










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



In uno studio approfondito su come COVID-19 colpisce il cervello di un paziente, i ricercatori del National Institutes of Health hanno costantemente individuato i segni distintivi del danno causato da vasi sanguigni cerebrali assottigliati e che perdono in campioni di tessuto di pazienti deceduti poco dopo aver contratto la malattia.
Inoltre, non hanno visto segni di SARS-CoV-2 nei campioni di tessuto, suggerendo che il danno non sia stato causato da un attacco virale diretto al cervello. I risultati sono stati pubblicati come corrispondenza nel New England Journal of Medicine.
I nostri risultati suggeriscono che ciò potrebbe essere causato dalla risposta infiammatoria del corpo al virus “, ha detto Avindra Nath, MD, direttore clinico presso il National Institute of Neurological Disorders and Stroke (NINDS) del NIH e autore senior dello studio.
“Ci auguriamo che questi risultati aiuteranno i medici a comprendere l’intero spettro di problemi che i pazienti possono soffrire in modo da poter trovare trattamenti migliori”.
Sebbene COVID-19 sia principalmente una malattia respiratoria, i pazienti spesso sperimentano problemi neurologici tra cui mal di testa, delirio, disfunzione cognitiva, vertigini, affaticamento e perdita dell’olfatto. La malattia può anche indurre i pazienti a soffrire di ictus e altre neuropatologie.
Diversi studi hanno dimostrato che la malattia può causare infiammazioni e danni ai vasi sanguigni. In uno di questi studi, i ricercatori hanno trovato prove di piccole quantità di SARS-CoV-2 nel cervello di alcuni pazienti. Tuttavia, gli scienziati stanno ancora cercando di capire come la malattia colpisce il cervello.
In questo studio, i ricercatori hanno condotto un esame approfondito di campioni di tessuto cerebrale di 19 pazienti che erano morti dopo aver sperimentato COVID-19 tra marzo e luglio 2020.
I campioni di 16 pazienti sono stati forniti dall’Office of the Chief Medical Examiner di New York City, mentre gli altri 3 casi sono stati forniti dal dipartimento di patologia dell’Università di Iowa College of Medicine, Iowa City. I pazienti sono morti in una vasta gamma di età, dai 5 ai 73 anni.
Sono morti da poche ore a due mesi dopo aver segnalato i sintomi. Molti pazienti presentavano uno o più fattori di rischio, inclusi diabete, obesità e malattie cardiovascolari. Otto dei pazienti sono stati trovati morti a casa o in ambienti pubblici. Altri tre pazienti sono crollati e sono morti all’improvviso.
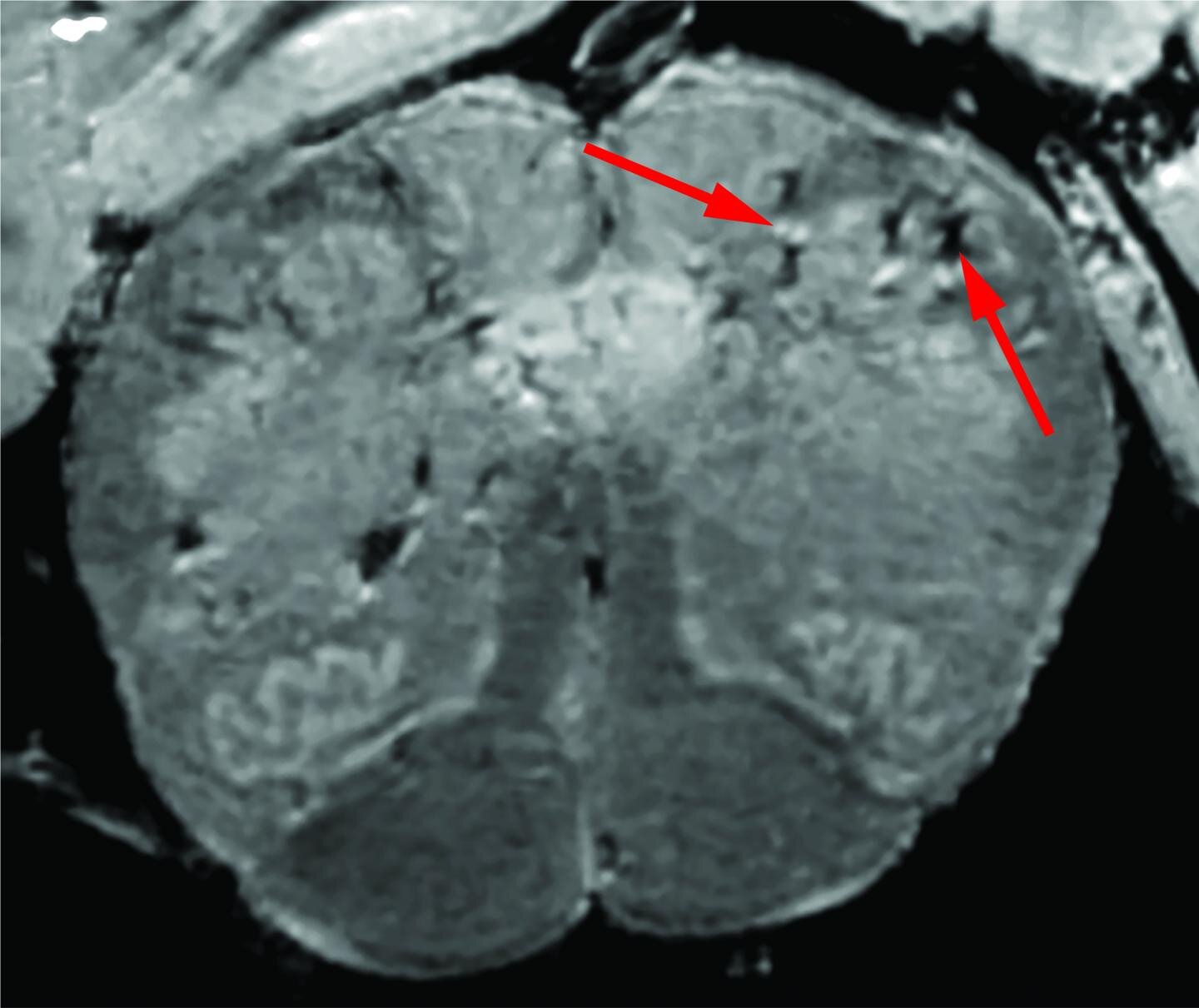
Inizialmente, i ricercatori hanno utilizzato uno speciale scanner per risonanza magnetica (MRI) ad alta potenza che è da 4 a 10 volte più sensibile della maggior parte degli scanner MRI, per esaminare i campioni dei bulbi olfattivi e del tronco cerebrale di ciascun paziente.
