










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A simple eye exam combined with powerful artificial intelligence (AI) machine learning technology could provide early detection of Parkinson’s disease, according to research being presented at the annual meeting of the Radiological Society of North America (RSNA).
Parkinson’s disease is a progressive disorder of the central nervous system that affects millions of people worldwide.
Diagnosis is typically based on symptoms like tremors, muscle stiffness and impaired balance – an approach that has significant limitations.
“The issue with that method is that patients usually develop symptoms only after prolonged progression with significant injury to dopamine brain neurons,” said study lead author Maximillian Diaz, a biomedical engineering Ph.D. student at the University of Florida in Gainesville, Florida.
“This means that we are diagnosing patients late in the disease process.”
Disease progression is characterized by nerve cell decay that thins the walls of the retina, the layer of tissue that lines the back of the eyeball.
The disease also affects the microscopic blood vessels, or microvasculature, of the retina. These characteristics present an opportunity to leverage the power of AI to examine images of the eyes for signs of Parkinson’s disease.
For the new study, Diaz collaborated with graduate student Jianqiao Tian and University of Florida neurologist Adolfo Ramirez-Zamora, M.D., under the direction of Ruogu Fang, Ph.D., director of the J. Crayton Pruitt Department of Biomedical Engineering’s Smart Medical Informatics Learning and Evaluation Lab (SMILE).
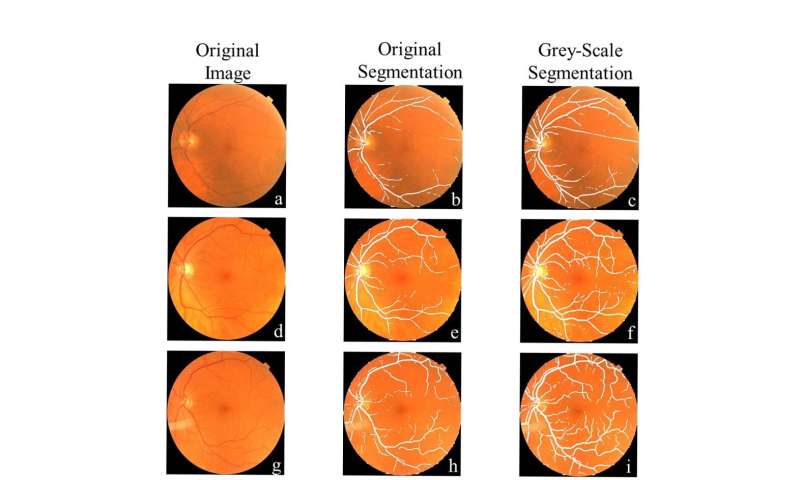
The researchers deployed a type of AI called support vector machine (SVM) learning that has been around since 1989. Using pictures of the back of the eye from both patients with Parkinson’s disease and control participants, they trained the SVM to detect signs on the images suggestive of disease.
The results indicated that the machine learning networks can classify Parkinson’s disease based on retina vasculature, with the key features being smaller blood vessels. The proposed methods further support the idea that changes in brain physiology can be observed in the eye.
“The single most important finding of this study was that a brain disease was diagnosed with a basic picture of the eye,” Diaz said. “This is very different from traditional approaches where to find a problem with the brain you look at different brain images.”
Diaz noted that those traditional imaging approaches with MRI, CT and nuclear medicine techniques can be very costly.
In contrast, the new approach uses basic photography with equipment commonly available in eye clinics to get an image. The images can even be captured by a smartphone with a special lens.
