelati quando si utilizza questo strumento nelle cellule staminali pluripotenti umane (hPSC) che hanno dimostrato che l’attivazione di p53 in risposta ai DSB tossici introdotti da CRISPR spesso innesca la successiva apoptosi (69).
Pertanto, è più probabile che le modifiche CRISPR di successo si verifichino nelle cellule soppresse con p53, determinando un bias verso la selezione per la sopravvivenza delle cellule oncogene (70). Inoltre, in diversi casi sono state riportate ampie delezioni che abbracciano kilobasi e riarrangiamenti complessi come conseguenze non intenzionali dell’attività sul bersaglio (71, 72), evidenziando un importante problema di sicurezza per le applicazioni cliniche della terapia CRISPR che induce DSB.
Altre variazioni di Cas9, come l’endonucleasi morta cataliticamente inattiva Cas9 (dCas9) in cui i domini nucleasi sono disattivati, possono fornire utilità terapeutica mitigando i rischi dei DSB (73). dCas9 può manipolare transitoriamente l’espressione di geni specifici senza introdurre DSB attraverso la fusione di domini o proteine di attivazione o repressione trascrizionale nell’effettore di legame al DNA (74).
Si possono anche considerare altre varianti come Cas9n, che induce SSB piuttosto che DSB. Ulteriori modifiche di queste varianti Cas9 hanno portato allo sviluppo di editor di base e editor principali, un’innovazione chiave per un’applicazione terapeutica sicura della tecnologia CRISPR (vedere la sezione Editing genetico di precisione con CRISPR).
Immunotossicità
Oltre ai limiti tecnici, CRISPR / Cas9, come la terapia genica tradizionale, solleva ancora preoccupazioni per la tossicità immunogenica.
Charlesworth et al. hanno mostrato che più della metà dei soggetti umani nel loro studio possedeva anticorpi anti-Cas9 preesistenti contro gli ortologhi batterici più comunemente usati, SaCas9 e SpCas9 (75).
Inoltre, i vettori AAV sono anche ampiamente utilizzati per fornire componenti CRISPR per la terapia genica. A tal fine, diversi ortologhi Cas9 e sierotipi AAV sono stati testati sulla base di somiglianze di sequenza e forza di legame prevista per MHC di classe I e II per lo screening di ortologhi immunitari che possono essere utilizzati per la somministrazione ripetuta sicura della terapia genica AAV-CRISPR.
Sebbene non siano stati trovati due sierotipi di AAV in grado di aggirare completamente il riconoscimento immunitario, lo studio ha verificato 3 ortologi Cas9 [SpCas9, SaCas9 e Campylobacter jejuni Cas9 (CjCas9)] che hanno mostrato una robusta efficienza di editing e tollerato la somministrazione ripetuta a causa della ridotta tossicità immunogenica nei topi AAV e Cas9 (76).
Un avvertimento importante è l’immunità preesistente negli esseri umani contro 2 di questi ortologhi: SpCas9 e SaCas9, lasciando CjCas9 come l’unica opzione corrente per questa coorte di pazienti. Tuttavia, questo ortologo non è stato ben studiato rispetto agli altri 2 ortologi e necessiteranno di ulteriori indagini per fornire prove della sua sicurezza ed efficacia per l’uso clinico. Studi futuri potrebbero anche identificare altri ortologhi immunoortogonali Cas9 per una terapia genica ripetuta sicura.
Editing genetico di precisione con CRISPR
L’editing preciso del genoma è essenziale per le prospettive di terapia genica CRISPR. Sebbene i percorsi HDR possano facilitare una modifica desiderata, la sua bassa efficienza rende la sua utilità per l’editing genetico preciso per l’intervento clinico altamente limitante, con NHEJ come percorso predefinito che le cellule umane prendono per la riparazione.
Il miglioramento dell’efficienza dell’HDR è stato ottenuto tramite la soppressione del percorso NHEJ attraverso l’inibizione chimica degli enzimi chiave che modulano l’NHEJ come Ku (77), DNA ligasi IV (78) e proteine chinasi DNA-dipendenti (DNA-PKcs) (79). Altre strategie che migliorano l’efficienza HDR includono l’utilizzo di un modello di oligodesossinucleotide a filamento singolo (ssODN), che contiene i bracci di omologia per facilitare la ricombinazione e la sequenza di modifica desiderata, invece del DNA a doppio filamento (dsDNA).