Si ritiene che queste regioni siano altamente suscettibili a COVID-19. I bulbi olfattivi controllano il nostro senso dell’olfatto mentre il tronco cerebrale controlla la nostra respirazione e la frequenza cardiaca. Le scansioni hanno rivelato che entrambe le regioni avevano un’abbondanza di punti luminosi, chiamati iperintensità, che spesso indicano infiammazione, e macchie scure, chiamate ipointensità, che rappresentano il sanguinamento.
I ricercatori hanno quindi utilizzato le scansioni come guida per esaminare i punti più da vicino al microscopio. Hanno scoperto che i punti luminosi contenevano vasi sanguigni più sottili del normale e che a volte perdevano proteine del sangue, come il fibrinogeno, nel cervello.
Questo sembrava innescare una reazione immunitaria. Le macchie erano circondate da cellule T del sangue e dalle cellule immunitarie del cervello chiamate microglia. Al contrario, le macchie scure contenevano vasi sanguigni coagulati e che perdevano ma nessuna risposta immunitaria.
“Siamo rimasti completamente sorpresi. In origine, ci aspettavamo di vedere danni causati dalla mancanza di ossigeno. Invece, abbiamo visto aree multifocali di danno che di solito sono associate a ictus e malattie neuroinfiammatorie “, ha detto il dott. Nath.
Infine, i ricercatori non hanno visto segni di infezione nei campioni di tessuto cerebrale anche se hanno utilizzato diversi metodi per rilevare materiale genetico o proteine da SARS-CoV-2.
“Finora, i nostri risultati suggeriscono che il danno che abbiamo visto potrebbe non essere stato causato dal virus SARS-CoV-2 che infetta direttamente il cervello”, ha detto il dott. Nath.
“In futuro, abbiamo in programma di studiare come COVID-19 danneggia i vasi sanguigni del cervello e se questo produce alcuni dei sintomi a breve e lungo termine che vediamo nei pazienti”.
I coronavirus (CoV) appartengono all’ordine Nidovirales e alla famiglia Coronaviridae che è raggruppata in quattro generi basati sulla filogenesi: Alphacoronavirus, Betacoronavirus, Gammacoronavirus e Deltacoronavirus (International Committee for Taxonomy of Virus; https://talk.ictvonline.org/).
I CoV sono particelle avvolgenti e sferiche di diametro compreso tra 80 e 120 nm con strutture a forma di corona. Questa famiglia contiene il genoma di RNA più grande e non segmentato, formato con un senso positivo a filamento singolo di dimensioni comprese tra 27 e 32 kb [1]. Il genoma di CoV condivide l’organizzazione strutturale, sebbene differisca nel numero e nella sequenza delle coppie di basi, anche tra i CoV strettamente correlati. I frame di lettura aperti 1a / b (ORF1a e ORF1b), situati all’estremità 5 ‘codificano componenti non strutturali, le poliproteine pp1a e pp1b.
Due proteasi virali scindono queste proteine per generare 16 proteine non strutturali (da nsp1 a nsp16), inclusa la RNA polimerasi RNA-dipendente (RdRP), un’importante proteina coinvolta nella trascrizione e replicazione del genoma. L’estremità 3 ‘include ORF che codificano quattro principali proteine strutturali: la glicoproteina di superficie spike (S), una piccola proteina dell’involucro (E), la proteina di membrana (M) e la proteina nucleocapsidica (N) che copre l’RNA (Fig. ( Fig.1) .1). Inoltre, il genoma del CoV mantiene geni che codificano per proteine accessorie indispensabili per l’adattamento e la virulenza e che hanno successo per l’ospite specifico [2, 3].