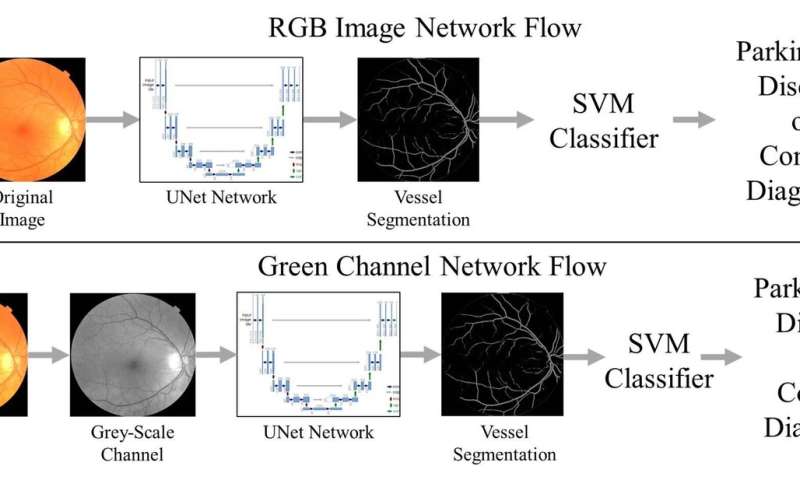
“It’s just a simple picture of the eye, you can have it done in less than a minute, and the cost of the equipment is much less than a CT or MRI machine,” Diaz said.
“If we can make this a yearly screening, then the hope is that we can catch more cases sooner, which can help us better understand the disease and find a cure or a way to slow the progression.”
The approach may also have applications in identifying other diseases that affect the structure of the brain, such as Alzheimer’s disease and multiple sclerosis, Diaz said.
The eye as a window to the brain
The strong link between the eye and the brain has led to an emerging concept in neurobiological research: “the eye as a window to the brain” [34, 75]. Indeed, the retina is an integral part of the CNS, displaying striking similarities to the other CNS structures in terms of anatomy, response to insult, and immunology [75].
This strong interplay between the brain and the retina is underscored by the presence of retinal manifestations in many neurodegenerative diseases or insults, e.g., Alzheimer’s disease, Huntington’s disease, multiple sclerosis, and stroke [3, 29, 86, 120, 124].
Furthermore, the retina is the most accessible part of the CNS, and the unique properties of the eye in terms of in vivo imaging, manipulation, and administration of compounds, can be exploited for both fundamental and clinical research of CNS diseases [34, 75].
In addition, some retinal manifestations are suggested to be present and/or detectable before typical cerebral manifestations [34, 93, 125, 138].
Indeed, given that the resolution of current retinal imaging techniques is at least an order of magnitude higher than available brain imaging techniques, they might be able to detect more subtle changes (i.e., earlier disease stages). The eye thus can offer an alternative approach, which can complement brain examinations/research, especially in terms of early diagnosis and disease monitoring [77, 93].
Thanks to the accessibility and the transparency of the anterior segment of the eye (in contrast to the bony skull that covers the brain), an array of technologies allows non-invasive, high-resolution imaging of the retina, e.g., confocal scanning laser ophthalmoscopy (cSLO), optical coherence tomography (OCT), adaptive optics, and oximetry [13].
In combination with electroretinography (ERG), visual-evoked potentials, and visual function tests, these techniques are routinely used to detect (early) changes in retinal morphology, blood flow, electrophysiology, and visual performance. Especially relevant, or most commonly used, to the study of PD in the retina, are OCT, cSLO, and ERG.
Optical coherence tomography provides structural information on ocular tissues with 1-to-10 μm resolution, and enables direct observation of remodelling of the retinal tissue and morphometric measurements of retinal layer thickness [6]. A second non-invasive, diagnostic imaging technique for the retina is confocal scanning laser ophthalmoscopy, which produces an en face view of the retina on which changes in the optic nerve head, vasculature, and retinal tissue morphology can be assessed [147].
In addition, cSLO can be used to detect fluorescent signals, an application upon which the ‘Detection of Apoptosing Retinal Cells’ (DARC) technique for single-cell imaging of apoptotic cells is based [14, 44]. Electroretinography, finally, together with visual-evoked potentials (VEP), has proven very sensitive in revealing alterations in the electrical responses of various retinal cell types, thereby pinpointing the exact circuitries affected. Pattern electroretinography (PERG), more specifically, uses pattern-reversal stimuli to capture retinal ganglion cell activity [46].
Retinal manifestations of Parkinson’s disease
Visual symptoms in Parkinson’s disease patients
Almost 80% of PD patients report at least one visual symptom, resulting from either defects in primary vision, such as visual acuity [8, 54, 61, 85], spatial contrast sensitivity [19, 21], and colour vision [52, 114, 122, 130], or deficits in more complex visual tasks [8, 9, 11, 49, 110, 135, 148]. One of the most prevalent visual dysfunctions among PD patients is impaired colour vision, which is reported to manifest several years before diagnosis and to correlate with disease progression (i.e., motor function) [38, 52, 114, 122].
