










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Since these deletions happen in a part of the sequence that encodes for the shape of the spike protein, the formerly neutralizing antibody can’t grab hold of the virus, the researchers report today in Science.
And because the molecular “proofreader” that usually catches errors during SARS-CoV-2 replication is “blind” to fixing deletions, they become cemented into the variant’s genetic material.
“You can’t fix what’s not there,” said study senior author Paul Duprex, Ph.D., director of the Center for Vaccine Research at the University of Pittsburgh. “Once it’s gone, it’s gone, and if it’s gone in an important part of the virus that the antibody ‘sees,’ then it’s gone for good.”
Ever since the paper was first submitted as a preprint in November, the researchers watched this pattern play out, as several variants of concern rapidly spread across the globe. The variants first identified in the United Kingdom and South Africa have these sequence deletions.
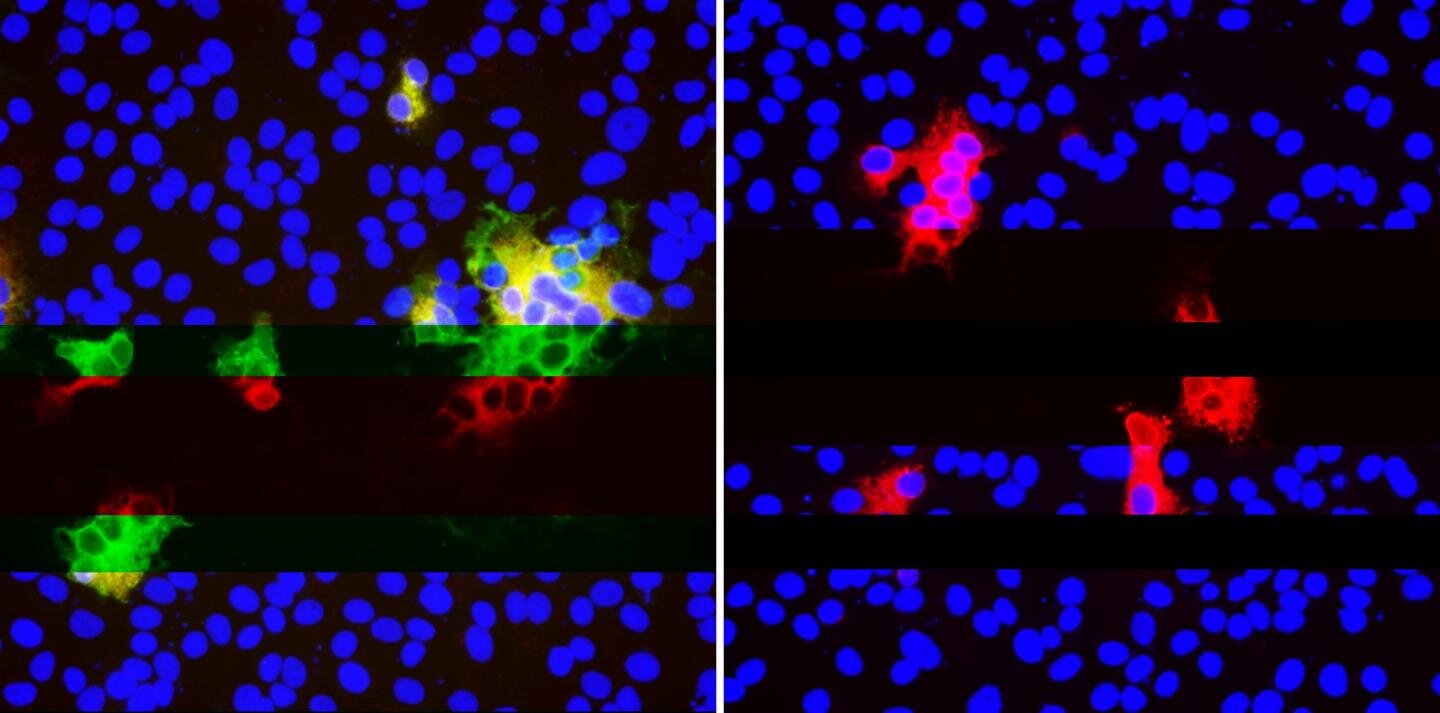
Duprex’s group first came across these neutralization-resistant deletions in a sample from an immunocompromised patient, who was infected with SARS-CoV-2 for 74 days before ultimately dying from COVID-19. That’s a long time for the virus and immune system to play “cat and mouse,” and gives ample opportunity to initiate the coevolutionary dance that results in these worrisome mutations in the viral genome that are occurring all over the world.