Organizzazione del genoma di SARS-CoV-2. La dimensione del genoma SARS-CoV-2 è di circa 32 kb ed è un senso positivo a filamento singolo di RNA che codifica 16 proteine non strutturali (estremità 5 ′) e 4 proteine strutturali (estremità 3 ′) (S, E, M, e N) e 6 proteine accessorie. Genoma di SARS-CoV-2. Il genoma contiene una coda PoliA all’estremità 3 ‘
I Betacoronavirus sono patogeni zoonotici che hanno origine animale selvatico, ad esempio origine pipistrello o roditore. Questo gruppo include CoV umani patogeni (HCoV), come HCoV-229E, HCoV-OC43, HCoV-NL63 e HCoV-HKU1, che infettano i mammiferi e provocano malattie respiratorie lievi correlate in neonati, bambini piccoli, anziani e ospiti immunocompetenti [1, 3].
Tuttavia, il CoV correlato alla sindrome respiratoria acuta grave (SARS-CoV) e il CoV correlato alla sindrome respiratoria del Medio Oriente (MERS-CoV) causano una sindrome respiratoria grave nell’uomo che ha suscitato pandemie su larga scala durante il 2002-2003 e il 2012, rispettivamente [4, 5].
È importante sottolineare che, alla fine di dicembre 2019, la Commissione sanitaria municipale di Wuhan, in Cina, ha segnalato un gruppo di casi di polmonite atipica a Wuhan, nella provincia di Hubei, in Cina, associati a un virus che si è diffuso rapidamente in tutto il mondo.
Neurotropismo di SARS-CoV-2
Ad oggi, molti rapporti hanno descritto l’associazione tra infezioni virali respiratorie con sintomi neurologici. Esistono diversi patogeni respiratori riconosciuti che hanno accesso al sistema nervoso centrale (SNC), ad esempio, il virus respiratorio sinciziale, il virus dell’influenza, il metapneumovirus umano e gli HCoV (HCoV229E, HCoV-OC43 e SARS-CoV) [39] , che inducono manifestazioni come convulsioni febbrili o afebbrili, tra le altre encefalopatie [40, 41].
Colture primarie di astrociti e microglia umani e varie linee cellulari neuronali umane, come il neuroblastoma SK-N-SH, il neuroglioma H4 e l’oligodendrocitico MO3.13, hanno un potenziale tropismo per HCoV-OC43 [42]. Utilizzando un modello animale sperimentale, l’infezione da HCoV-OC-43 ha mostrato anche neuro-invasività e neuro-virulenza [43].
Pertanto, non è sorprendente trovare autopsie cerebrali positive alla SARS. Utilizzando l’ibridazione in situ, la sequenza genomica della SARS è stata rilevata nel citoplasma dei neuroni dell’ipotalamo e della corteccia cerebrale [44]. Inoltre, Moriguchi et al. [45] ha confermato la presenza del nuovo SARS-CoV-2 nel liquido spinale cerebrale.
In accordo, la ricerca epidemiologica e clinica ha descritto sintomi neurologici, non comuni e manifestazioni neurologiche associate all’infezione da SARS-CoV-2 (Tabella (Tabella 2) .2). Queste caratteristiche cliniche includono nevralgia, confusione, iposmia, ipogeusia e coscienza alterata, sintomi che evidenziano l’invasione neurotropica da parte di SARS-CoV-2 [19, 41].
Tavolo 2
Manifestazioni neurologiche associate all’infezione da SARS-CoV-2
| Tipo di studio e dati dei pazienti | Diagnostica neurologica, sintomi e campioni clinici per il rilevamento di SARS-CoV-2 | Riferimento |
|---|---|---|
| Serie di casi n = 4 pazienti73 Y / A maschio, 83 Y / A femmina, 80 Y / A femmina e 88 Y / A femmina | Ictus acuto Stato mentale alterato, abbassamento del viso, linguaggio confuso, debolezza del lato sinistro, emiplegia e afasia Campione non specifico | [46] |
| Case report n = 2 pazienti31 Y / A maschio, 62 Y / A femmina | Hunt and Hess emorragia subaracnoidea di grado 3 da rottura di aneurisma Mal di testa e perdita di coscienza Ictus ischemico Campione nasale | [47] |
| Case report n = 5 pazienti, <50 Y / A | Ictus dei grandi vasi Mal di testa, disartria, intorpidimento, emiplegia e ridotto livello di coscienza Campione non specifico | [48] |
| Case report n = 1 paziente41 S / A | Meningoencefalite Convulsioni, letargia, fotofobia, peggioramento dell’encefalopatia, disorientamento, allucinazioni e rigidità del collo Campione non specificato | [49] |
| Caso clinico n = 1 paziente24 S / A | Meningite / encefalite, stanchezza e febbre, vomito, convulsioni, perdita di coscienza e rigidità del collo Campione di liquido spinale cerebrale | [45] |
| Riportare retrospettivamente n = 24 maschi e 19 femmine16-85 Y / A | Ictus e ictus con tromboembolia polmonare Sindrome di Guillain -Barré Campione nasofaringeo | [37] |
| Case report n = 2 pazienti 52 Y / A maschio, 39 Y / A maschio | Varianti della sindrome di Guillain-Barré Sindrome di Miller Fisher Diplopia, instabilità dell’andatura, cefalea, anosmia e ageusia Polineurite cranica ed ageusia Campione orofaringeo | [50] |
| Caso clinico n = 1 paziente61 Y / A femmina | Sindrome di Guillain-Barré acuta Debolezza delle gambe e grave affaticamento Campione orofaringeo | [51] |
| Case reportn = 1 patient65 Y/A male | Sindrome di Guillain-Barré Quadriparesi ascendente simmetrica progressiva acuta, paresi facciale bilaterale Campione orofaringeo | [52] |
| Case report n = 5 pazienti | Campione di sindrome di Guillain-Barré nasofaringea | [53] |
| Case report n = 1 paziente | Campione di sindrome di Guillain-Barré nasofaringea | [54] |
| Caso clinico n = 1 paziente58 Y / A femmina | Encefalopatia necrotizzante emorragica acuta Stato mentale alterato Campione nasofaringeo | [55] |
Nonostante le prove che dimostrano il neurotropismo dei virus respiratori, l’esatto meccanismo di neuroinvasione realizzato dai virus rimane attualmente sconosciuto. Tuttavia, la via di invasione del SNC è stata recentemente descritta per HCoV-OC-43. Questo virus accede al sistema nervoso centrale attraverso il bulbo olfattivo, spostandosi lungo il nervo olfattivo.