Next, one in three PD patients has issues with seeing fine detail due to reduced visual acuity and visuospatial processing, a feature that is also correlated with motor performance [61, 85]. Third, some of the earliest visual abnormalities due to PD, together with colour recognition, are poor vision at dim light conditions and the disability to perceive small increments of light versus dark, which are manifestations of reduced contrast sensitivity.
Given its progression and correlation with disease severity, reduced contrast sensitivity has been suggested to be a valuable biomarker for early PD [19, 38, 49, 52, 109, 114, 134, 138, 146]. Notably, changes in colour discrimination and contrast sensitivity have shown a discriminatory power for early diagnosis that outweighs non-motor symptoms such as hyposmia and sleep disturbance [148].
Besides these manifestations in the neuroretina of PD patients, other parts of the eye are reported to be affected as well, e.g., the pupil and lens, leading to blurred vision due to disordered pupil reactivity and cataract, respectively [94].
While visual dysfunctions experienced by PD patients are at least in part to be attributed to defects in subcortical and cortical areas for visual processing [16, 86], electrophysiological tests, and structural imaging prove that they are (also) the result of local retinal pathogenesis. Indeed, several OCT studies have demonstrated retinal nerve fibre layer thinning in PD patients as compared to age-matched controls [12, 24, 70, 75, 84].
In addition, studies comparing retinal nerve fibre layer thickness in the four retinal quadrants demonstrated that the temporal quadrant is most commonly affected. This is of particular interest, given that this quadrant is typically affected in mitochondrial optic neuropathies [34, 68, 79] and that PD has been linked to mitochondrial dysfunction [4, 58, 68, 91].
PD patients also display thinning of the inner retinal layers: the ganglion cell layer, inner plexiform layer, and inner nuclear layer [1, 22, 71, 128, 133]. Together, these observations are suggestive of a loss of retinal ganglion cells (cfr. retinal nerve fibre layer, ganglion cell layer, and inner plexiform layer changes) and a thinning of the dopaminergic plexus (cfr. inner plexiform layer and inner nuclear layer changes) [19, 32, 88, 139].
These findings are corroborated by ERG studies, which have shown significant changes in ERG responses in PD patients. On one hand, decreases in the amplitude of the photopic b-wave and scotopic oscillatory potentials were observed [27, 48, 57, 95], which are diagnostic indicators of the functioning of retinal interneurons, which comprise, amongst many other cells, dopaminergic amacrine cells [10].
Of note, decreased dopamine levels have been shown to affect scotopic oscillatory responses [95]. On the other hand, a reduced amplitude and delayed latency of the P50 component of the PERG were recorded [45, 48, 69, 92, 103,104,105, 121], for which there is also evidence that these can be correlated with disturbed dopamine signalling affecting the retinal ganglion cells [10, 120].
Histopathological evidence
In the brain, the defining neuropathological features of PD are loss of dopaminergic neurons and accumulation of αSYN inclusions, both of which are confirmed post-mortem via histopathological examination. While in vivo evidence from OCT and ERG studies points to dopaminergic degeneration in the retina as well, only one paper reveals post-mortem evidence for a reduction in dopamine levels to support and better understand these findings [30, 39, 51].
Nonetheless, dopaminergic involvement is underscored by the transient improvement in visual function tests and PERG responses upon administration of L-DOPA and by post-mortem studies revealing reduced retinal tyrosine hydroxylase expression [7, 51, 61, 120].
In addition, a recent study reported reduced melanopsin-immunoreactive retinal ganglion cell (mRGC) density and complexity of the melanopsin plexus in the retina of PD patients. While being involved in circadian rhythm, commonly affected in PD, a subset of these mRGCs also receives dopaminergic input, and their loss may thus be linked to a diminution of retinal dopamine levels (i.e., a loss of their main synaptic inputs) [99]. Furthermore, little histopathological proof for retinal accumulation of αSYN inclusions has been gathered thus far.