Then, Duprex enlisted the help of lead author Kevin McCarthy, Ph.D., assistant professor of molecular biology and molecular genetics at Pitt and an expert on influenza virus – a master of immune evasion – to see whether the deletions present in the viral sequences of this one patient might be part of a larger trend.
McCarthy and colleagues pored through the database of SARS-CoV-2 sequences collected across the world since the virus first spilled over into humans.

When the project started, in the summer of 2020, SARS-CoV-2 was thought to be relatively stable, but the more McCarthy scrutinized the database, the more deletions he saw, and a pattern emerged. The deletions kept happening in the same spots in the sequence, spots where the virus can tolerate a change in shape without losing its ability to invade cells and make copies of itself.
“Evolution was repeating itself,” said McCarthy, who recently started up a structural virology lab at Pitt’s Center for Vaccine Research. “By looking at this pattern, we could forecast. If it happened a few times, it was likely to happen again.”
Among the sequences McCarthy identified as having these deletions was the so-called “U.K. variant” – or to use its proper name, B.1.1.7. By this point, it was October 2020, and B.1.1.7 hadn’t taken off yet. In fact, it didn’t even have a name, but it was there in the datasets.
The strain was still emerging, and no one knew then the significance that it would come to have. But McCarthy’s analysis caught it in advance by looking for patterns in the genetic sequence.
Reassuringly, the strain identified in this Pittsburgh patient is still susceptible to neutralization by the swarm of antibodies present in convalescent plasma, demonstrating that mutational escape isn’t all or nothing. And that’s important to realize when it comes to designing tools to combat the virus.
“Going after the virus in multiple different ways is how we beat the shapeshifter,” Duprex said. “Combinations of different antibodies, combinations of nanobodies with antibodies, different types of vaccines. If there’s a crisis, we’ll want to have those backups.”
Although this paper shows how SARS-CoV-2 is likely to escape the existing vaccines and therapeutics, it’s impossible to know at this point exactly when that might happen.
“How far these deletions erode protection is yet to be determined,” McCarthy said. “At some point, we’re going to have to start reformulating vaccines, or at least entertain that idea.”

(A) Top. Sequences of viruses isolated from PLTI1 (PT) and viruses from patients with deletions in the same NTD region. Chromatograms are shown for sequences from PLTI1, which include sequencing of bulk reverse transcription products (CON) and individual cDNA clones. Bottom. Sequences from other long term infections from individuals AM (18) MA-JL (MA) (19) and a MSK cohort (M) with individuals 2, 3, 4, 6, 8, 11, 13 (13) Letters (A and B) designate different variants from the same patient. (B) Sequences of viruses from two patients with deletions in a different region of the NTD. All sequences are aligned to reference sequence (REF) MN985325 (WA-1). Genetic analysis of patient isolates is in fig. S1.
The primary target of SARS-CoV-2 infection is the lower respiratory tract causing flu-like illness with symptoms such as cough, fever, fatigue and arthralgia. However, the presentation and the course of the disease can range from asymptomatic to mild respiratory infections and pneumonia.
Some infected patients develop more severe disease with acute respiratory syndrome distress (ARD) about 7–10 days after onset of symptoms, following a rapid viral replication, increased pro-inflammatory cytokine production, “cytokine storm”, as well as chemokine responses and inflammatory cell infiltrates [[3], [4], [5]].
Among other risk factors identified, age has shown to be an important factor for the development of a more severe disease. Younger individuals often are asymptomatic or present mild symptoms and thus might have a crucial role in the spread of the disease [6,7].
During viral infection both innate and adaptive immune responses play a role in the pathogenesis of SARS-CoV-2. Specific motifs present in some SARS-CoV-2 protein structure or mediators released by infected damaged cells are recognized by the conserved innate pattern recognition receptors (PRRs) and seems to be involved in the modulation of the immune response.