È stato dimostrato che modelli ssODN progettati in modo razionale con complementarità di lunghezza ottimizzata aumentano i tassi di HDR fino al 60% nelle cellule umane per la sostituzione di un singolo nucleotide (80). Inoltre, la fase del ciclo cellulare gioca un ruolo chiave nel determinare il percorso di riparazione del danno al DNA che una cellula può seguire. Gli eventi HDR sono generalmente limitati alle fasi S e G2 tardive del ciclo cellulare, data la disponibilità del cromatide fratello a fungere da modello in queste fasi, mentre NHEJ predomina nelle fasi G1, S e G2 (81).
L’arresto farmacologico nella fase S con afidicolina ha aumentato la frequenza HDR in HEK293T con consegna della ribonucleoproteina guida Cas9 (RNP). È interessante notare che l’arresto cellulare nella fase M utilizzando nocodazolo con basse concentrazioni del complesso RNP guida Cas9 ha prodotto frequenze più elevate di eventi HDR in queste cellule, raggiungendo una frequenza massima fino al 31% (82).
Sebbene l’HDR sia considerato limitato alle cellule mitotiche, uno studio recente ha rivelato che l’editing CRISPR / Cas9 può raggiungere l’HDR nei neuroni postmitotici maturi. Nishiyama et al. modificato con successo il locus CaMKIIα tramite HDR in neuroni ippocampali postmitotici di topi adulti in vitro utilizzando un Cas9 consegnato da AAV, RNA guida e modello di donatore nel locus CaMKIIα, che ha ottenuto modifiche mediate da HDR di successo in circa il 30% delle cellule infette.
Sebbene l’efficienza HDR fosse dose-dipendente dal macchinario HDR fornito dall’AAV e l’attività fuori bersaglio non fosse monitorata, questo studio ha dimostrato la potenziale utilità di CRISPR per le neuroscienze traslazionali dopo ulteriori sviluppi (83). Per sfruttare ulteriormente il controllo dello stadio del ciclo cellulare come mezzo per favorire la riparazione basata su modelli, la coniugazione di Cas9 con una parte di Geminin, un substrato per la degradazione del proteosoma G1, può limitare l’espressione di Cas9 agli stadi S, G2 e M.
È stato dimostrato che questa strategia facilita gli eventi HDR mitigando le modifiche NHEJ indesiderate nelle cellule staminali e immortalate umane (84, 85). Una strategia più recente ha combinato un Cas9 modificato chimicamente al donatore ssODN o un adattatore di DNA che recluta il modello del donatore, entrambi i quali hanno migliorato l’efficienza HDR localizzando il modello del donatore vicino al sito di scissione (86).
Nonostante questi progressi, l’HDR è ancora raggiunto con un’efficienza relativamente bassa nelle cellule eucariotiche e l’uso di agenti relativamente dannosi nelle cellule come gli inibitori chimici NHEJ potrebbe non essere l’ideale in un contesto clinico.
Un recente progresso che consente l’editing genetico di precisione indipendentemente dallo sfruttamento dei meccanismi di risposta al danno del DNA è il sistema di editing di base CRISPR (BE). In questo sistema, un Cas9 morto cataliticamente inattivo (dCas9) è coniugato alla deaminasi, che può catalizzare la conversione dei nucleotidi tramite deaminazione.
Per una maggiore efficienza di editing, la nickasi Cas9 (Cas9n) fusa con la deaminasi è stata recentemente utilizzata di più su dCas9 per l’editing di base, poiché le tacche create in un singolo filamento di DNA inducono una maggiore efficienza di editing. Attualmente, i due tipi di editor di base CRISPR sono gli editor di base della citidina (CBE) e gli editor di base dell’adenosina (ABE).
I CBE catalizzano la conversione della citidina in uridina, che diventa timina dopo la replicazione del DNA. Gli ABE catalizzano la conversione dell’adenosina in inosina che diventa guanina dopo la replicazione del DNA (87). Gli editor di base forniscono un mezzo per modificare singoli nucleotidi senza correre il rischio di causare tossicità indotta da DSB.
Tuttavia, gli editor di base sono limitati alle conversioni da “A a T” e da “C a G”, restringendo il campo di applicazione dell’edizione di geni a base singola solo a queste basi. Inoltre, gli editor di base devono ancora affrontare alcune delle stesse sfide dei sistemi CRISPR descritti in precedenza, comprese le OTE, più con i CBE che con gli ABE (88, 89) e i vincoli di imballaggio, in particolare nei vettori AAV a causa delle grandi dimensioni degli editor di base ( 90).
