











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



With fall and winter holidays coming up, many will be pondering the relationship between food and sleep.
Researchers led by Professor Masashi Yanagisawa at the University of Tsukuba in Japan hope they can focus people on the important middlemen in the equation: bacterial microbes in the gut.
Their detailed study in mice revealed the extent to which bacteria can change the environment and contents of the intestines, which ultimately impacts behaviors like sleep.
The experiment itself was fairly simple. The researchers gave a group of mice a powerful cocktail of antibiotics for four weeks, which depleted them of intestinal microorganisms.
Then, they compared intestinal contents between these mice and control mice who had the same diet. Digestion breaks food down into bits and pieces called metabolites. The research team found significant differences between metabolites in the microbiota-depleted mice and the control mice.
As Professor Yanagisawa explains, “We found more than 200 metabolite differences between mouse groups. About 60 normal metabolites were missing in the microbiota-depleted mice, and the others differed in the amount, some more and some less than in the control mice.”
The team next set out to determine what these metabolites normally do. Using metabolome set enrichment analysis, they found that the biological pathways most affected by the antibiotic treatment were those involved in making neurotransmitters, the molecules that cells in the brain use to communicate with each other.
For example, the tryptophan–serotonin pathway was almost totally shut down; the microbiota-depleted mice had more tryptophan than controls, but almost zero serotonin.
This shows that without important gut microbes, the mice could not make any serotonin from the tryptophan they were eating. The team also found that the mice were deficient in vitamin B6 metabolites, which accelerate production of the neurotransmitters serotonin and dopamine.
The team also analyzed how the mice slept by looking at brain activity in EEGs. They found that compared with the control mice, the microbiota-depleted mice had more REM and non-REM sleep at night – when mice are supposed to be active – and less non-REM sleep during the day – when mice should be mostly sleeping.
The number of REM sleep episodes was higher both during the day and at night, whereas the number of non-REM episodes was higher during the day. In other words, the microbiota-depleted mice switched between sleep/wake stages more frequently than the controls.
Professor Yanagisawa speculates that the lack of serotonin was responsible for the sleep abnormalities; however, the exact mechanism still needs to be worked out. “We found that microbe depletion eliminated serotonin in the gut, and we know that serotonin levels in the brain can affect sleep/wake cycles,” he says.
“Thus, changing which microbes are in the gut by altering diet has the potential to help those who have trouble sleeping.”
So, this holiday season, when you’re feeling sleepy after eating tryptophan-stuffed turkey, please don’t forget to thank your gut microbes!
Tryptophan is an essential amino acid found in many protein-based foods and dietary proteins [1] including meats, dairy, fruits, and seeds. High-glycaemic index and -glycaemic load meals also increase the availability of tryptophan [2].
Levels of plasma tryptophan are determined by a balance between dietary intake [3], and its removal from the plasma as a part of its essential role in protein biosynthesis [4].
Aside from its role in protein formation, tryptophan is a precursor for a number of metabolites, most notably kynurenine and the neurotransmitter, serotonin which is the focus of this review.
Serotonin and Kynurenine
Tryptophan is the sole precursor of peripherally and centrally produced serotonin [4]. However, the second most prevalent metabolic pathway of tryptophan after protein synthesis is the synthesis of kynurenine, which accounts for approximately 90% of tryptophan metabolism [5].
Kynurenine is the precursor of kynurenic acid, an antagonist at glutamate ionotropic receptors. There is strong evidence implicating the kynurenines in behavioural and cognitive symptoms of neurological disease [6], however the relationship between the central effects of tryptophan depletion/supplementation and the kynurenine pathway is as yet not clear [7–9]. The role of kynurenine in the brain is beyond the scope of this review.
Serotonin and Its Receptors
Serotonin synthesis occurs in the periphery within the gut neurons and enterochromaffin cells and centrally within the neurons of the raphe in the brain stem.
The effects of tryptophan depletion on peripheral serotonin production will be discussed later. For central serotonin production to occur, tryptophan first needs to gain access to the central nervous system (CNS) via the blood-brain barrier. Tryptophan is a substrate for the large neutral amino-acid transporter system and competes for transport with several other amino acids essential for brain function.
This competition for transport is the basis for some acute tryptophan depletion diets (e.g., [10]). It is generally accepted that most of our tryptophan is bound to plasma albumin and hence is unavailable for transport into the brain.