Studies on αSYN and phosphorylated αSYN in the retina of Parkinson’s disease patients
Between 2014 and 2018, four studies have investigated the presence of αSYN or phospho-αSYN in the retina of PD patients, in a total of 27 PD patients and 24 age-matched controls [15, 20, 53, 98]. In the years before, endogenous αSYN had already been described for the healthy retina [72, 82].
The methodology and primary findings of these studies are summarised in Table 1 and Fig. 1. First of all, in healthy subjects, two independent studies described the presence of endogenous αSYN in the inner nuclear layer and in the ganglion cell layer [72, 82].
In addition, the outer segments of the photoreceptors in the photoreceptor layer and their terminals in the outer plexiform layer were strongly immunoreactive for αSYN, and αSYN-positive cell bodies and neurites were detected in the inner nuclear layer and the inner plexiform layer, respectively [82].
These observations might be reconciled with αSYN in its monomeric form, which is predominantly located in the cytoplasm of presynaptic nerve terminals of both dopaminergic and non-dopaminergic neurons, and of which the physiological function has been suggested to be related to synaptic plasticity, vesicle trafficking, and neurotransmission [41].
Table 1 Overview of the studies investigating the presence of retinal (phospho-)αSYN expression and/or inclusions in human subjects, with the main findings on localisation and appearance of (phospho-)αSYN and study design
From: Retinal α-synuclein deposits in Parkinson’s disease patients and animal models
| Observation | Retinal layers containing αSYN | Indications of αSYN aggregation | Subjects | Tissue specimens | Antibody | References |
|---|---|---|---|---|---|---|
| Non-PD patients | ||||||
| Endogenous αSYN in somata and neurites of INL neurons and in outer segments of photoreceptors | GCL, IPL, INL, OS | None | n.s. | Sections | Polyclonal goat anti-αSYNa, Santa Cruz | [82] |
| Endogenous αSYN in somata of INL neurons Proportion of patients with intracytoplasmic inclusions of αSYN and ubiquitin in INL was higher with ageing | INL | Intracytoplasmic inclusions of co-labelled αSYN and ubiquitin | 19 | Sections | Monoclonal mouse anti-αSYNa, clone LB509, Zymed; Polyclonal rabbit anti-ubiquitin, Dako | [72] |
| PD patients | ||||||
| Healthy controls: faint diffuse αSYN immunoreactivity in somata and neurites of few INL neurons PD patients: more intense αSYN immunoreactivity and presence of Lewy neurites in IPL; 8-10 µm globular Lewy bodies in INL (border IPL); intra- and extracellular inclusions in GCL | GCL, IPL, INL | αSYN+ inclusions in GCL αSYN+ Lewy neurites IPL αSYN+ Lewy bodies in INL (Note, phospho- αSYN not assessed) | 3 PD 8 Control | Sections | Monoclonal mouse anti-αSYNa, clone 4B12, Covance | [20] |
| PD patients: phospho-αSYN immunoreactive nerve fibres in RNFL, GCL and IPL; little evidence for intraneuronal inclusions | RNFL, GCL, IPL | Intraneuronal phospho-αSYN+ inclusions in one case | 9 PD 4 Control | Wholemounts | Rabbit anti-phospho-Ser-129 αSYN, Dr. Haruhiko Akiyama | [15] |
| PD patients: αSYN expression in somata of INL neurons and in outer segments of photoreceptors, similar to controls PD patients: phospho-αSYN in axonal fibres, neuronal somata and neurites in the RNFL, GCL and IPL of all PD patients, but not in controls; phospho-αSYN-positive intraneuronal inclusions in RGCs; phospho-αSYN-positive Lewy-like neurites with dystrophic morphology PD patients: retinal Lewy-type synucleinopathy status positively correlates with disease stage | αSYN: ubiquitous, mainly OS, INL phospho-αSYN: RNFL, GCL, IPL | Phospho-αSYN+ Lewy-like bodies and neurites | 9 PD 6 Control | Wholemounts | Rabbit anti-phospho-Ser-129 αSYN, Dr. Haruhiko Akiyama | [98] |
| PD patients: diffuse cytoplasmic αSYN immunoreactivity in INL and GCL neurons, similar to controls | GCL, INL | None | 6 PD 6 Control | Sections | Monoclonal mouse anti-αSYNb, clone 42, BD Transduction Laboratory | [53] |