An important feature in the pathophisiology of SARS-CoV-2 is the overproduction of early pro-inflammatory cytokines such as tumor necrosis factor (TNF), IL-6 and IL-1β which may lead to increased risk of vascular hyperpermeability, multi organ failure and eventually death if the high concentrations of cytokines is not controlled [[8], [9], [10]].
Moreover, it has been observed that adults with COVID-19 often present a decrease in both CD4+ and CD8 + T-cell subsets at the early stage of the disease what could contribute to virus replication and disease severity in some patients [11].
Innate immunity is important to inhibit viral replication and clearance, as well as to induce tissue repair and a prolonged immune response [12]. In this context, type I interferons (IFN-I, IFNα, IFNβ) play an important role in conferring antiviral activity in host cells.
However, it seems that SARS-CoV-2 have evolved mechanisms to evade IFN antiviral activity [13]. Nevertheless, given the complexity of COVID-19 pathophysiology, type I interferons may have different roles at different stages of infection or at mild versus severe COVID-19 patients [14]. Despite these findings, the immunity profile in COVID-19 is not completely understood.
Even before COVID-19 was declared a pandemic on March 11th, 2020 [15]; efforts trying to develop a vaccine against SARS-CoV-2 had been initiated, when the first viral genomic sequence became available in early January. However, the recognition of the pandemic intensified and induced a rush for the development of vaccines in different countries.
Thus, considering the rapid global spread of SARS-CoV-2 infection and the increasing death toll, development of an effective vaccine became priority. As result, great advances in a shorter time than expected for this research field was conquered, and several vaccine candidates are currently in phase II/III in China, UK, USA, Russia, Brazil and other countries [16].
Vaccines have played an important role in public health for decades to help prevent diseases like mumps, polio, rubella and yellow fever. Yet, we still have not understood well about their durabilities. Several questions have been raised regarding the novel vaccines being developed for SARS-CoV-2, specially concerned to the efficacy and durability.
This review will highlight important structure-function relationships of key SARS-CoV-2 proteins with focus on their role in pathogenesis and ability to elicit immune responses. We also will discuss the impact of new mutations in the genome of SARS-CoV-2 and how these changes can contribute for the emergence of new sub-strains with novel fitness and transmissibility ability, which could circumvent the efficacy and durability of the vaccines under development.
Mechanism of infection and SARS-CoV-2-induced human immune response
The first step in the infection by SARS-CoV-2 is the binding of the virus structural protein S to the host cell through its target receptor angiotensin-converting enzyme 2 (ACE2).
The glycoprotein S consists of two subunits: the subunit S1 contains a receptor-binding domain (RBD) that interacts with the ACE2 receptor while the S2 subunit mediates the fusion of viral and host cell membranes via formation of a six-helix bundle fusion core [40,41].
The serine protease TMPRSS2 also participates in this process by cleaving the S protein and allowing the fusion of its S2 subunit with cellular membrane [42]. It is believed that the formation of neutralizing antibodies targeting RDB-S1 or S2 region may block binding of protein S to ACE2 and prevent membrane fusion and entry of the virus into cells, inhibiting SARS-CoV-2 infection [43,44].
To better understand immunity towards the SARS-CoV-2 it is important to recall the basic concepts of the immune response. Innate immunity is the first line of defense against pathogens and comes from germline through both myeloid hematopoietic such as neutrophils, monocytes, and macrophages, and non-hematopoietic lymphoid such as natural killer [NK] and γδ T cells.
Adaptive immunity actions mainly via T cells and B cells characterized by their somatic genetic diversification of antigen-specific responses [45]. Acquired or adaptive immunity is established at the level of the individual, either through natural infection with a pathogen or through immunization with a vaccine [46].
A variety of clinical manifestations and different outcomes have been observed in patients with COVID-19 suggesting the complexity of the interaction of this new coronavirus with human host. It has been shown that immune response against SARS-CoV-2 involves different arms of the immune system including tissue barriers, innate and adaptive response as well as modulatory molecules mediators.