This normally limits the tryptophan available for central serotonin synthesis but release of tryptophan from this pool could increase transport. In addition to free tryptophan levels, findings from exercise studies demonstrate that there must be other, currently unknown, mechanisms controlling central uptake of tryptophan [11].
Once in the CNS, L-tryptophan is hydroxylated to 5-hydroxytryptophan by the enzyme tryptophan hydroxylase type 2, the rate limiting step in brain serotonin synthesis. This is followed by subsequent decarboxylation involving the enzyme L-aromatic acid decarboxylase to serotonin (5-hydroxytryptamine, 5-HT).
Serotonin is then taken up into vesicles by the vesicular monoamine transporter isoform 2 of the raphe neurons. Degradation of serotonin is via monoamine oxidase type A and aldehyde dehydrogenase to the major serotonin metabolite 5-hydroxyindoleacetic acid (5HIAA).
Levels of serotonin are also influenced by the tryptophan-degrading enzyme, indoleamine 2,3-dioxygenase and tetrahydrobiopterin, the cofactor of tryptophan hydroxylase.
All but one subtype of the many serotonin receptors are metabotropic G protein–coupled receptors.
Multiple serotonin receptors have been found, with receptor families from 5-HT1 to 5-HT7 [12,13]. The 5-HT3 receptor is unique among the currently known serotonergic receptor subtypes in that it belongs to the ionotropic, ligand-gated ion channel family.
Serotonergic neurons innervate large areas of the human brain, with most projections arising from neuronal cell bodies in the dorsal and median raphe and neighbouring nuclei of the lower brain stem. There are projections to the hippocampus, amygdala, hypothalamus, thalamus, neocortex, and basal ganglia, although most structures receive some serotonergic innervation [14].
Through this diffuse network within the central nervous system, serotonin modulates a wide array of functions including sleep, control of appetite and temperature, and the focus of this review, mood and cognition.
Serotonin and Mood
Lowered mood is one of the major symptoms of depression, an affective disorder which is the leading cause of disability worldwide, affecting approximately 20% of the world’s population [15].
The major therapeutic agents for treating depression are antidepressants, mostly selective serotonin reuptake inhibitors or combined serotonin/noradrenaline reuptake inhibitors [16]. The mechanism of these medications is believed to be in part by increasing synaptic levels of monoamines, mainly serotonin and noradrenaline and subsequent activation of serotoninergic and noradrenergic postsynaptic and autoreceptors [17].
The therapeutic benefits of increased levels of monoamines were discovered in the middle of last century, when monoamine oxidase inhibitors and tricyclic antidepressants showed efficacy in treating depression. This led to the monoamine hypothesis where depression was thought to be caused by a deficiency in monoamine neurotransmitters [18].
However, antidepressants are only partly effective in the treatment of depression of moderate and greater severity in adults (response rates of approximately 48% compared with 30% for placebo) [19,20], suggesting that the monoamine hypothesis only partially explains depression [21,22].
The effect of serotonin on mood has been investigated using an acute tryptophan depletion technique where lowering dietary tryptophan levels causes a lowering of brain serotonin levels, allowing analysis of serotonin-dependent behaviour [23]. This is discussed in more detail below.
Serotonin and Cognition
The serotonergic system plays a role in behaviours that involve a high cognitive demand. Serotonin receptors are found in brain regions involved in learning and memory including the cortex, amygdala, and hippocampus [24].
As drug targets for cognitive improvement or enhancement, serotonin receptors have received attention with a focus on several serotonin-receptor subtypes shown to be involved in cognition and memory. Converging evidence suggests that the administration of 5-HT2A/2C or 5-HT4 receptor agonists or 5-HT1A or 5HT3 and 5-HT1B receptor antagonists prevents memory impairment and facilitates learning in situations involving a high cognitive demand.
In contrast, receptor antagonists for 5-HT2A/2C and 5-HT4, or agonists for 5-HT1A or 5-HT3 and 5-HT1B generally have opposite effects on memory and learning [25–30].
Whether serotonin plays a role in modulating cognitive function through specific effects on learning, memory and executive function are still not understood. This may be attributed partially to the differing roles of various serotonin receptor subtypes in cognition [30]. However, lowering central serotonin levels through tryptophan depletion experimentally has enabled some elucidation of the role of serotonin in different modes of learning.
