









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



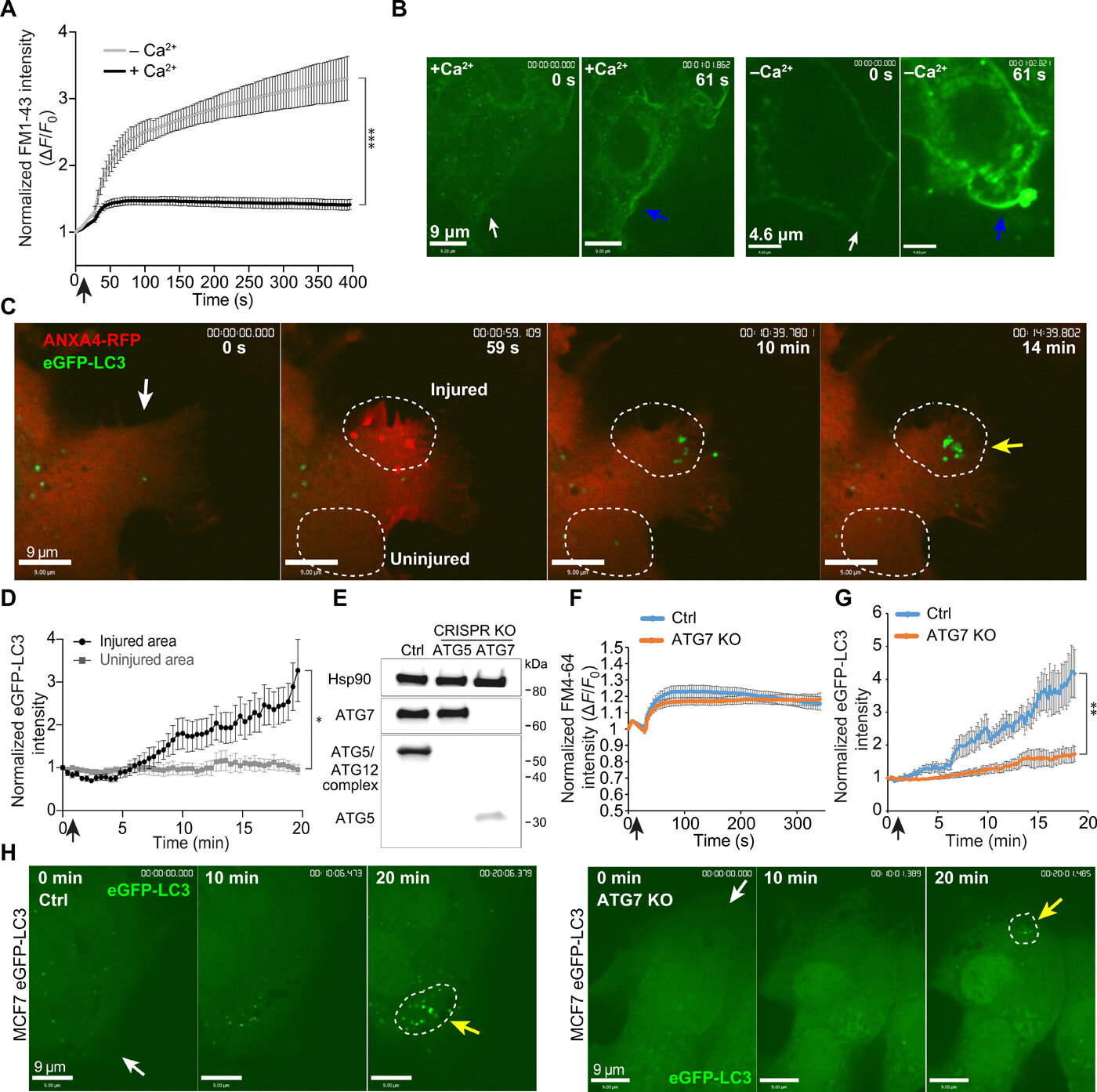
It is the membrane of cancer cells that is at the focus of the new research now showing a completely new way in which cancer cells can repair the damage that can otherwise kill them.
In both normal cells and cancer cells, the cell membrane acts as the skin of the cells. And damage to the membrane can be life threatening. The interior of cells is fluid, and if a hole is made in the membrane, the cell simply floats out and dies—a bit like a hole in a water balloon.