Quindi, la neuro-propagazione avviene lungo le molteplici connessioni assonali che si espandono attraverso il SNC (ad esempio, propagazione da neurone a neurone o particelle diffuse) [56]. Simile a HCoV-OC-43, un modello in vivo di infezione da SARS-CoV ha suggerito che il virus entra nel cervello attraverso il bulbo olfattivo e quindi potrebbe verificarsi una diffusione transneuronale [57].
Inoltre, alcuni virus infettivi trasmessi per via ematica che colpiscono principalmente gli organi periferici hanno sviluppato strategie per contrastare la barriera emato-encefalica (BBB). Queste strategie includono l’infezione diretta delle cellule endoteliali microvascolari cerebrali che formano il BBB, un ingresso paracellulare che comporta l’alterazione delle giunzioni strette, o l’invasione del “cavallo di Troia”, attraverso il traffico di monociti / macrofagi infetti che migrano attraverso il BBB, in un in modo simile al virus dell’immunodeficienza non respiratoria 1 (HIV-1) [58, 59].
Allo stesso modo, quando i monociti primari umani vengono attivati a seguito dell’infezione da HCoV-229E e alla fine diventano macrofagi, possono invadere i tessuti, compreso il SNC [60, 61]. Inoltre, è stato riportato che attraverso l’attivazione del microendotelio cerebrale, il danno causato dalla risposta infiammatoria, consente al virus di raggiungere il SNC.
In questo senso, la neuroinvasione di SARS-CoV-2 potrebbe avvenire attraverso il trasferimento trans-sinaptico, attraverso il nervo olfattivo, l’infezione dell’endotelio vascolare o la migrazione dei leucociti attraverso il BBB [38].
SARS-CoV-2 e la funzione del BBB
Le cellule endoteliali microvascolari che formano il BBB proteggono il SNC da un’ampia varietà di tossine e microrganismi presenti nel sangue. Queste cellule esprimono proteine a giunzione stretta che limitano il movimento tra cellule adiacenti e sono attraverso trasportatori specifici e proteine recettrici che controllano l’entrata e l’uscita delle molecole provenienti dal sangue verso il parenchima cerebrale [84].
Pertanto, lo studio del danno indotto alle cellule endoteliali microvascolari rappresenta il quadro centrale per la comprensione dei meccanismi molecolari dell’infezione virale nel SNC [85, 86].
L’interruzione del BBB si verifica in caso di infezione con diversi virus neurotropici riconosciuti. L’arbovirus che appartiene alla famiglia dei Flaviviridae, come il virus del Nilo occidentale e il virus Zika, può indurre danni nel BBB causati dalla risposta della cellula ospite a fattori virali.
Esperimenti condotti utilizzando modelli in vitro e in vivo del BBB hanno dimostrato che questi virus si replicano nelle cellule endoteliali microvascolari cerebrali e inducono sottoregolazione e degradazione delle proteine a giunzione stretta che portano alla rottura del BBB [87-90]. Allo stesso modo, Bleau et al. [91] hanno valutato la capacità di CoV di entrare nel sistema nervoso centrale, utilizzando il virus dell’epatite di topo altamente epatotropico di tipo 3 e l’epatite di topo debolmente epatotropico di tipo A59.
I topi infettati di tipo 3 hanno mostrato un’invasione cerebrale correlata a una maggiore permeabilità BBB. L’effetto era associato a una diminuzione dell’espressione della proteina 1 zona occludens, della VE-caderina e dell’occludina. Poiché i CoV sono molecolarmente correlati nella loro modalità di replicazione, si ipotizza che altri tipi di CoV utilizzino un meccanismo d’azione simile per infettare le cellule endoteliali microvascolari cerebrali [92, 93].
È importante sottolineare che è stata identificata la presenza di SARS-CoV-2 nelle cellule endoteliali microvascolari cerebrali nel tessuto del lobo frontale ottenuto all’esame post-mortem da un paziente con COVID-19 [94]. Inoltre, particelle virali e sequenze del genoma virale di SARS-CoV sono state rilevate nel citoplasma dei neuroni del cervello, principalmente nell’ipotalamo e nella corteccia [44, 95]. Questa prova suggerisce che SARS-CoV-2 attraversa il BBB così come altri HCoV.
Pertanto, l’infezione da diversi virus respiratori, incluso SARS-CoV-2, influisce sull’integrità del BBB attraverso diversi meccanismi. Il virus provoca uno stress cellulare diretto, associato alla maggior parte degli effetti citotossici che portano alla degenerazione delle cellule infette, ad esempio, SARS-CoV induce l’apoptosi [96].