Tryptophan Depletion
Initial studies aiming to deplete central tryptophan employed the irreversible tryptophan hydroxylase inhibitor, 4-chloro-DL-phenylalanine methyl ester (PCPA), which depletes serotonin by stopping the rate-limiting step in its synthesis [31]. However concerns about its toxicity and dose range largely limited its experimental use [23].
An alternative to inhibiting the synthesis enzyme for serotonin is to deplete its substrate tryptophan from the brain. Rapid dietary depletion of tryptophan allows the investigation of the effect of lowered tryptophan levels, and as such provides a paradigm for studying the role of serotonin in central processes.
The ingestion of a diet or solution containing large neutral amino acids but deficient in tryptophan induces an acute and reliable lowering of plasma tryptophan. This effect is thought to be due to the phenomena that removal of tryptophan from the diet stimulates protein synthesis in the liver, which uses up the available plasma tryptophan.
This effect has been observed experimentally in animals including mice [32], rats [33–35], and primates [36]; and in humans [37,38].
In addition to increased liver protein synthesis, the large neutral amino acids included in the diet compete with tryptophan for transport across the blood brain barrier and thus restrict the entry of tryptophan into the brain.
This depletes tryptophan, and thus serotonin, centrally. Rodent studies have shown that acute tryptophan depletion reduced tryptophan levels in the brain by up to 70% [35], with associated decreases in central serotonin and reduced 5-HT1A receptor binding [35,39]. In humans acute tryptophan depletion inhibits serotonin synthesis [40] and also lowers cerebrospinal fluid concentrations of tryptophan [41] and 5-hydroxyindoleacetic acid (5-HIAA), the major serotonin metabolite [41,42].
Tryptophan Depletion, Serotonin and Mood
Clinical and preclinical studies have used the tryptophan depletion model to investigate the idea that low serotonin synthesis is associated with depressed mood [43,44].
Clinical Studies
Tryptophan depletion studies in never-depressed individuals are variable, with no or little overall effect on lowering of mood [45,46]. Interestingly, reports of moderate mood lowering are seen more often in studies with healthy women than in studies with healthy men [47].
However in never-depressed healthy volunteers who are at high risk for depression through a familial risk factor, acute tryptophan depletion produces clear abnormalities in mood control [48,49]. Finally, in remitted depressed patients, temporarily lowering tryptophan levels can result in an acute depressive
Serotonin and Cognition
The serotonergic system plays a role in behaviours that involve a high cognitive demand. Serotonin receptors are found in brain regions involved in learning and memory including the cortex, amygdala, and hippocampus [24]. As drug targets for cognitive improvement or enhancement, serotonin receptors have received attention with a focus on several serotonin-receptor subtypes shown to be involved in cognition and memory.
Converging evidence suggests that the administration of 5-HT2A/2C or 5-HT4 receptor agonists or 5-HT1A or 5HT3 and 5-HT1B receptor antagonists prevents memory impairment and facilitates learning in situations involving a high cognitive demand. In contrast, receptor antagonists for 5-HT2A/2C and 5-HT4, or agonists for 5-HT1A or 5-HT3 and 5-HT1B generally have opposite effects on memory and learning [25–30].
Whether serotonin plays a role in modulating cognitive function through specific effects on learning, memory and executive function are still not understood. This may be attributed partially to the differing roles of various serotonin receptor subtypes in cognition [30]. However, lowering central serotonin levels through tryptophan depletion experimentally has enabled some elucidation of the role of serotonin in different modes of learning.
Tryptophan Depletion
Initial studies aiming to deplete central tryptophan employed the irreversible tryptophan hydroxylase inhibitor, 4-chloro-DL-phenylalanine methyl ester (PCPA), which depletes serotonin by stopping the rate-limiting step in its synthesis [31]. However concerns about its toxicity and dose range largely limited its experimental use [23].
An alternative to inhibiting the synthesis enzyme for serotonin is to deplete its substrate tryptophan from the brain. Rapid dietary depletion of tryptophan allows the investigation of the effect of lowered tryptophan levels, and as such provides a paradigm for studying the role of serotonin in central processes.
The ingestion of a diet or solution containing large neutral amino acids but deficient in tryptophan induces an acute and reliable lowering of plasma tryptophan. This effect is thought to be due to the phenomena that removal of tryptophan from the diet stimulates protein synthesis in the liver, which uses up the available plasma tryptophan.
This effect has been observed experimentally in animals including mice [32], rats [33–35], and primates [36]; and in humans [37,38].