Therefore, damage to the cell membrane must be repaired quickly, and now research from a team of Danish researchers shows that cancer cells use a technique called macropinocytosis. The technique, which is already a known tool for cells in other contexts, consists in the cancer cells pulling the intact cell membrane in over the damaged area and sealing the hole in a matter of minutes. Next, the damaged part of the cell membrane is separated into small spheres and transported to the cells’ lysosomes, where they are broken down.
In the laboratory, the researchers damaged the membrane of the cancer cells using a laser that shoots small holes in the membrane and triggers macropinocytosis. Here, they can see that if the process is inhibited with substances blocking the formation of the small membrane spheres, the cancer cell can no longer repair the damage and dies.
“Our research provides very basic knowledge about how cancer cells survive. In our experiments, we have also shown that cancer cells die if the process is inhibited, and this points towards macropinocytosis as a target for future treatment.
It is a long-term perspective, but it is interesting,” says group leader Jesper Nylandsted from the Danish Cancer Society’s Research Center and the University of Copenhagen, who has headed the new research and who for many years has investigated how cancer cells repair their membranes.
Possibility of recycling
One of the most dangerous properties of cancer is when the disease spreads in the body. If tumors occur in new parts of the body, the disease becomes more difficult to treat and typically requires more extensive forms of treatment. It is also when cancer cells spread through the body’s tissues that they are particularly prone to damage to their membrane.
Researchers at the Danish Cancer Society have previously shown how cancer cells can use another technique to repair the membrane, namely by tying off the damaged part, rather like when a lizard throws its tail.
However, the experiments in the laboratory could indicate that especially the aggressive cancer cells use macropinocytosis. This may be due to the fact that the cancer cell has the opportunity to reuse the damaged membrane when it is degraded in the lysosomes. This type of recycling will be useful for cancer cells because they divide frequently, requiring large amounts of energy and material for the new cells.
And although the researchers have now published the new results, their work is not over. This is explained by another member of the research team, postdoc Stine Lauritzen Sønder: “We continue to work and investigate how cancer cells protect their membranes.
In connection with macropinocytosis in particular, it is also interesting to see what happens after the membrane is closed. We believe that the first patching is a bit rough and that a more thorough repair of the membrane is needed afterwards. It can be another weak point in the cancer cells, and is something we want to examine closer,” she says.
Cancer macropinocytosis is an endocytic uptake process by which cancer cells internalize extracellular proteins or necrotic cell debris and deliver them to lysosomes for further degradation [1-4]. The decomposition of these macropinocytic cargos contributes to the supply of the desperately needed amino acids that support cancer cells’ survival and growth.
As shown in Figure 1, the molecular mechanism of macropinocytosis in cancer cells is quite complicated. The Ras and PI3K signaling pathways are the most common in cancer macropinocytosis (Table 1). The macropinocytic induction is associated with the activation of oncogenes (e.g., RAS or EGFR) or deactivation of tumor suppressor genes (e.g., PTEN) in cancer cells [5-7].
Macropinocytosis is closely related to actin cytoskeleton remodeling and the macropinosomes’ generation process includes plasma membrane ruffling, macropinocytic cup formation and closure, and detachment from the plasma membrane. Several pivotal regulators of actin polymerization, such as small GTPases (e.g., Ras, Rac, Cdc42, Arf6, and Rab5), p21-activated kinase 1 (Pak1), and phosphoinositide 3-kinase (PI3K), have been linked to the formation of plasma membrane protrusions and macropinocytic activity [8].
Interestingly, RAS and PI3K activation is related to the stimulation of receptor tyrosine kinases (RTK), such as platelet-derived growth factor receptor (PDGFR) and epidermal growth factor receptor (EGFR) [1,9]. Certainly, there are also negative-regulatory factors that weaken macropinocytosis. For instance, the PTEN phosphatase blocks the PI3K signaling pathway by converting PI (3, 4, 5) P3 to PI (4, 5) P2.
Importantly, PI (3, 4, 5) P3 is necessary for plasma membranes’ ruffling and macropinosomes’ closures [7]. In addition, one particular study demonstrated that mTORC1 could suppress the lysosomal catabolism of extracellular proteins [2]. Of course, there are also other specific pathways, such as the Wnt signaling pathway, that play an important role in micropinocytosis in colorectal and bladder cancers.