L’attivazione delle cellule endoteliali come parte della risposta infiammatoria provoca un aumento dell’espressione delle proteasi, come la metalloproteinasi della matrice, che promuove la degradazione delle proteine a giunzione stretta [97]. Tuttavia, è probabile che la risposta infiammatoria svolga il ruolo più importante nell’induzione del danno a BBB.
La risposta infiammatoria
L’attività regolare della neuroinfiammazione è principalmente quella di ripristinare l’omeostasi nel cervello [98]. Tuttavia, un’infiammazione prolungata del SNC e una risposta infiammatoria sistemica come risultato di un’ampia varietà di patologie come le infezioni virali possono influenzare l’integrità del BBB e ulteriori esiti nei disturbi neurologici [99, 100]. Pertanto, SARS-CoV-2 potrebbe causare danni al BBB attraverso l’attivazione della risposta immunitaria infiammatoria associata a una disregolazione attorno a questo processo [101].
L’attivazione delle cellule endoteliali microvascolari è stata associata a cambiamenti nella permeabilità del BBB. Ad esempio, in condizioni fisiologiche, la migrazione delle cellule immunitarie nel SNC è rigorosamente controllata da meccanismi che operano a livello del BBB.
In particolare, la migrazione delle cellule immunitarie circolanti nel SNC è bassa e limitata a specifici sottoinsiemi di cellule immunitarie innate e adattative, come linfociti, macrofagi e cellule presentanti l’antigene come cellule dendritiche che mantengono la sorveglianza immunitaria nel SNC [102].
Tuttavia, durante le infezioni virali, la migrazione delle cellule immunitarie è aumentata. Ciò è supportato dall’esame istopatologico del tessuto cerebrale in pazienti con SARS-CoV, dove è stata riscontrata infiltrazione patologica di monociti / macrofagi CD68 + e linfociti T CD3 + nel mesenchima cerebrale [95].
Allo stesso modo, il processo di infiltrazione, correlato alle interazioni tra le integrine β1 e β2 espresse sui leucociti e sui loro ligandi [cioè, molecole di adesione intercellulare (ICAM): ICAM-1, ICAM-2 e molecola di adesione cellulare vascolare-1 (VCAM-1 )] presenti sulla superficie delle cellule microendoteliali, che inducono stravaso attraverso il BBB in condizioni infiammatorie è stato segnalato [103-107].
L’evidenza suggerisce che l’infezione e l’attivazione delle cellule endoteliali microvascolari da parte dei tipici virus neurotrofici aumentano l’espressione delle molecole di adesione endoteliale [88]. Questa condizione facilita il traffico di cellule immunitarie infettate da virus nel sistema nervoso centrale tramite il meccanismo del “cavallo di Troia” [88].
Allo stesso modo, durante la replicazione virale nelle cellule ospiti, il danno è causato perché SARS-CoV-2 è un virus citopatico che induce il rilascio di pattern molecolari associati al danno (DAMP) [108]. I DAMP sono molecole endogene rilasciate da cellule danneggiate che interagiscono con molecole chiamate recettore di riconoscimento del pattern (PRR) che inducono nelle cellule epiteliali, nelle cellule endoteliali e nei macrofagi adiacenti uno stato di elevata infiammazione [109].
Una volta che il virus interagisce con le cellule ospiti, anche il genoma virale e le proteine virali possono essere riconosciuti dai PRR e attivati la risposta immunitaria. Diversi PRR riconoscono SARS-CoV-2, ad esempio i recettori Toll-like (TLR), che sono molecole espresse in molte linee cellulari, comprese le cellule endoteliali, i macrofagi e le cellule dendritiche. TLR3, TLR7, TLR8 e TLR9 inducono diversi percorsi di attivazione che producono citochine proinfiammatorie e altre molecole antivirali per controllare l’infezione. Tuttavia, questa risposta può essere disregolata e esacerbata la produzione di citochine [110].
Inoltre, il recettore NOD-like (NLR), altro PRR, attiva il complesso inflammasoma e induce lo stato di attivazione in alcuni tipi di cellule come i macrofagi e l’epitelio e persino nelle cellule endoteliali microvascolari che portano a un’elevata produzione di interleuchina (IL) -1β e IL-18. Tuttavia, questo meccanismo deve essere studiato in dettaglio per il nuovo CoV [111].
D’altra parte, l’RNA virale attiva molecole tipiche come il gene inducibile dell’acido retinoico 1 e il gene 5 associato alla differenziazione del melanoma e induce uno stato antivirale, in cui vengono secreti interferoni (IFN) (principalmente IFN di tipo I e III). Gli interferoni sono molecole importanti per eliminare l’infezione virale e impedire la replicazione dei virus [112, 113].
Nei pazienti con COVID-19, vengono rilevati alti livelli di IFN, specialmente IFN I; questa molecola blocca la replicazione virale nelle cellule adiacenti e produce alcuni effetti contro l’infezione virale come l’induzione dell’espressione genica stimolata dall’interferone, la stimolazione della produzione di citochine e l’attivazione delle cellule di risposta immunitaria (cioè macrofagi, monociti e neutrofili ) [112, 114]. Altre infezioni da CoV hanno una risposta simile [115, 116].
Inoltre, i pazienti infetti da SARS-CoV-2 hanno livelli aumentati di diverse citochine e chemochine: TNF-α, IFN-γ, antagonista del recettore dell’interleuchina-1 (IL-1RA), IL-2, IL-6, IL-7, IL-8, IL-9, IL-10 e il fattore stimolante le colonie dei granulociti macrofagi; cosa importante, alti livelli di IL-6 sono stati collegati a una prognosi peggiore nei pazienti con COVID-19 [116, 117].