In addition to increased liver protein synthesis, the large neutral amino acids included in the diet compete with tryptophan for transport across the blood brain barrier and thus restrict the entry of tryptophan into the brain.
This depletes tryptophan, and thus serotonin, centrally. Rodent studies have shown that acute tryptophan depletion reduced tryptophan levels in the brain by up to 70% [35], with associated decreases in central serotonin and reduced 5-HT1A receptor binding [35,39].
In humans acute tryptophan depletion inhibits serotonin synthesis [40] and also lowers cerebrospinal fluid concentrations of tryptophan [41] and 5-hydroxyindoleacetic acid (5-HIAA), the major serotonin metabolite [41,42].
Tryptophan Depletion, Serotonin and Mood
Clinical and preclinical studies have used the tryptophan depletion model to investigate the idea that low serotonin synthesis is associated with depressed mood [43,44].
Clinical Studies
Tryptophan depletion studies in never-depressed individuals are variable, with no or little overall effect on lowering of mood [45,46]. Interestingly, reports of moderate mood lowering are seen more often in studies with healthy women than in studies with healthy men [47].
However in never-depressed healthy volunteers who are at high risk for depression through a familial risk factor, acute tryptophan depletion produces clear abnormalities in mood control [48,49]. Finally, in remitted depressed patients, temporarily lowering tryptophan levels can result in an acute depressive
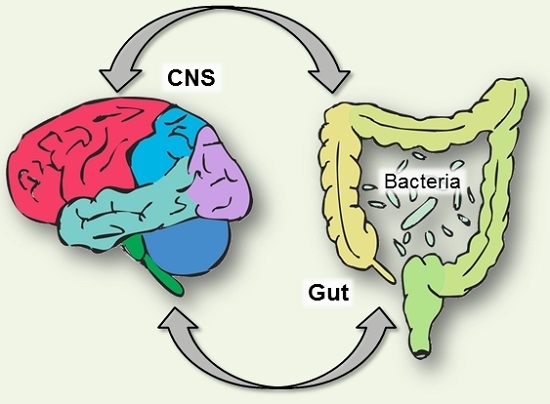
Tryptophan and the Gut Microbiota
Another piece of the serotonin puzzle involves the resident community of microorganisms that have colonised the digestive tract.
The gut microbiota is primarily found in the large intestine, but smaller numbers can be found throughout the gastrointestinal tract [99]. Cross-talk between the gastrointestinal epithelium and enteric flora contributes to functions such as immune responses and regulation of hormones, and is proving to be critical to the maintenance of both homeostasis and health (Figure 1).
How the bacterial community establishes early in life [100], or changes across the lifespan, can have consequences on the metabolism of tryptophan, and thus the serotonergic system.
A balance is needed between bacterial utilization of tryptophan and the tryptophan necessary for serotonin synthesis in both enteric and central nervous systems [101].
There is both direct and indirect regulation of tryptophan and serotonin in the gut by the resident microbiota. Indirect regulation of tryptophan availability and serotonin formation by the gut microbiota is primarily via the kynurenine pathway.
As noted, the synthesis of kynurenine accounts for approximately 90% of tryptophan metabolism [5]. Recent evidence for direct regulation comes from germ-free animals that are laboratory-raised and are gut microbiota-deficient.
These animals show increased levels of circulating tryptophan [102] and decreased serotonin [103]. When these animals have tryptophan metabolising bacteria introduced to their gut, circulating levels of tryptophan fall, with this alteration accompanying a sex-specific effect on hippocampal serotonin concentrations in male germ-free animals [102].
Within the brain, an increase in hippocampal serotonin levels and turnover was observed, along with a decrease in anxiety-like behaviour, demonstrating the influence of gut microbiota on both behavioural correlates and brain neurochemistry [104].
Interestingly, these animals also displayed a reduction in brain-derived neurotrophic factor messenger RNA levels and reduced expression of the synaptic signalling genes PSD-95 and synaptophysin in regions of the brain responsible for motor control and anxiety such as the striatum [104].
In irritable bowel syndrome, changes in the balance of microbiota are associated with symptomatology as well as alterations to both gut and brain serotonin levels [105,106]. Moreover, the expression of toll-like receptors, which act to alert the body to pathogens, are altered in both plasma and colonic samples from irritable bowel syndrome patients [107,108]. Recent data also shows that bacterial products such as short chain fatty acids can upregulate serotonin production by the enterochromaffin cells [109].