The infection and destruction of lung cells triggers a local immune response recruiting immune cells that release cytokines and prime adaptive T and B-cell responses. In most individuals this process is capable of resolving the infection. However, in some cases a dysfunctional immune response occurs, which triggers a cytokine storm that mediates widespread lung inflammation and leads to a severe form of COVID-19. This severe disease may lead to damage to other organs including the heart and brain [[47], [48], [49]].
After infection, the median incubation period of COVID-19 has been estimated approximately 4–5 days before symptoms onset [6,50,51]. However it has also been shown that around 97 % of symptomatic patients develop symptoms within 11.5 days and it includes fever, dry cough and less commonly, difficulty in breathing, muscle and/or joint pain, headache, dizziness, diarrhoea and nausea [[52], [53], [54], [55], [56], [57]].
Some individuals also experience temporary loss of taste (dysgeusia) and smell (anosmia) [[58], [59], [60]]. Within 5–6 days of symptoms onset, SARS-CoV-2 viral load reaches its peak [4,61]. Although majority of SARS-CoV-2-infected individuals are asymptomatic or have mild symptoms, severe COVID-19 patients can progress to acute respiratory distress syndromes (ARDs), which occurs on average around 8–9 days after symptom onset and is characterized by difficulty in breathing and decreased blood oxygen level [3,8,53].
ARDs are the cause of death in 70 % of the fatal cases of COVID-19 [8]. Studies with SARS-CoV have shown that infection reduces ACE2 expression in lung cells and this downregulation is considered to be an important factor in the COVID-19 pathophysiology [62,63].
The reason for this is related to ACE2 regulate the renin-angiotensin system (RAS) which is known to modulate blood pressure and fluid electrolyte balance. Therefore, a reduction of ACE2 function would enhance inflammation and vascular permeability in the airways [64].
Earlier studies have shown that the virus targets specially airway epithelial cells, alveolar epithelial cells, vascular endothelial cells and macrophages in the lung where ACE2 is expressed [[64], [65], [66], [67]].
After epithelial or other type of cells are infected, the replicating SARS-CoV-2 can cause cell lysis and promote direct damage to the tissue. Infected epithelial cells may present virus antigens to CD8 + T cells, which together with natural killers (NK) cells become cytotoxic to the virus-infected epithelial cells, leading them to perforin/granzyme induced apoptosis.
Dendrictic cells also can recognize antigens and present them to CD4 + T cells and induce their differentiation into memory Th1 and Th17 as well as memory T follicular helper (TFH) effector CD4 + T cells. Each cell subtype expresses different transcription factors, which regulate the function and cytokine secretion pattern of the cells and build the immune response.
The TFH cells can help B-cells to divide into plasma cells (PC) and synthetize IgM, IgA and IgG anti- SARS-CoV-2 specific antibodies. The diversity of antibody production during viral infection demonstrate the ability of adaptive immune response to try to overcome the obstacles imposed by the virus. Tissue macrophages can also mediate antigen presentation to CD4 + T-cells (Fig. 3 ).

Immune response against SAR-CoV-2 infection. 1. SARS-CoV-2 infects ACE2 expressing target cells such as alveolar epitelial type 2 cells in the lungs. 2. Virus may overcome induced antiviral Interferon (IFN) responses leading to uncontrolled replication. 3. Neutrophils and monocytes/macrophages are recruited to the site of infection and may cause overproduction of pro-inflammatory cytokines such as IL-6, IL-8, IL-12,TNF-α and others, involved in the immunopathology of COVID-19 in the lungs known as “cytokine storm”. Both humoral and cellular immune responses are elicited. 4. Infected epithelial cells may present virus antigens to CD8 + T cells, which together with natural killers (NK) cells become cytotoxic to the virus-infected epithelial cells leading to apoptosis. 5. Dendrictic cells (DC) present virus antigen to CD4 + T cells and induce their differentiation into memory Th1 and Th17 as well as memory T follicular helper (TFH) effector CD4 + T cells. 6. Activated B-cells and plasma cells synthesize IgM, IgA and IgG anti- SARS-CoV-2 specific antibodies. 7. Macrophages and dendrict cells present antigens to CD4 + T cells via MHC-TCR interaction. 8. Memory T cells subset are produced and may provide immunity against reinfection with the same virus strain for a period still not well established.