Inoltre, la finestra di modifica per gli editor di base è limitata a una gamma ristretta di poche basi a monte del PAM (90). Più recentemente, il prime editing è stato sviluppato come strategia per modificare il genoma per inserire un tratto di modifiche desiderato senza indurre DSB (91).
Questa tecnologia combina la fusione di Cas9n con una trascrittasi inversa e una guida di editing principale RNA (pegRNA), che contiene la sequenza di sgRNA, il sito di legame del primer (PBS) e un modello di RNA che codifica la modifica desiderata all’estremità 3 ‘. I redattori principali usano Cas9n per intaccare un filamento del DNA e inserire la modifica desiderata tramite la trascrizione inversa del modello di RNA.
La modifica sintetizzata viene incorporata nel genoma e il filamento non modificato viene tagliato e riparato per corrispondere alla modifica inserita. Con un sistema di consegna ottimizzato in atto, gli editor di base e gli editor di primer possono aprire la porta all’editing genetico di precisione per correggere e potenzialmente curare una moltitudine di malattie genetiche (Figura 3).

Modifica genetica precisa. (A) CRISPR / Cas9-HDR. Cas9 induce un DSB. Il ssODN esogeno che trasporta la sequenza per i bracci di modifica e omologia desiderati viene utilizzato come modello per la modifica genica mediata da HDR. (B) Editor di base. dCas9 o Cas9n è legato alla porzione catalitica di una deaminasi. La citosina deaminasi catalizza la formazione di uridina dalla citosina. I meccanismi di riparazione del disadattamento del DNA o la replicazione del DNA producono una modifica da C: G a T: una singola base nucleotidica. L’adenosina deaminasi catalizza la formazione di inosina dall’adenosina. I meccanismi di riparazione del disadattamento del DNA o la replicazione del DNA producono una modifica di base a singolo nucleotide da A: T a G: C. (C) Prime Editor. Cas9n è legato alla porzione catalitica della trascrittasi inversa. Il sistema prime editor utilizza pegRNA, che contiene la sequenza del distanziatore guida, il primer della trascrittasi inversa, che include la sequenza per la modifica desiderata e un sito di legame del primer (PBS). PBS si ibrida con la regione complementare del DNA e la trascrittasi inversa trascrive il nuovo DNA portando la modifica desiderata. Dopo la scissione del lembo 5 ‘risultante e la legatura, i meccanismi di riparazione del DNA correggono il filo non modificato in modo che corrisponda al filo modificato. HDR, riparazione diretta dall’omologia. DSB, rottura a doppio filamento; SSB, rottura a filamento singolo; ssODN, oligodesossinucleotide a filamento singolo. rottura a filamento singolo; ssODN, oligodesossinucleotide a filamento singolo. rottura a filamento singolo; ssODN, oligodesossinucleotide a filamento singolo.
Erogazione della terapia genica CRISPR
La modalità di consegna degli strumenti CRISPR influenza notevolmente la sua sicurezza e l’efficacia terapeutica. Mentre la terapia genica tradizionale che utilizza virus è stata esaminata per il rischio di immunotossicità e oncogenesi inserzionale, i vettori AAV rimangono un veicolo di consegna chiave per la terapia genica CRISPR e continuano ad essere ampiamente utilizzati per la sua alta efficienza di consegna (92).
Il toolkit CRISPR può essere confezionato come DNA plasmidico che codifica i suoi componenti, inclusi Cas9 e gRNA, oppure può essere consegnato come mRNA di Cas9 e gRNA. Gli acidi nucleici di CRISPR possono essere confezionati in vettori AAV per la consegna o introdotti nelle cellule bersaglio tramite elettroporazione / nucleofection o microiniezione, con questi ultimi metodi che evitano i rischi associati ai virus.
Tuttavia, la microiniezione può essere tecnicamente impegnativa ed è adatta solo per la consegna ex vivo. Anche l’elettroporazione è ampiamente utilizzata per ex vivo, ma può essere utilizzata in vivo per alcuni tessuti bersaglio (93). Tuttavia, lo shock ad alta tensione necessario per permeabilizzare le membrane cellulari tramite l’elettroporazione può essere tossico e può portare alla permeabilizzazione permanente delle cellule trattate (94).
Oltre alla tossicità virale, la somministrazione AAV dei componenti CRISPR produce longevità di espressione, portando a una maggiore incidenza di OTE. In alternativa, il rilascio della proteina Cas9 e del gRNA come complessi RNP ha ridotto le OTE pur mantenendo l’efficacia dell’editing, grazie alla sua espressione transitoria e alla rapida clearance nella cellula (95).