Behaviour and the Gut Microbiome
As discussed, central serotonin plays a major role in mood and cognition. An influence of gut microbiota on behaviour is becoming increasingly evident, via a variety of proposed mechanisms including changes to tryptophan uptake and serotonin synthesis.
Germ-free mice display less anxiety-like behaviours than their traditionally colonised counterparts [102,110]. Meanwhile, chronic treatment with lactic acid bacteria Lactobacillus rhamnosus to mice induced alterations in GABA receptors in cortical hippocampus, and amygdala in comparison with control-fed mice, while also reducing stress-induced corticosterone levels and anxiety- and depression-related behaviour [111].
Interestingly, these effects were not found in vagotomized mice, identifying the vagus as a major modulatory communication pathway between the gut bacteria and the brain [111] (Figure 1A).
In animal models of depression, both environmental [112] and surgical [113], animals display depressive-like behaviour and an altered intestinal microbial profile. These findings have now been replicated within a clinical population. In a recent study in major depression patients, several predominant genera were found in significantly different levels between the depressive and control groups showing either a predominance of some potentially harmful bacterial groups or a reduction in beneficial bacterial genera [114].
The influence of the gut microbiota on behaviour also extends to cognitive function in preclinical models, though all animal behavioural testing has an anxiety component, this suggests that cognitive
deficits are not observed without a degree of stress. Mice infected with an enteric pathogen exhibited working memory dysfunction [115], and socially-associated behavioural impairment [116] but only after acute water stress. Clinically, there is discussion of the engagement of gut microbial flora in the pathogenesis of Alzheimer’s disease, but at this stage this is speculative [117,118].
Tryptophan Depletion and the Gut-Brain Axis
The central control of pain is an important component of irritable bowel syndrome and serotonin has been shown to play a role. In healthy women, a painful balloon distension to the rectum resulted in increased brain activity as shown by functional magnetic resonance imaging.
When these stimuli were repeated during acute tryptophan depletion, there was an enhanced response in the amygdala, emotional arousal areas, and homeostatic afferent networks. There was also a decrease in negative feedback inhibition of the amygdala.
When these tests were repeated in women with constipation-predominant irritable bowel syndrome, a similar pattern of brain activity was observed. This suggests that there are enhanced change in brain activity, namely the homeostatic afferent network and the emotional arousal network, after aversive visceral stimulation [119,120].
In addition, cognitive performance is altered in irritable bowel syndrome [121]. Female patients with irritable bowel syndrome and healthy controls underwent a battery of neuropsychological tests after a placebo or acute tryptophan depletion. The results showed that acute tryptophan depletion produces decreased hippocampal-mediated cognitive performance [67].
A similar test in female patients with diarrhea-predominant irritable bowel syndrome and healthy controls showed acute tryptophan depletion was significantly associated with impaired immediate and delayed recall performance in an affective memory test, though there was no difference in scores between patient and control groups [121].
These patients also showed an enhanced visceral perception to an aversive visceral stimulus during acute tryptophan depletion similar to the study by Labus et al [120].
Interestingly, acute tryptophan depletion has not been shown to have an effect on mucosal concentrations of serotonin or the metabolite 5-hydroxyindoleacetic acid [122]. However, acute tryptophan depletion studies investigating effects on regulation of gastrointestinal motility and sensation have shown lowered plasma tryptophan decreased the sensation of nausea during balloon distension without affecting gastric sensitivity and compliance.
Acute tryptophan depletion also enhanced the postprandial intragastric volume increase, but this was not reflected by an increased nutrient intake [123]. In contrast, motor function of the rectum during acute tryptophan depletion was tested in female patients with diarrhea-predominant irritable bowel syndrome [124]. While the patient group had significantly altered rectal motor function, acute tryptophan depletion did not alter this.
Significant associations of tryptophan hydroxylase 1 gene polymorphisms, which may modify levels of circulating serotonin, are observed with irritable bowel syndrome-related cognitions in female patients. Employing the Cognitive Scale for Functional Bowel Disorders, tryptophan hydroxylase 1 gene polymorphisms were associated with negative cognitions regarding pain and anxiety around bowel movement.
These polymorphisms were also associated with reductions in quality of life scores, in particular mental health and energy subscales, suggesting that subsets of the tryptophan hydroxylase 1 gene may impact the onset and course of irritable bowel syndrome, along with symptom severity and the emotional consequences of living with this disorder [125].
reference link : https://doi.org/10.3390/nu8010056
Source: University of Tsukuba



{kind=link}