










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



In a study published in Nature Metabolism, the team reported that irisin, secreted by the muscles during exercise, could be an effective therapeutic for addressing deficits of the brain that result from Alzheimer’s disease.
“Preserving cognitive function is a major challenge in an increasingly aging population,” says Christiane Wrann, DVM, Ph.D., leader of the Program in Neuroprotection in Exercise at MGH and senior author of the study. “Exercise is known to have positive effects on brain health, which is why identifying key mediators of those neuroprotective benefits, like irisin, has become such a critical goal of research.”
At the same time, the MGH study found that elevating irisin levels in the bloodstream improved cognitive function and neuroinflammation in mouse models for Alzheimer’s disease.
“What makes this study particularly strong is that we show irisin’s effect on cognitive function in not one but four different mouse models,” states Bruce Spiegelman of Dana-Farber Cancer Institute and Harvard Medical School, who discovered irisin in 2012 and is a co-author of the current paper.
Researchers were further encouraged by the fact that irisin treatment was effective in Alzheimer’s disease mouse models even after the development of significant pathology.
Another important finding of the study is that irisin protects against neuroinflammation by acting directly on glia cells in the brain. Co-author Rudy Tanzi, co-director of the McCance Center for Brain Health at MGH, explains that “it’s hard to imagine anything better for brain health than daily exercise, and our findings shed new light on the mechanism involved: protecting against neuroinflammation, perhaps the biggest killer of brain neurons as we age.”
Wrann adds that “since irisin does not specifically target amyloid plaques, but rather neuroinflammation directly, we’re optimistic it could have beneficial effects on neurodegenerative diseases beyond just Alzheimer’s.”
The beneficial effects of exercise on the brain are well known. In general, exercise offers an effective way to improve cognitive function in all ages, particularly in the elderly, who are considered the most vulnerable to neurodegenerative disorders. In this regard, myokines, hormones secreted by muscle in response to exercise, have recently gained attention as beneficial mediators. Irisin is a novel exercise-induced myokine, that modulates several bodily processes, such as glucose homeostasis, and reduces systemic inflammation.
Irisin is cleaved from fibronectin type III domain containing 5 (FNDC5), a transmembrane precursor protein expressed in muscle under the control of peroxisome proliferator-activated receptor-γ coactivator-1α (PGC-1α). The FNDC5/irisin system is also expressed in the hippocampus, where it stimulates the expression of the neurotrophin brain-derived neurotrophic factor in this area that is associated with learning and memory. In this review, we aimed to discuss the role of irisin as a key mediator of the beneficial effects of exercise on synaptic plasticity and memory in the elderly, suggesting its roles within the main promoters of the beneficial effects of exercise on the brain.
. . . .
Physical activity refers to body movement that is produced by the contraction of Skeletal Muscle (SkM) and that increases energy expenditure. It includes activities in the workplace (e.g., typing), around the house (e.g., household chores, such as cleaning) and during leisure time (e.g., walking, swimming, dancing, cycling).ùSeveral studies showed a relationship between physical and mental health and Physical Exercise (PE).
PE is known to slow down the process of age-relatedneurodegeneration, and existing hypotheses suggest that PE represents a potential adjunctive treatment for cognitive impairment. Regular physical activity can modulate the potential risk factors of dementia and other neurodegenerative disorders, such as Alzheimer’s disease (AD) and/or Parkinson disease (PD) [1].
Recently, a meta-analysis considered evidence on the safety and efficacy of PE as an additional therapeutic intervention for the quality of life, cognition, and depressive symptoms across several chronic brain disorders. Silverman and Deuster (2014) suggested that regular physical activity affects the following biological pathways:
(i) optimization of neuroendocrine and physiological responses to psychosocial and physical stressors;
(ii) action as a buffer against stress and stress-related diseases/chronic diseases;
(iii) promotion of an anti-inflammatory state; and (iv) enhancement of neuroplasticity and growth factor expression [2].
PE exerts these effects by influencing various molecular pathways and myokines; through autocrine, paracrine, and endocrine mechanisms [3,4,5,6]. In this regard, myokines are released in response to variation in muscular contraction following exercise of different intensity, mode, and volume. Some myokines may be anabolic and have direct growth-promoting effects, while others generate signals that may mediate some of the health benefits of PE.
In 2012, a new myokine expressed through the activation of transcription factor Peroxisome proliferator-activated receptor Gamma Co-activator-1α (PGC-1α) by exercise-induced effects, was discovered. The newly identified molecule has been called “irisin” and has been defined as a communicator between the SkM and adipocytes, and thus a potential bearer of positive effects of PE on other target tissues outside the muscle [7].
Irisin is cleaved from fibronectin type III domain containing 5 (FNDC5), a transmembrane precursor protein expressed in muscle under the control of PGC-1α. FNDC5 is also known to be profoundly expressed in many regions of thebrain, including cerebellar Purkinje cells, the hypothalamus, and the hippocampus, a region of the brain involved in memory and spatial awareness [7,8,9,10,11,12].
Previous work suggested that irisin may be responsible for the PE effect in some rodent models of neuropathological conditions, including cerebral ischemia and depression [13,14]. Furthermore, the overexpression of irisin and FNDC5 were associated with neuroplasticity, asthey may modulate neuronal proliferation [15], differentiation [16], and neurotrophin synthesis [8], conferring neuroprotection against amyloid peptide 1–42 (Aβ1–42) in mice [17], and having an antidepressant-like effect in rats subjected to unpredictable mild stress [13].