Una volta selezionata la modalità di consegna, le modifiche CRISPR / Cas9 possono essere agevolate ex vivo, dove le cellule vengono modificate geneticamente al di fuori del paziente e reintrodotte, o in vivo con la consegna dei componenti CRISPR direttamente nel paziente dove le cellule vengono modificate (Figura 4 ). Entrambi i sistemi pongono la propria serie di vantaggi e sfide.
I vantaggi per la somministrazione ex vivo includono una maggiore sicurezza poiché i pazienti non sono esposti allo strumento di alterazione genica, fattibilità tecnica e controllo di qualità più stretto delle cellule modificate. Tuttavia, le sfide a questo metodo includono la sopravvivenza e la conservazione della funzione in vivo delle cellule al di fuori del paziente dopo la manipolazione genetica e un’ampia coltura in vitro. Inoltre, è necessaria una fornitura adeguata di cellule per un reinnesto efficiente. Queste condizioni limitano questo metodo a certi tipi di cellule che possono sopravvivere ed essere espanse in coltura, come le cellule staminali e progenitrici ematopoietiche (HSPC) (96) e le cellule T (97).

Erogazione della terapia CRISPR. Gli acidi nucleici che codificano CRISPR / Cas9 o il suo complesso RNP possono essere imballati in veicoli di consegna. Una volta confezionati, le modifiche possono essere agevolate ex vivo o in vivo. L’editing ex vivo prevede l’estrazione di cellule bersaglio dal paziente, la coltura cellulare e l’espansione in vitro, la consegna dei componenti CRISPR per produrre le modifiche desiderate, la selezione e l’espansione delle cellule modificate e infine la reintroduzione delle cellule modificate terapeutiche nel paziente. L’editing in vivo può essere somministrato sistemicamente tramite infusioni endovenose al paziente, dove il carico CRISPR viaggia attraverso il flusso sanguigno attraverso le arterie che portano al tessuto bersaglio, o somministrato localmente con iniezioni direttamente al tessuto bersaglio. Una volta consegnate, le modifiche vengono agevolate in vivo per fornire benefici terapeutici.
Sebbene la terapia genica ex vivo abbia fornito benefici terapeutici per i disturbi ematologici e l’immunoterapia del cancro, molti tipi di tessuto non sono adatti per questo metodo, limitando gravemente la sua utilità terapeutica per altre malattie genetiche. La manipolazione in vivo è quindi necessaria per espandere l’utilità del CRISPR nel trattamento di una gamma più ampia di malattie genetiche, come la distrofia muscolare di Duchenne (DMD) (98) e la tirosinemia ereditaria (99).
I componenti di CRISPR possono essere somministrati in vivo per via sistemica tramite iniezioni endovenose o possono essere iniettati localmente a tessuti specifici (Figura 4). Con la somministrazione sistemica, i componenti CRISPR e il suo veicolo vengono introdotti nel sistema circolatorio dove l’espressione del kit di strumenti di modifica genica può essere controllata per mirare a organi specifici tramite promotori tessuto-specifici (100). Tuttavia, le sfide della consegna in vivo includono la degradazione da parte di proteasi o nucleasi circolanti, opsonizzazione da opsonine o eliminazione da parte del sistema dei fagociti mononucleari (MPS).
Inoltre, il carico deve raggiungere il tessuto bersaglio e bypassare l’endotelio vascolare, che spesso sono strettamente collegati da giunzioni cellula-cellula (101), impedendo l’accessibilità a veicoli di consegna più grandi (diametro> 1 nm). Inoltre, una volta che il carico ha raggiunto le cellule bersaglio, queste devono essere interiorizzate, il che è generalmente facilitato dall’endocitosi dove possono essere trasportate e degradate dagli enzimi lisosomiali (102).
Inoltre, la localizzazione del macchinario di editing vicino al punto di iniezione può comportare una distribuzione non uniforme del repertorio cellulare modificato all’interno del tessuto, che può portare a risultati terapeutici non ottimali (102). Mentre i progressi continuano a perfezionare le tecniche di somministrazione, i sistemi attuali hanno consentito di utilizzare la terapia genica CRISPR in clinica.