Noteworthy, severe lymphopenia and eosinopenia are often observed and related to a defect in antiviral and immune regulatory immunity. T lynphocytopenia has been inversely correlated with increased peripheral pro-inflammatory cytokines in COVID-19 patients [68,69]. Furthermore, low CD8 + T cell count has been considered a predictor for high mortality risk and illness severity [70].
These observations support the hypothesis that unbalanced adaptive immune responses can potentially induce detrimental effects on acute infected patients. Also, it has been suggested that SARS-CoV-2 may infect human T-cells via a novel route that involves the CD147 receptor, known as Basigin or EMMPRIN, expressed on the surface of T-lymphocytes [71,72]. Although still controversial, the interaction between SARS-CoV-2 and T cells could interfere with immunity. CD147 plays a role in differentiation, cell proliferation, migration, inflammation and apoptosis [73]. Thus, activation of downstream CD147 signaling pathway could lead to T-cell apoptosis or contribute to severity of COVID-19, however there is no study addressing this issue and a recent study found no evidence that the proposed CD147 can act as receptor for SARS-CoV-2 [74].
SARS-CoV-2- specific CD4+ and CD8 + T cells have shown to exhibit strongest response directed to the spike protein and produce effector and Th1 cytokines, such as interferon gama (IFN-γ) and tumor necrosis factor alpha (TNF-α), in addition to Th2 and Th17 cytokines, such as interleukin (IL)-4, IL-5, IL-13 and IL-17.
While Th1 cells are responsible for cell-mediated immune responses, Th2 are responsible for humoral-mediated immunity [75]. It has been shown that IL-17 can act on antigen-presenting cells (APCs) such as macrophages and dendrictic cells and induce cytokine and chemokine production [76]. SARS-CoV-2 T-cell responses have been detected not only to the spike protein S but also to M, N and other ORFs encoded proteins, although the response was more robust against spike protein, the main target of the most vaccines [77]. Thus, cytokine kinetics during COVID-19 has been crucial for understanding the fine balance between immunity and inflammation at different sites of infection.
Interestingly, reactive T cells was also detected in some health control patients not previously exposed to SARS-CoV-2, indicative of possible cross-reactivity due to past infection with coronaviruses probably from “common cold” [78]. SARS-CoV-2 seropositivity rates have been shown to vary from 2 to 73% in individuals who have probably not been exposed to the virus [79].
It is known that at least seven types of coronaviruses naturally infect humans. Besides SARS-CoV-2, severe acute respiratory syndrome coronavirus (SARS-CoV), and Middle East respiratory syndrome coronavirus (MERS-CoV) can cause severe acute respiratory illnesses. On the other hand, four endemic genotypes, such as 229E, NL63, OC43 and HKU1, usually cause mild upper respiratory tract infections, and thus can be classified as low-pathogenic human coronaviruses [80].
Therefore, considering the genetic relationship among different coronaviruses, it is not surprising that cross-reactivity may occur and be detected in serologic assays at different degrees and in different localities. Notwhithstanding, genetically attenuated virus can induce antibodies in animal models that neutralize both SARS-CoV and SARS-CoV-2 [17]. However, when spike protein is used as antigen, human sera seems to contain only antibodies that neutralize one type of virus [81].
Not less important, genetic susceptibility to infectious disease has a close relation to the major histocompatibility complex antigen loci (HLA) [82,83]. Antigen receptors on CD4+ or CD8 + T-cells recognize the conformational structure of the antigen-binding -grove together with the associated antigen peptides.
Thus, HLA molecules with increased binding specificities to SARS-CoV-2 virus peptides on the cell surface of presenting cells seems to be advantageous to individuals with determined HLA haplotype. Notwhistanding, host factors and population heterogeneity can contribute significantly to variability in cross reactivity and susceptibility to infection, specially due to the major histocompatibility complex antigen loci (MHC-HLA) [84] and possibly to polymorphisms in the gene ACE2 that encodes the receptor used for virus entry into host cells.