- PD Parkinson’s disease, CJD Creutzfeldt–Jakob disease, RNFL retinal nerve fibre layer, GCL ganglion cell layer, INL inner nuclear layer, IPL inner plexiform layer, INL inner nuclear layer, OS outer segments of photoreceptors, RGCs retinal ganglion cells
- aAntibody recognising the C terminus of αSYN
- bAntibody recognising the NAC region of αSYN
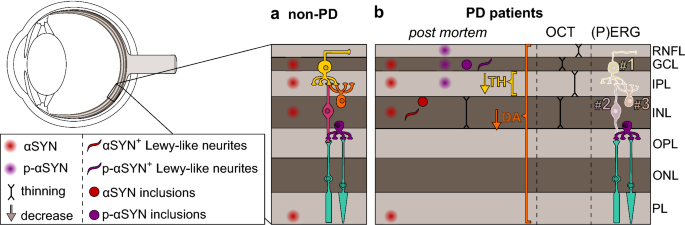
In PD retinas, however, additional αSYN was found that had accumulated into insoluble aggregates, undergone conformational changes, and/or abnormal phosphorylation. First, Bodis-Wollner et al. [20] demonstrated a distinct pattern of intense αSYN immunoreactivity in different retinal layers: upon immunostaining for native αSYN, 8–10-μm globular inclusions were reported in inner nuclear layer neurons, αSYN-positive neurites were seen in the inner plexiform layer, and both intra- and extracellular αSYN-positive inclusions were found in the ganglion cell layer.
Importantly, this immunoreactivity pattern was seen in the central and peripheral retina and was consistent for all eight PD retinas investigated. Furthermore, morphometric analysis of the retina pointed to a thinner inner retina, especially the inner nuclear layer.
Although the authors suggest that this latter change might be due to a loss of dopaminergic cells, one should consider that the fraction of dopaminergic cell bodies in the inner nuclear layer is limited and, therefore, unlikely to account for substantial inner nuclear layer thinning.
In fact, there is more evidence pointing to the loss of dopaminergic processes in the inner plexiform layer, which constitute a large plexus and, therefore, may account for substantial inner retinal thinning when lost [19, 32, 88, 139].
More evidence for the accumulation of αSYN deposits in the retina was gathered by Ortuno-Lizaran et al. [98] and Beach et al. [15], who both investigated the presence of αSYN phosphorylated at serine-129. These studies investigated nine PD patients each, and found phospho-αSYN immunoreactivity in all nine patients and in seven out of nine PD patients, respectively.
Immunostaining for phospho-αSYN was also found in a subset of incidental Lewy body disease patients (three of four and one of three, respectively), yet was absent in all control retinas (four and six subjects, respectively). Ortuno-Lizaran et al. [98] described that phospho-αSYN deposits were present in axonal fibres, dendrites, and neuronal perikarya.
Although the cell morphology and soma size of the affected neurons varied, double stainings confirmed that these cells were all retinal ganglion cells, and no inclusions were seen in (dopaminergic) amacrine cells [98]. In contrast, Beach et al. did not observe any intraneuronal inclusions, except for one case, and concluded that the phospho-αSYN immunopositive fibres seen in their study must be centrifugal/retinopetal fibres originating from the brain or retinal ganglion cell (RGC) axons [15].
Strikingly, both Bodis-Wollner et al. [20] and Ortuno-Lizaran et al. [98] claimed that the retinal αSYN inclusions that they found displayed characteristics reminiscent of classic Lewy bodies and neurites. According to Bodis-Wollner et al., intracytoplasmic αSYN staining in the inner nuclear layer and ganglion cell layer was condensed into globular inclusions replacing other cell components, while Lewy-like neurites were observed in the IPL.
Ortuno-Lizaran et al. performed a phospho-αSYN staining and revealed deposits in neuronal perikarya, as well as phospho-αSYN-positive neurites resembling Lewy neurites with typical dystrophic morphology. Finally, abnormal beading, swollen axonal segments, and increased tortuosity and swelling of the dendrites suggested that the phospho-αSYN-positive neurons found in this study were dysfunctional and/or undergoing neurodegeneration [20, 98].