Questa elevata produzione e squilibrio di tutte queste molecole è definita tempesta di citochine (CS), che potrebbe essere un fattore essenziale per causare l’interruzione del BBB [110]. È interessante notare che CS induce l’attivazione di piastrine, neutrofili, monociti e macrofagi; inoltre, alcune di queste molecole possono interagire con il complemento e i sistemi di coagulazione e contribuire all’infiammazione patogena [117].
Inoltre, alcune chemochine possono attrarre alcune cellule di risposta immunitaria innata come monociti, cellule natural killer, cellule dendritiche e cellule T [118] e indurre la produzione di altre citochine come la proteina chemiotattica dei monociti-1, il fattore stimolante le colonie di granulociti, la proteina infiammatoria dei macrofagi 1-α e IL-10 che reclutano linfociti e monociti e avviano la risposta umorale. Insieme, tutti questi meccanismi possono contribuire alla gravità dei sintomi neurologici dell’infezione da SARS-CoV-2 nel BBB [30].
Sono stati descritti anche altri disturbi fisiologici nei pazienti COVID-19 come trombocitopenia, linfopenia (CD8 + T, CD4 + T, cellule Treg e piastrine) ed eosinopenia. Campioni di sangue, milza e linfonodi presentano questi tipi di disregolazione dovuta alle cellule reclutate nei siti infetti per controllare la replicazione virale (Tabella (Tabella 3) 3) [119, 120]. Inoltre, diversi pazienti presentano livelli elevati di D-dimero nella fase iniziale dell’infezione (Tabella (Tabella 3) .3). Questa molecola è un importante marker nel disturbo della coagulazione. Rappresenta uno stato trombotico che porta a eventi vascolari embolici e può produrre coaguli venosi e indurre danni cerebrali [121]. Perciò,
Tabella 3
Risultati di laboratorio nella manifestazione neurologica in COVID-19
| Autore | Manifestazione | Rilevamento di laboratorio | Riferimento |
|---|---|---|---|
| Avula et al. | Ictus | – Linfopenia | [46, 48] |
| – C-reattivo elevato [26 mg / dl (0,04 mg / dl)] | |||
| – D-dimero elevato [media 8704 ng / ml (<880 ng / ml) | |||
| – Elevata lattato deidrogenasi (712 U / L) | |||
| Oxley et al. | – D-dimero [5972 ng / ml (0–500 ng / ml)] | ||
| Moringuchi et al. | Meningoencefalite | – Elevati neutrofili | [45] |
| – Aumento della proteina C reattiva | |||
| Guitierrez-Ortiz et al. | Sindrome di Guillain Barre | – Linfopenia (1000 cellule / μl) | [50, 51, 54] |
| – Leucopenia (3100 / cellule / μl) | |||
| – Proteina C reattiva elevata (2,8 mg / dl) | |||
| – Anticorpo ganglioside anti GD1b-IgG positivo | |||
| Virani et al. | – Linfopenia e trombocitopenia | ||
| Zhao et al. | – Linfocitopenia [0,52 × 10 9 / L (1,1–3,2 × 10 9 / L)] | ||
| – Trombocitopenia [113 × 10 9 / L (125–3000 × 10 9 / L)] |
Un meccanismo inesplorato che potrebbe produrre danni nel BBB è la risposta immunitaria adattativa. La generazione di anticorpi (Abs) contro SARS-CoV-2 può cross-reagire con alcune molecole delle cellule endoteliali microvascolari cerebrali e produrre danni attraverso l’attivazione del sistema del complemento (proteine C3 e C4).
Inoltre, il fenomeno di potenziamento dipendente dagli addominali può aumentare l’infezione e contribuire alla lesione. Questo processo è stato ampiamente studiato nell’infezione da virus Dengue e Zika, dove gli Abs prodotti nella prima esposizione possono reagire in modo crociato in una seconda esposizione e potenziare l’infezione invece di neutralizzarla [124, 125]. Inoltre, gli addominali possono generare un attacco autoimmune e interagire con il virus formando immunocomplessi e indurre l’attivazione del sistema del complemento [126].
Recentemente, alcuni studi hanno dimostrato che la risposta immunitaria cellulare potrebbe essere centrale per determinare la condizione della malattia. Diversi virus, incluso SARS-CoV-2, possono attivare CD4 + e CD8 + e indurre espansione clonale, effettori cellulari specifici e memoria cellulare [127, 128].
Inoltre, i linfociti T possono reagire in modo crociato inducendo uno stato di protezione osservato in persone non esposte con SARS-CoV-2 [129, 130]. Infine, devono essere chiariti ulteriori studi sull’interazione con SARS-CoV-2 e il sistema immunitario dell’ospite. La ricerca sui meccanismi coinvolti nella risposta all’infiammazione può consentire lo sviluppo di strategie che potrebbero aiutare a mitigare le conseguenze sulla salute di questa pandemia.
Implicazioni neurologiche della distruzione del BBB da SARS-CoV-2
Come discusso in precedenza, più virus respiratori possono influenzare il sistema nervoso centrale. Ad esempio, il virus dell’epatite del topo induce infiammazione, danno BBB e demielinizzazione nei modelli di ratto [131]. Allo stesso modo, un caso clinico di HCoV-OC43 rilevato in campioni di liquido spinale nasofaringeo e cerebrale di un paziente bambino ha mostrato encefalomielite disseminata acuta, una malattia del SNC a bassa prevalenza che induce demielinizzazione [132].