The latter has been subject of controversial debate and need more studies to reach a conclusion [85]. MHC class I HLA-B46:01 genotype was shown to have the fewest predicted binding peptides for SARS-CoV-2, suggesting that individuals with this allele may be particularly vulnerable to COVID-19, as they were previously shown to be for SARS. In contrast, HLAB15:03 showed the greatest capacity to present highly conserved SARS-CoV-2 peptides that are shared among common human coronaviruses, suggesting that it could enable cross-protective T-cell-based immunity [86].
COVID-19 severe patients experience symptoms where pneumonia, acute respiratory distress syndrome and cytokine storm become part of a complex immunopathological condition [6,87]. Development of neutralizing antiviral T-cell and antibody immunity is observed in the majority of patients with self-limiting viral respiratory disease at one week post infection. Immunoglobulin (Ig)M and IgA titers against SARS-CoV-2 can be detected in median 5 days after symptom onset while IgG titers, 14 days [88].
These antibodies have been used as a measurement to predict population exposure to SARS-CoV-2 as well as to determine the cross-reactivity with other coronaviruses [89]. Regarding the duration of anti-SARS-CoV-2 antibodies in individuals that have recovered from COVID-19 there are no conclusive response.
In one study, anti-SARS-CoV-2 IgG responses have been detected in most patients with either severe or mild disease, at 9 days after onset of infection and the antibody levels remained high throughout the study period which endured about 35–40 days [90]. On the other hand, titers of anti- SARS-CoV-2 IgG antibodies were found to decay in asymptomatic and early convalescent individuals after 90 days of onset of symptoms [91,92].
New studies have shown that IgA and IgM antibodies against RBD region of the spike protein were short-lived with median times to seroreversion of 71 and 49 days after symptom onset. Nevertheless, IgG antibodies decayed slowly through 90 days and these antibodies strongly correlated with anti S-neutralizing antibody titers [93]. Additionally, another study reported detection of IgA, IgM and IgG anti -SARS-CoV-2 not only in serum but also in the saliva of acute and convalescent patients and IgG could be detected for up to 3 months [94].
Interestingly, it has been reported cases of people presenting positivity for SARS-CoV-2 in molecular tests without detectable levels of protective IgG antibodies. Moreover, a recently published work analysed the antibody levels in 254 patients who had COVID-19, during a period of five months and demonstrated that SARS-CoV-2 IgG antibodies progressively decreased in outpatient and asymptomatic patients during observation up to five months post infection [95].
Together, these results indicate that acquired immunity by people with COVID-19 may not be lasting suggesting concerns about the long term efficacy of potential vaccines being developed. On the other hand, the apparent controversy on the durability of IgG could possibly be explained by variations in the methodology employed in different studies or be indicative of a more relevant role of cellular protective immune response.
The most common technique used to measure anti–SARS-CoV-2 antibody titers is an enzyme-linked immunosorbent assay (ELISA). While some authors measured anti spike protein receptor-binding domain (RBD) IgG antibodies, others measured antibodies anti whole spike protein or nucleocapsid from SARS-CoV-2 [96].
As mentioned before, spike protein is present in a variety of coronaviruses and may be an important factor to be considered in terms of cross-reactivity and interpretation of kinetics of antibodies against SARS-CoV-2 infections. Also, some cases of putative reinfection have been recently reported suggesting the possibility of waning immunity and arguing in favor of a decay in antibody titers overtime.
Although reinfection is currently subject of intense debate, a number of studies has been reporting cases of a second episode of positive SARS-CoV-2 infection after recovery [97]. In some cases, the patient follow up was not well conducted and the appropriate control tests were not performed, casting doubt about a true reinfection and suggesting only a persistent virus shedding [98].
A series of six cases reported in Brazil, suggest the possibility of reinfection but the results could also be due to viral reactivation [99]. On the other hand, a second episode of COVID-19 was reported in an asymptomatic patient from Hong Kong following a first symptomatic episode. SARS-CoV-2 whole genome sequencing was performed directly on respiratory specimens collected during the two episodes of COVID-19.
Interestingly, viral genomes from first and second episodes were shown to belong to different clades/lineages [100]. If the virus can be reactivated after a while following recovery from a infection is an open question. Therefore, the possibility of true SARS-CoV-2 reinfection remains unclear and further studies are required.