Importantly, in the study by Bodis-Wollner et al., an antibody for total non-modified αSYN was used, while Ortuno-Lizaran et al. used a phospho-αSYN specific antibody. Given that phospho-αSYN is an indicator of increased αSYN aggregation/toxicity and more relevant than total or native (unphosphorylated) αSYN to evaluate Lewy body pathology (as highlighted in Table 2), caution is thus warranted when interpreting the findings of Bodis-Wollner et al., and overall, one can assume phospho-αSYN to have a higher value as PD biomarker.
Table 2 Table outlining the basic biochemistry from soluble αSYN to aggregates to deposits
From: Retinal α-synuclein deposits in Parkinson’s disease patients and animal models
| Soluble αSYN | αSYN aggregates | αSYN deposits | |
|---|---|---|---|
| Native αSYN | + | + | + |
| Phosphorylated αSYNa | − | ± | + (+) |
| Morphological interpretation of Lewy body/neurite characteristics | − | − | + |
- Progression of αSYN aggregation in the tissues is accompanied by increased phosphorylation of the Serine-129 residue of αSYN and the formation of morphologically distinct deposits (the so-called Lewy bodies and Lewy neurites). Typically, Lewy bodies are spherical intraneuronal cytoplasmic inclusions, characterised by a dense core, concentric lamellar bands, and (at least for some types of Lewy bodies) narrow pale halos, and immunoreactivity for αSYN (and other aggregation markers, such as p62 and ubiquitin), while Lewy neurites appear as αSYN-positive, thread-like dystrophic neurites [129]. Interpretation of immunohistological stainings of αSYN should ideally take into account these defining features to describe the pathological status of the tissue under investigation
- aOf note, whether phosphorylation comes before or mainly after the formation of αSYN deposits is still debated [76, 100, 153]
Although Ortuno-Lizaran et al. [98] reported that phospho-αSYN accumulation was sparse, with relatively few retinal ganglion cells affected, quantification of the number of immunopositive retinal ganglion cells in the nasal-inferior quadrant revealed a strong positive correlation between Lewy‐type synucleinopathy density in the retina and brain.
Furthermore, retina Lewy‐type synucleinopathy density score correlated with the Unified Parkinson’s disease pathology stage. Importantly, retinas from patients with incidental Lewy body disease were included in this study, besides cases of moderate-to-severe PD [81].
Hereby, this study was the first to present evidence that phospho-αSYN accumulates in the retina in parallel with the brain, including in the early stages preceding development of clinical signs of Parkinsonism or dementia.
Notably, Ortuno-Lizaran et al. [98] also looked at native αSYN, albeit using a different antibody than Bodis-Wollner et al. [20], and found ubiquitous expression in the photoreceptor outer segments and the amacrine cells in the inner nuclear layer, that was identical in PD patients and healthy subjects [98].
Likewise, Ho et al. [53] reported diffuse cytoplasmic staining of native αSYN in the inner nuclear layer and ganglion cell layer that was comparable in six PD patients and six age-matched controls. Both studies failed to find Lewy bodies in the retina of PD patients. On the one hand, this reconciles the notion that phospho-αSYN is the preferred neuropathological marker of disease (cfr. above and Table 2).
On the other hand, these contradicting findings point out the shortcomings of the studies that are currently available. First of all, due to the difficulty in obtaining high-quality post-mortem human retinas, the number of patients in each of the studies is relatively low, and at this point still does not allow to make extrapolations of the prevalence and specificity of retinal αSYN deposits to the general population.
Second, different αSYN species (native versus phosphorylated) have been investigated, being indicative of a distinct pathological status; total αSYN antibodies stain both soluble and aggregated forms of αSYN, while phosphorylation-specific antibodies identify αSYN species with increased aggregation propensity and toxicity (Table 2).
Third, different methodologies might underlie the different results. These include differences in and/or lack of immunohistochemical methods, comprehensive retinal tissue processing and wholemount techniques, and quality of available αSYN antibodies [31].
In particular, for αSYN immunostainings, the time between death of the subject and fixation of the tissue, the fixation time, and antigen retrieval method have been shown to lead to variability in staining results [2, 66, 106]. Furthermore, whereas most antibodies against αSYN are directed against an epitope at the C terminus of αSYN (Table 1) [37], the exact amino acid sequences differ, and this may (co-)account for variability in the staining results.