Therefore, with the identification of the molecular mechanism of macropinocytosis, novel therapeutic approaches can be developed to treat cancers. At present, the major therapeutic methods in targeting macropinocytosis include small molecules and antibodies. Intriguingly, nucleic acids, as an emerging therapeutic drug, might become the third modality in cancer treatments using macropinocytosis [10].
Here, we aim to summarize the role of macropinocytosis in different type of cancers, including pancreatic, lung, colorectal, bladder, prostate, brain and nervous system cancers. Meanwhile, we discuss three major therapeutic modalities, including chemotherapy, immunotherapy, and nucleic acid therapy that are used for targeting macropinocytosis in cancers.

Schematic diagram of cancer cells ingesting extracellular small particles, such as protein, necrotic debris, and ATP, through macropinocytosis. The activation of Ras and PI3K pathways, by oncogenic mutations, integrin αvβ3, EGFR or PDGFR, activates upstream effectors (e.g., Ras and PI3K) which then activate downstream effectors, such as Rac1, Cdc42, and Pak1, that are significant regulators of macropinocytosis. PTEN loss can activate PI3K, which is closely linked to membrane phospholipid conversion. AMPK activation can activate Rac1, which can trigger macropinocytosis, and prompt the transport of transcription factors into the nucleus, resulting in elevated expression of lysosomal genes in the nucleus. The activation of Rac1 and Cdc42 and the lysosomal degradation of macropinocytic cargos are sensitive to pH changes, which are regulated by the Na+/H+ exchanger (NHE) and the vacuolar H+-ATPase (v-ATPase). In the lysosome, extracellular proteins, or necrotic debris, can be degraded into amino acids, which can fuel the TCA cycle, leading to increased cell growth and survival. The lysosomal degradation process of the macropinocytosed protein into amino acids can be inhibited by mTORC1. Interestingly, AMPK can antagonize the mTORC1 pathway and improve the degradation efficiency of internalized proteins in the lysosome.
Table 1
Example of micropinocytosis in different cancer types
| Type of cancer | Molecular driver | Macropinocytic cargo | Signaling Pathway | Reference |
|---|---|---|---|---|
| pancreatic cancer | KRAS | proteins | RAS, PI3K | [1,9] |
| lung cancer | KRAS | proteins, ATP | RAS, PI3K, Rac | [26,28-31] |
| galectin-3 | ||||
| integrin αvβ3 | ||||
| AMPK | ||||
| colorectal cancer | KRAS | proteins | RAS, PI3K, Wnt | [39,42,43] |
| Fz | ||||
| Lrp6 | ||||
| APC | ||||
| PRMT1 | ||||
| bladder cancer | HRAS | proteins | RAS, PI3K, Wnt | [46,48] |
| KRAS | ||||
| PTEN | ||||
| brain and nervous system cancers | ||||
| GBM | HRAS | proteins | RAS, PI3K | [49] |
| KRAS | ||||
| neuroblastoma | IGF-1 | proteins | PI3K | [57] |
| PMA | proteins | ? | [57] | |
| medulloblastoma | TrkA | proteins | ? | [58] |
| prostate cancer | PTEN | necrotic cell debris | PI3K | [7,27,61-63] |
Therapeutic modalities for exploiting cancers macropinocytosis
Chemotherapy
Since cancer cells can absorb extracellular small molecules, such as serum albumin, necrotic debris and ATP, through macropinocytosis, small molecule drugs can therefore be used to block the macropinocytosis pathway and treat cancers by disrupting the metabolic activity of cancer cells [69-72]. For example, numerous studies have shown that 5-(N-ethyl-N-propyl) amiloride (EIPA) can inhibit both macropinocytosis and actin polymerization by blocking NHE (Table 2) [1,42,73].
Similarly, v-ATPase inhibitors, such as bafilomycin A1, that have an impact on lysosomal acidic pH, can also result in the inhibition of macropinocytosis and reduction of intracellular amino acid levels (Table 2) [74]. In addition, inhibitors of the signaling pathway networks that mediate the activity of macropinocytosis are also effectively used in anti-cancer treatment. For instance, a recent study indicated that the EGFR inhibitor, gefitinib, can suppress the macropinocytosis pathway in NSCLC cells (Table 2) [75].