Il virus H1N1, l’agente eziologico di alti tassi di mortalità, presentava anche complicazioni neurologiche. Uno studio retrospettivo delle cartelle cliniche di 55 pazienti infetti da H1N1 ha rilevato il 50% dei sintomi neurologici visibili [133]. È interessante notare che la maggior parte dei pazienti con manifestazioni neurologiche dovute a infezione da H1N1 manifestavano edema cerebrale [134].
È importante sottolineare che negli studi autoptici, i pazienti con SARS-CoV hanno mostrato un’attivazione endoteliale associata alla perdita dell’integrità vascolare cerebrale con emorragia multifocale [135]. L’esame istologico di campioni di tessuto cerebrale di pazienti con infezione da SARS-CoV ha mostrato anche degenerazione neuronale, necrosi, edema, estesa iperplasia delle cellule gliali e infiltrazione cellulare delle pareti vascolari da parte di monociti e linfociti [40].
Con questo background, diversi studi hanno tentato di caratterizzare le manifestazioni neurologiche di SARS-CoV-2. È emerso un numero crescente di segnalazioni di casi individuali che descrivono disturbi neurologici acuti che vanno dalla sindrome di Guillain-Barré e mielite acuta all’encefalopatia necrotizzante emorragica acuta [136]. Sebbene le implicazioni neurologiche a lungo termine dell’infezione da SARS-CoV-2 siano ancora sconosciute, importanti indizi suggeriscono che le complicanze della malattia sono correlate all’invasione del SNC danneggiando il BBB.
Implicazioni neurologiche della distruzione della BBB da SARS-CoV-2 nella demenza a lungo termine
Esiste un’interazione molto complessa tra le cellule cerebrali e il sistema vascolare cerebrale. Di conseguenza, la conservazione della funzione cerebrovascolare e della sua integrità ha un ruolo centrale in questa sofisticata comunicazione. Inoltre, eventuali squilibri possono avere conseguenze acute e croniche deleterie come lo sviluppo di malattie neurodegenerative e demenza [137].
Si sospetta che gli agenti infettivi contribuiscano alla demenza, specialmente nella malattia di Alzheimer (AD). È interessante notare che l’interruzione del BBB sembra essere una caratteristica precoce di questa malattia [138]. Ad esempio, Bell et al. [139] hanno dimostrato che la degradazione del BBB era derivata dall’infiltrazione di proteine neurotossiche (p. Es., Peptidi β-amiloidi, il segno distintivo dell’AD), che colpivano i neuroni e che iniziarono o esacerbarono la neurodegenerazione.
Inoltre, Ueno et al. [140] utilizzando modelli animali sperimentali che mostrano alcuni fenotipi di demenza vascolare hanno mostrato che il danno BBB potrebbe essere correlato all’accumulo di peptidi β-amiloidi. Di conseguenza, il danno indotto al BBB da agenti infettivi potrebbe innescare malattie neurodegenerative in pazienti predisposti [141]. Pertanto, infezioni virali come SARS-CoV-2 potrebbero essere associate a un aumentato rischio di sviluppo di AD e a un tasso più rapido di declino cognitivo nelle popolazioni più anziane.
L’AD nell’infezione da virus sistemica è un esempio di una condizione che è principalmente neurodegenerativa; tuttavia, in molti casi, non è chiaro se i cambiamenti di BBB siano la causa o l’effetto di una neuropatologia. Inoltre, è possibile che le anomalie BBB e la malattia si guidino a vicenda in modo autoalimentato, contribuendo alla progressione del danno [142]. Come accennato, l’infiammazione sistemica acuta e cronica accelera il progresso dell’AD [143].
Inoltre, uno studio di follow-up di 5 anni ha mostrato che le infezioni virali, come quella indotta dal citomegalovirus, sono collegate a un declino cognitivo più rapido e allo sviluppo dell’AD [144]. Inoltre, l’infiammazione sistemica nell’AD è associata a diversi cambiamenti di BBB, che favoriscono ulteriormente l’accumulo di peptidi β-amiloidi nel cervello perché la lesione altera l’afflusso e l’efflusso dei peptidi [145]; di conseguenza, l’infiammazione sistemica accelera la deposizione di peptidi amiloide-β dell’ippocampo [146].
Pertanto, la disfunzione del BBB potrebbe svolgere un ruolo significativo nella patogenesi della demenza vascolare indotta dall’infezione da SARS-CoV-2, ma sono necessarie ulteriori osservazioni.
Inoltre, il danno al BBB non è l’unico meccanismo in cui l’infezione da SARS-CoV-2 può provocare demenza. Alcuni dati suggeriscono che la proteina β-amiloide possiede attività antimicrobica e antivirale in vitro [147]. Pertanto, la presenza di depositi insolubili di peptidi β-amiloidi potrebbe essere un fattore (p. Es., Predisposizione genetica) che altera la risposta all’infezione virale da SARS-CoV-2.
Pertanto, è concepibile che SARS-CoV-2 contribuisca al danno BBB e crei anche un effetto feed-forward per cui il danno indotto da patogeni favorisce un’ulteriore diffusione delle zone di transito del patogeno e persino lo sviluppo sequenziale della patologia associata all’AD [138 ].