Modifica CRISPR in embrioni umani e considerazioni etiche
Sebbene l’editing somatico per la terapia CRISPR sia stato consentito dopo un’attenta considerazione, l’editing della linea germinale umana per intenti terapeutici rimane molto controverso. Con l’edizione somatica, qualsiasi rischio potenziale sarebbe contenuto all’interno dell’individuo dopo il consenso informato a partecipare alla terapia. L’editing embrionale non solo rimuove l’autonomia nel processo decisionale degli individui nati più tardi, ma consente anche che effetti collaterali imprevisti e permanenti si trasmettano di generazione in generazione.
Questo stesso potere richiede di procedere con cautela per prevenire gravi battute d’arresto come testimoniato dalla terapia genica tradizionale. Tuttavia, un controverso processo CRISPR su embrioni umani condotto da Jiankui He potrebbe aver già violato gli standard etici stabiliti per tali studi.
Questo studio pilota ha coinvolto l’ingegneria genetica del gene CCR5 (CCR5) del recettore per le chemochine negli embrioni umani, con l’intenzione di conferire resistenza all’HIV, come si è visto da una mutazione CCR5Δ32 presente in natura in alcuni individui (108). Tuttavia, sulla base delle prove limitate, CRISPR / Cas9 è stato probabilmente utilizzato per indirizzare questo gene, ma piuttosto che replicare la delezione a 32 basi naturalmente osservata e benefica, le modifiche hanno semplicemente indotto DSB a un’estremità della delezione, consentendo a NHEJ di riparare il danneggiato DNA introducendo mutazioni casuali e non caratterizzate.
Pertanto, non è noto se la proteina risultante funzionerà in modo simile alla proteina CCR5Δ32 presente in natura e conferirà resistenza all’HIV. Inoltre, solo uno dei due embrioni, denominato con lo pseudonimo di Nana, ha avuto modifiche riuscite in entrambe le copie del gene CCR5, mentre l’altro embrione, con lo pseudonimo di Lulu, ha avuto un editing riuscito in una sola copia. Nonostante questi risultati, entrambi gli embrioni sono stati reimpiantati nella madre, sapendo che la resistenza all’HIV sarà discutibile a Nana e inesistente a Lulu (109, 110).
Inoltre, studi recenti hanno dimostrato che il meccanismo di infezione di alcune varianti del virus HIV altamente mutabile può fare affidamento sul co-recettore del recettore CXC di tipo 4 (CXCR4) per le chemochine (108, 111). Senza tentativi di modificare CXCR4, questo aggiunge un ulteriore livello di scetticismo verso il raggiungimento della resistenza all’HIV con questa strategia. Inoltre, le OTE, in particolare durante la vita di un individuo, rimangono una delle principali preoccupazioni per l’applicazione di questa tecnologia negli esseri umani.
I recenti progressi nello strumento di modifica per limitare le OTE, come l’utilizzo di varianti Cas9 ad alta fedeltà, non sono stati sfruttati. Inoltre, la logica alla base della selezione della prevenzione dell’HIV per il primo utilizzo di CRISPR in embrioni umani impiantati contribuisce allo scarso rapporto rischio / beneficio di questo studio, considerando che i pazienti HIV possono vivere una vita lunga e sana con un regime farmacologico.
Un primo tentativo più appropriato sarebbe stato quello di impiegare questa tecnologia per una malattia più grave. Ad esempio, la correzione del gene MYBPC3 è probabilmente un bersaglio migliore per l’editing genico embrionale, poiché le mutazioni in MYBPC3 possono causare cardiomiopatia ipertrofica (HCM), una condizione cardiaca responsabile della maggior parte delle morti cardiache improvvise nelle persone di età inferiore ai 30 anni.
La correzione genica per questa mutazione patologica è stata ottenuta recentemente per la prima volta negli Stati Uniti in embrioni umani vitali utilizzando il sistema CRISPR / Cas9 mediato da HDR. Tuttavia, questi embrioni sono stati modificati per scopi di ricerca di base e non destinati all’impianto. In questo studio, gli spermatozoi portatori della mutazione patogena MYBPC3 e il meccanismo CRISPR / Cas9 come complesso RNP sono stati microiniettati in ovociti donatori sani arrestati al MII, ottenendo il 72,4% di embrioni omozigoti di tipo selvatico rispetto al 47,4% di embrioni non trattati.
La correzione genica mediata da HDR è stata osservata a frequenze considerevolmente elevate senza OTE rilevabili in blastomeri selezionati, probabilmente a causa del rilascio diretto di microiniezione del complesso RNP nello zigote iniziale. È interessante notare che il DNA di tipo selvatico materno è stato utilizzato preferenzialmente per la riparazione basata su modelli rispetto al modello ssODN esogeno fornito (112).