Analogously, one particular study has demonstrated that the Galectin-3 inhibitor, GCS-100, could disrupt the interaction between Galectin-3 and integrin αvβ3, thereby inhibiting the macropinocytosis pathway in KRAS-addicted lung and pancreatic cancer cells (Table 2) [28]. In fact, DOCK1 is a Rac-specific guanine nucleotide exchange factor (GEF) and the selective inhibitor of DOCK1,1-(2-(3’-(trifluoromethyl)-[1,1’-biphenyl]-4-yl)-2-oxoethyl)-5-pyrrolidinylsulfonyl-2(1H)-pyridone (TBOPP), can repress DOCK1-mediated macropinocytosis in RAS-transformed cancer cells (Table 2) [76].
There are other frequently used small molecule inhibitors of macropinocytosis, such as actin inhibitors (blebbstatin [77], cytochalasin D [78]) and PI3K inhibitors (Wortmannin, LY294002 [79]) (Table 2). Moreover, recent studies reported that a novel lysosomal inhibitor, palmitoyl-protein thioesterase 1 (PPT1), can suppress lysosomal activity that play a critical role in degrading proteins during macropinocytosis (Table 2) [80,81]. Interestingly, it was shown that better effects could be obtained when mTOR inhibitors are delivered in combination with inhibitors that could suppress the uptake or the process of lysosomal extracellular proteins scavenging [2,23].
In addition to blocking cancer cell macropinocytosis, another anti-cancer method is to cause a significant increase in cancer cell catastrophic macropinocytosis by using related inhibitors or inducers. For example, one particular study has demonstrated that treatment with the CK2 inhibitor, silmitasertib, and gives rise to large number of vacuoles that derived from massive macropinocytosis, resulting in colorectal cancer cells methuosis-like death (Table 2; Figure 2) [39]. Similarly, several studies have shown that inducers can also cause massive macropinocytosis that lead to methuosis in brain and nervous system cancers cells (Table 2) [55,56,58,59].
Lastly, researchers also made great efforts to exploit macropinocytosis for delivering anti-cancer agents into cancer cells [71]. For example, a study demonstrated that albumin-conjugated doxorubicin (DOX) could be internalized into KRAS-driven PDAC cells by macropinocytosis, thereby releasing DOX in the PDAC cells to exert its toxic effect (Table 2) [82]. Similarly, it has been shown that macropinocytosis can be exploited for internalization of drug that combined albumin nanoparticles (nab-paclitaxel) with gemcitabine for PDAC therapy (Table 2) [83].
Interestingly, a recent research indicated that triptolide prodrug-loaded UPSM (T-UPSM) could be absorbed into KRAS-mutant PDAC cells through macropinocytosis, which pH triggered the rapid release of drug in lysosomes (Table 2) [84]. Furthermore, Tubeimoside-1 (TBM1) can induce macropinocytosis and increase 5-FU transport into colorectal cancer cells where it promotes synergistic anti-cancer effects (Table 2) [85]. Analogously, a recent study has shown that MOMIPP could also induce massive macropinocytosis, leading to increased uptake of temozolomide by GBM cells and resulting in additional anti-cancer effects (Table 2) [49]. Therefore, understanding the mechanism of cancer cells’ macropinocytosis is very helpful for cancer treatment.
Immunotherapy
Currently, therapeutic methods, such as monoclonal antibodies (mAbs) and vaccines, are widely used in cancer immunotherapy [86]. Interestingly, recent studies have elaborated that mAbs could be internalized into cancer cells through macropinocytosis [87,88]. In addition, macropinocytosis can be selectively upregulated in different types of cancer. Therefore, mAbs that can be effectively internalized via macropinocytosis are meaningful for the development of cancer therapies.
For instance, bevacizumab nanoparticles, which can be internalized by macropinocytosis, were applied to target intracellular VEGF in NSCLC (Table 2) [89]. In recent years, more studies have reported on the application of antibody-drug conjugates (ADCs) in cancer therapy [90-92]. Intriguingly, one particular study has indicated that the ScFv antibody that was based on albumin domain and its cytotoxic conjugate, exhibits characteristics of targeted-EGFR, intensive-macropinocytosis and cytotoxicity, resulting in apparent growth inhibition of KRAS-mutant pancreatic cancer (Table 2) [93].