D’altra parte, gli individui con AD sono più vulnerabili agli effetti dell’infezione periferica, in particolare SARS-CoV-2, principalmente a causa dell’associazione di comorbidità fisiche. È più probabile che questi individui abbiano malattie cardiovascolari, diabete e polmonite [148]. Inoltre, vi è una diminuzione complessiva della diversità delle cellule T ingenue dopo i 65 anni [149-151]; questo può limitare la capacità dell’individuo di indurre una risposta immunitaria sufficiente alle infezioni. Insieme, questi dati indicano la vulnerabilità di questi pazienti alle infezioni.
Implicazioni neurologiche dell’interruzione del BBB da SARS-CoV-2 nella sclerosi multipla
La sclerosi multipla (SM) è una malattia infiammatoria cronica del SNC, caratterizzata da diversi processi patologici, tra cui infiammazione, migrazione transendoteliale, demielinizzazione, assonopatia e perdita di neuroni mediata dalle cellule immunitarie [152]. La SM rappresenta una malattia neurologica in cui un agente infettivo gioca un ruolo scatenante, essendo i virus il colpevole più probabile in individui geneticamente predisposti [141].
Si presume che diversi virus neurotropi che utilizzano meccanismi simili possano essere coinvolti nella patogenesi della SM [153]. Alcuni virus che sono stati implicati nello sviluppo della SM includono i virus dell’herpes, i paramixovirus, i picornavirus, nonché i virus che colpiscono classicamente il sistema respiratorio come il virus dell’influenza [154].
Un passaggio critico nella patogenesi della SM è l’infiltrazione di linfociti T CD4 + autoreattivi nel SNC dopo l’attivazione nella periferia. Valutazione di citochine proinfiammatorie (IL-1β, IL-6 e IL-8), citochine Th1 (IFN-γ, TNF-α, IL-2 e IL-12) e citochine Th2 (IL-4, IL- 5 e IL-10) nei sieri raccolti da pazienti con SARS entro 2 giorni dal ricovero ospedaliero hanno mostrato un aumento sostanziale di IL-12, IL-6, IL-8, IL-10 e IFN-γ [155].
Inoltre, Sonar et al. [156] ha rivelato che l’IFN-γ ha favorito la migrazione transendoteliale dei linfociti T CD4 + dal lato apicale (lato luminale) al lato basale (lato abluminale) del monostrato endoteliale (Fig. (Fig.2) .2). Inoltre, utilizzando l’immunofluorescenza multicolore e l’analisi microscopica confocale, questi autori hanno indicato che l’IFN-γ induce la rilocalizzazione di ICAM-1, la molecola di adesione delle cellule endoteliali piastriniche-1, la proteina 1 della zona occludente e la VE-caderina nelle cellule endoteliali.
Questi risultati rivelano che l’IFN-γ prodotto durante la risposta all’infezione e all’infiammazione potrebbe contribuire all’interruzione del BBB e promuovere la migrazione cerebrale delle cellule T CD4 +. È interessante notare che l’interruzione del BBB appare nell’encefalomielite autoimmune sperimentale, un tipico modello di SM, e la gravità clinica è collegata al grado di integrità della BBB [157]. Inoltre, studi di imaging hanno mostrato la rottura della BBB nella sostanza bianca che appare normale nella SM [158].
Questi dati sono importanti poiché la rottura del BBB precede lo sviluppo di nuove lesioni da SM [159]. In sintesi, è possibile che il danno causato dall’infezione da SARS-CoV-2 alle cellule endoteliali causi anche la perdita dell’integrità del BBB, favorendo la progressione della SM.

Possibile meccanismo di danno alla barriera emato-encefalica (BBB) per azione di SARS-CoV-2. a L’espressione dell’enzima di conversione dell’angiotensina 2 (ACE2) e della pro-proteina convertasi furina (PCF) nella membrana delle cellule endoteliali microvascolari cerebrali facilita l’infezione da SARS-CoV-2. b L’infezione da SARS-CoV2 attiva le cellule microendoteliali cerebrali inducendo un’elevata espressione delle molecole di adesione vascolare e intercellulare (VCAM e ICAM). Allo stesso modo, SARS-CoV-2 induce l’espressione e l’attivazione delle metalloproteinasi della matrice (MMP) che degradano le proteine delle giunzioni strette. c Il riconoscimento di ICAM e ICAM attraverso le integrine β1 e β2 provoca il legame dei leucociti circolanti alle cellule endoteliali che portano allo stravaso transcellulare. Questo processo facilita l’ingresso virale nel parenchima cerebrale attraverso il meccanismo del “cavallo di Troia”. d La replicazione virale di SARS-CoV-2 induce la contrazione e la lisi delle cellule endoteliali. La maggiore permeabilità del BBB consente lo stravaso delle proteine plasmatiche e delle cellule del sangue. L’attivazione dei leucociti e delle piastrine contribuisce al danno BBB. Inoltre, la morte delle cellule endoteliali disturba il microambiente del parenchima cerebrale consentendo il libero passaggio del virus SARS-CoV-2 e l’infezione di altre cellule del sistema nervoso centrale
link di riferimento: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7518400/
Maggiori informazioni: Myoung-Hwa Lee et al, Microvascular Injury in the Brains of Patients with Covid-19, New England Journal of Medicine (2020). DOI: 10.1056 / NEJMc2033369



{kind=link}