Sebbene le prove per la correzione genica fossero promettenti, la riparazione del DNA mediata dall’NHEJ è stata ancora osservata in molti embrioni, evidenziando la necessità di migliorare l’efficienza dell’HDR prima di poter prendere in considerazione l’applicazione clinica. Sebbene siano state sviluppate strategie per migliorare l’HDR, come gli inibitori chimici dell’NHEJ (77-79), tali tecniche possono avere risultati variabili nelle cellule embrionali e gli effetti collaterali che possono derivare dal trattamento devono essere studiati. L’editing genico della linea germinale rimarrà eticamente sfavorevole al suo stato attuale e le sue discussioni potrebbero non essere prese in considerazione fino a quando non saranno valutati sufficienti studi a lungo termine degli studi clinici sulla terapia somatica CRISPR in corso.
Potenziale per la terapia CRISPR durante la pandemia di COVID-19
La tecnologia CRISPR in rapida evoluzione può fornire aiuto durante i nostri tempi in rapida evoluzione. La recente epidemia di coronavirus 2 (SARS-CoV-2) di una nuova sindrome respiratoria acuta grave ha portato a una pandemia globale (113). Questi tempi pressanti richiedono una risposta urgente per sviluppare strumenti di test rapidi ed efficienti e opzioni di trattamento per i pazienti con malattia da coronavirus 2019 (COVID-19).
I metodi attualmente disponibili per i test richiedono tempo relativamente con accuratezza e sensibilità non ottimali (114). I due metodi di analisi predominanti sono i test molecolari o i test sierologici. I Centers for Disease Control and Prevention (CDC) degli Stati Uniti hanno sviluppato un test RT-PCR in tempo reale per test molecolari per la presenza di RNA virale per rilevare COVID-19 (115). Tuttavia, questo test ha una percentuale di falsi negativi di circa il 30% circa (116, 117) con un tempo di risposta da diverse ore a> 24 ore. I metodi di analisi sierologici sono molto più rapidi ma non sono in grado di rilevare l’infezione respiratoria acuta poiché gli anticorpi utilizzati per rilevare l’infezione possono richiedere diversi giorni o settimane per svilupparsi.
Recentemente, un test basato su CRISPR Cas12 denominato SARS-CoV-2 DETECTR è stato sviluppato per il rilevamento di COVID-19 con un breve tempo di risposta di circa 40 min e una precisione riportata del 95%. Il dosaggio prevede l’estrazione dell’RNA seguita dalla trascrizione inversa e dall’amplificazione isotermica simultanea utilizzando il metodo RT-LAMP. Vengono quindi presi di mira Cas12 e un RNA guida contro le regioni del gene N (nucleoproteina) e del gene E (involucro) di SARS-CoV-2, che possono essere visualizzati mediante scissione di una molecola reporter fluorescente.
Il test include anche una striscia a flusso laminare per una lettura visiva, dove una singola banda vicino a dove è stato applicato il campione indica un test negativo e 2 bande più alte o una singola banda più alta indicherebbero la scissione della sonda fluorescente e quindi positiva per SARS- CoV-2 (118).
Oltre all’utilità diagnostica di CRISPR, CRISPR può fornire opzioni terapeutiche per i pazienti COVID-19. Il Cas13 recentemente scoperto è un’endonucleasi RNA guidata da RNA che può servire come potenziale strumento terapeutico contro COVID-19. È stato sviluppato PAC-MAN (Prophylactic Antiviral CRISPR in huMAN cells), che utilizza la variante VI-D CRISPR-Cas13d derivata da Ruminococcus flavefaciens, selezionata per le sue piccole dimensioni che facilita il confezionamento più facile nei veicoli virali, alta specificità e forte attività catalitica nell’uomo cellule.
Questa tecnica è stata sviluppata per colpire simultaneamente più regioni per la degradazione dell’RNA, aprendo la porta a una strategia di targeting del pan-coronavirus tanto necessaria, date le prove che suggeriscono tassi di mutazione e ricombinazione relativamente alti di SARS-CoV-2 (119). Con questi progressi, il meccanismo CRISPR / Cas può essere nuovamente implementato per servire il suo scopo originale come sistema di lotta contro i virus per fornire aiuto durante questa pandemia.
link di riferimento: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7427626/
Informazioni : Cell



{kind=link}