Finally, it has been suggested that the sparse retinal αSYN inclusions can easily be missed when evaluating a limited number of retinal sections; thus, retinal wholemount preparations are preferred over sections [97, 98].
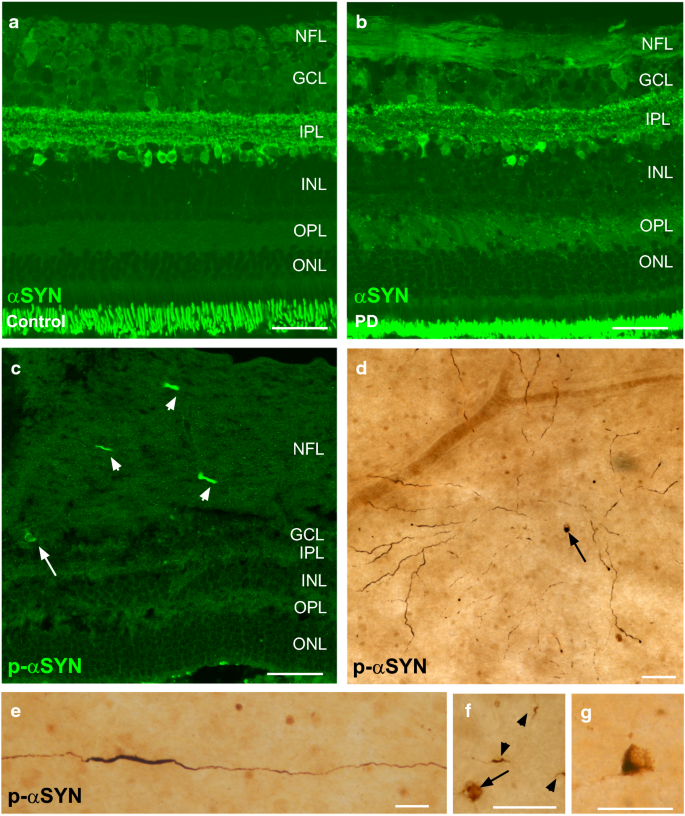
Figure 2 shows a series of unpublished immunostainings for total, unmodified αSYN and phospho-αSYN on retinal sections and flatmount preparations of PD subjects and healthy controls, according to the methods published by Ortuno-Lizaran et al. [98].
As described in their recent manuscript, phospho-αSYN staining was found in axonal fibres, dendrites, and neuronal perikarya of PD retinas only, and phospho-αSYN-positive neurites displayed features of a dystrophic morphology (Fig. 2c–g). Importantly, immunostainings for native αSYN revealed identical staining patterns in PD and control retinas (Fig. 2a, b), again corroborating phospho-αSYN but not total/native αSYN as a potential biomarker of PD.
In conclusion, half a dozen immunohistological studies have investigated the presence of native and phosphorylated αSYN in the retina. Their major findings are summarised in Fig. 1. They showed that native αSYN in healthy control retinas is most abundant in the photoreceptor outer segments, in the outer plexiform layer and in neurons and their processes in the inner nuclear layer and inner plexiform layer.
Results from studies looking at PD retinas are somewhat contradictory, yet overall, they do provide evidence for the presence of phospho-αSYN-positive cell bodies and neurites in the retinal nerve fibre layer, ganglion cell layer, and inner plexiform layer.
Several of these studies suggest that ganglion cells and (dopaminergic) amacrine cells are preferentially affected and that this also corresponds to the structural and functional changes that have been reported in OCT and ERG studies in PD patients [12, 24, 75, 84].
It currently remains unclear, however, how the significant structural effects can be reconciled with the relatively low numbers of dopaminergic amacrine cells in the retina, yet very large dopaminergic plexus, and detailed histological studies are warranted to univocally demonstrate the involvement of dopaminergic retinal neurons.
Finally, earlier this year, Ortuno-Lizaran et al. were the first to provide evidence that PD progression in the retina and brain is related and that retinal phospho-αSYN inclusions may provide information about the disease severity in the brain, already in an early presymptomatic stage of PD.
REFERENCE LINK : https://link.springer.com/article/10.1007/s00401-018-01956-z
Provided by Radiological Society of North America



{kind=link}