Although it is not clear the mechanisms by which SARS-CoV-2 can subvert the body’s innate antiviral cytokine responses in some patients, it is known by studies with other coronaviruses, that multiple viral structural and non-structural proteins can antagonize interferon responses. Some identified mechanisms include preventing recognition of virus RNA, preventing downstream interferon signaling pathway or inducing host mRNA degradation and inhibiting host protein translation [[101], [102], [103]].
On the other hand, dendritic cells play an important role in immune responses by their plasticity and unique ability to induce naïve T cell activation, coordinate and regulate adaptive immune responses. They are a heterogeneous group of cells that act as the strongest antigen-presenting cells (APCs) and effectively stimulate the activation of B and T lymphocytes, thus combining innate and adaptive immunity.
Notwithstanding, it has been shown that dendritic cells were significantly reduced or showed functional impairement in acute COVID-19 patients [104]. Dendritic cells are targeted through the interaction of virus S protein with dendritic cell –specific intercellular adhesion molecule 3–grabbing non-integrin (DC-SIGN) [105].
This adhesion molecule was initially discovered as an attachment factor for HIV virus and by interacting with viral glycoproteins augments infection [106]. DC-SIGN and the related protein DCSIGNR (also termed L-SIGN) have been shown to enhance infection in a variety of viruses including SARS-COV [107,108]. By targeting the C-type lectin DC-SIGN, viruses can subvert dendritic cell functions to scape immune surveillance [109].
Viral proteins such as “spike” are exposed to adaptive immune response and are the main targets of host antibodies [110]. The mechanisms underlying the generation of specific antibodies directed to a virus infection involve the production of antigenic peptides from viral proteins by the immune system B-cells.
These peptides bind to the major histocompatibility complex (MHC) and then are presented at the cell surface to other cell subsets. The naïve B-cells can also become stimulated by another type of cell named helper-T-cell and become a plasma cell able to produce antibodies. The diversity of antibodies generated by different stimulated B-cells have different binding epitope and protein sequences in a specific region of the antibody which may confer different degrees of affinity to antigens.
Nevertheless, viruses in a broad sense, have developed mechanisms to escape immune responses through rapid evolution of antibody-targeting epitopes, steric shielding of epitopes by glycan post-translational modifications, immune decoys such as soluble antigens that share viral spike epitopes, and immunosuppression to evade host recognition upon cellular entry [110].
Surface proteins outside of the virus structure are generally selected for antigens aiming therapeutic use, so that antibodies generated from a vaccine-primed B-cell can bind to the virus for neutralization. However, strain-specific antigens introduced as vaccines may have its use limited if variability somehow alter the antigen protein structure that may act as a mechanism of scape from the immune recognition [111].
Some of the stimulated B and T lymphocytes may become memory cells and persist for months or years in the body, allowing the mounting of faster and stronger responses in case of a new virus infection. Trained immunity is a term used for immunological memory and is thought to affect and prevent spread of virus infection. This property of lymphocytes is the basis of vaccine efficacy against specific infections [112].
Development of immunological memory and persistence of virus recognition memory responses are key for the long term-protection from SARS-CoV-2 reinfection and the durability of potentially successful vaccines. The humoral and cellular memory responses was analyzed in 15 COVID-19 recovered individuals who presented mild symptoms. Sustained neutralizing IgG antibodies and memory B-cells as well as SARS-CoV-2 specific memory T-cell were detectable up to 3 months and more than 3 months post symptom onset respectively [113,114].
The molecular mechanisms underlying long term reprogramming of immune cells are epigenetic in nature. To reach the memory status, the previously challenged cell needs to access regions of the genome that contain the target sequences and the regulatory elements of the genes involved in these processes. This process is regulated through durable epigenetic modifications, which allow unfolding of the chromatin and accessibility of transcription factors to the promoter and enhancer regions of the involved immune-related genes [115].
reference link: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7802525/
More information: “Recurrent deletions in the SARS-CoV-2 spike glycoprotein drive antibody escape,” by Kevin R. McCarthy et al., Science (2021). science.sciencemag.org/cgi/doi … 1126/science.abf6950



{kind=link}