In addition to mAbs, vaccines that are associated with macropinocytosis also play an important role in cancer immunotherapy. For example, one particular study has shown that oncogenic activation of macropinocytosis resulted in BCG internalization into bladder cancer cells (Table 2) [46]. Additionally, mycobacterium tuberculosis vaccine (MTBVAC) could also be internalized into bladder cancer cells through macropinocytosis, resulting in the inhibition of cancer cells’ growth (Table 2) [94].
Thus, MTBVAC can be used as a new immunotherapy drug for bladder cancer. Similarly, ApoE3-incorporated biomimetic nanoparticle is also very likely to be used as a safe and effective nanovaccine for cancer immunotherapy by exploiting the macropinocytosis pathway (Table 2) [95]. Briefly, both mAbs and vaccines can be effectively applied to cancer immunotherapy through exploiting macropinocytosis.
Nucleic acid therapy
Nucleic acid therapy may become another emerging therapeutic modality that could use the role of macropinocytosis in cancer cells [10]. Currently, the fastest clinically progressing nucleic acid drugs for cancer therapy are antisense oligonucleotides (ASOs), DNA, short interfering RNA (siRNA), microRNA (miRNA), and messenger RNA (mRNA) (Figure 4) [96]. However, due to the cell membrane barrier, these nucleic acids require carriers to enter the cell.
Fortunately, there are several delivery systems, such as lipids [97,98], polymers [99], and peptides [100], that can be used to carry nucleic acids into cancer cells and via macropinocytosis. For example, translationally controlled tumor protein (TCTP) ASOs that carried by lipids can be internalized into castration resistant prostate cancer cells through macropinocytosis, resulting in decreased expression of TCTP, which is tightly linked to cell growth (Table 2) [97].
Interestingly, siRNA, which is carried by lipofectamine, can drive the suppression of the transcription factor EB (TFEB) in KRAS-mutant cancer cells, leading to a substantial reduction in the lysosomal ability to degrade extracellular proteins (Table 2) [101]. Similarly, AGMA1 polyamidoamine that effectively carry siRNAs can be absorbed into prostate cancer cells by macropinocytosis and cause gene silencing without inducing cytotoxicity [102]. Furthermore, one particular study demonstrated that gold nanoparticles modified by cell-penetrating peptides (CPPs) can significantly increase cellular uptake and the absorption rate of nucleic acid drugs [103].

Nucleic acid drugs, including antisense oligonucleotide (ASO), DNA, short interfering RNA (siRNA), microRNA (miRNA), and messenger RNA (mRNA), can be applied to clinical cancer therapy. Carriers that contain nucleic acid drugs enter into cancer cells through macropinocytosis and are released from lysosomes. ASO is associated with the activity of RNase H endonuclease. MiRNAs and siRNAs are recognized by the RNA-induced silencing complex (RISC). Ribosomes and mRNAs participate in protein translation and expression. DNAs may be transported into the nucleus and affect the transcription of gene. Eventually, the expression of macropinocytosis-associated proteins will be reduced.
Interestingly, in addition to these carriers (e.g., lipids, polymers, and peptides), inorganic nanoparticles [53] and extracellular vesicles (EVs) [104-106] can also carry nucleic acids into cancer cells through macropinocytosis. For example, the nanostructure, lipoprotein, which carries the activating transcription factor-5 (ATF5) siRNA, can cross the blood-brain barrier, and be internalized by Ras-driven GBM cells via macropinocytosis. Subsequently, the release of ATF5 siRNA in the GBM cells causes the inhibition of cancer cell growth (Table 2) [53]. In addition, exosomes can carry siRNA that targets oncogenic KRASG12D, into pancreatic cancer cells through macropinocytosis, which inhibits cancer cell growth and increases overall survival (Table 2) [106]. Therefore, research on the use of macropinocytosis for intracellular delivery of nucleic acids drugs to cancer cells, is needed in the future.
reference link : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7840718/
More information: Stine Lauritzen Sønder et al, Restructuring of the plasma membrane upon damage by LC3-associated macropinocytosis, Science Advances (2021). DOI: 10.1126/sciadv.abg1969



{kind=link}