Similarly, to PE, the peripheral delivery of FNDC5 to the liver, via adenoviral vectors (resulting in elevated blood irisin levels), induced the expression of brain-derived neurotrophic factor (BDNF) and other neuroprotective genes in the hippocampus of mice [8]. In line with these results, the neurotrophin BDNF plays an important role in the homeostatic function and survival of the neurons; particularly in synaptic plasticity and neurogenesis. Decreased levels of BDNF have been identified in serum, as well as in hippocampal and cortex samples of AD and PD patients [18,19].
Since its discovery, irisin has been the subject of extensive research, which has enabled several insights to be gained about its pleiotropic properties. The role of irisin on memory and cognitive performance has been studied only recently, particularly during the last 5 years. Given the rising significance of irisin as a mediator of the beneficial effects of PE, in this review, we providean update onthe literature focusing on the relationship between FNDC5/irisin system and cognitive functions.
Irisin
Irisin Structure
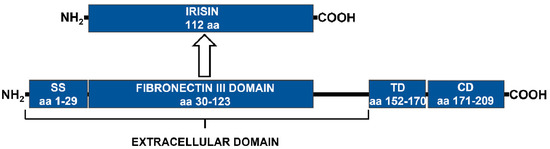
About 20 years ago, two independent groups identified FNDC5 as a protein exerting a role in the differentiation of myoblast. These first findings suggested that the gene was highly expressed in the SkM, but also in cerebral and cardiac tissues [54,55]. Human FNDC5 is a type I membrane protein of 212 amino acids (aa). The N-terminal is a signal sequence needed for final maturation and cleavage, the C-terminal is the cytoplasmatic domain, and in the middle there is a fibronectin III (FNIII) domain, an unknown domain, and a hydrophobic transmembrane domain [7,56,57].
The irisin represents the segment of FNDC5 that is cleaved under stimuli such as PE or cold. The portion contains 112 aa, formed by the residues from 29 to 140, including the tail at the C-terminus, the central FNIII domain, and the tail at the N-terminus [7,57]. The resulting peptide is characterized by a molecular weight of 12 kDa, and is dimerizing through the FNIII domain [56].
The irisin undergoes post-translational modification by N-glycosylation at two different residues. This modification, in addition to dimerization, determines a molecular weight reaching 35 kDa. The protein stability and the irisin secretion are regulated by the N-glycosylation. The process is strictly dependent on the presence of the signal peptide at the N-terminal, and is important for the irisin activity. The loss of glycosylation does not allow irisin to exert its main function in the browning stimulation of white adipose tissue (Figure 1) [58].
In contrast to rodents, the human FNDC5 gene has an ATA as a start codon instead of ATG, generating a transcript that results in a very low efficiency of translation [59]. More recently, Albrecht et al. (2020), identifying other non-canonical start codon, suggested that in SkM there are several transcripts for the human FNDC5 gene [60]. Studies on transcriptome profiling through RNA-sequencing (RNA-Seq) by the Functional Annotation of the Mammalian Genome/Genotype-Tissue Expression Project, established that the FNDC5 gene is mainly expressed in SkM, the heart, and several regions of the brain, mainly in the cerebellum, but also in hippocampus, cortex, and medulla oblongata [61].
A receptor for irisin has not yet been identified. The only evidence is provided by an interesting study by Kim et al. (2018), whichshowed that this myokine exerts its biological function by binding the integrins’ family of proteins. The integrins are ubiquitously expressed transmembrane receptors, consisting of eighteen α- and eight β-subunits, forming a total of 24 different heterodimers able to recognize also soluble ligands. Kim and coworkers described how the binding of irisin with the αVβ5 integrin heterodimer occurs in human adipocytes and osteocytes. Using the integrin inhibitor RGD peptide, which binds to αVβ5 in a selective manner, they also showed that any signaling response induced by irisin was significantly suppressed in these cells [62].
Irisin Functions
Bostrom and colleagues were the first to showthat irisin levels in the blood increase after PE, describing an increase of 65% in blood concentration in mice submitted to regular running for 21 days [7]. The level of irisin after PE is dependent on the type of physical activity, where training based on aerobic exercise is a higher inductor of serum irisin compared to resistance exercise [63,64].
In general, the level of irisin is influenced by lifestyle, characterized by specific residential place and associated activities, as suggested by the differences recorded between rural and urban inhabitants. Irisin concentration is lower in urban citizens, with a mean value of 3.6 ng/mL, while active individuals that live in rural areashad amean value of 4.3 ng/mL [65]. PE increases the level of irisin in the blood of healthy people [7], and in people with metabolic disorders [66]. Its circulating level is also related to the phenotype of different disease, as well obesity, type 2 diabetes [67], chronic renal disease [68] and hypothyroidism [69].
The first identified function of irisin was the “browning” of adipose tissue, in which irisin increases the expression of the mitochondrial protein uncoupling protein-1 (UCP-1) in mature fat cells, allowing the conversion of the white adipose tissue (WAT) to the brown adipose tissue (BAT) phenotype. The process ends with the formation of a third type of adipose tissue phenotype, named beige/brite adipose tissue. Irisin and PGC-1α regulate the expression of UCP-1 and thermogenesis in BAT, driving the metabolism of glucose and lipids toward the increase in energy consumption [70,71].
Furthermore, irisin is implicated in glucose homeostasis, by acting on different cell types and tissues involved in glucose metabolism, such as adipose tissue, SkM, liver, and pancreatic β cells. Due to this property, irisin is able to improve insulin sensitivity under insulin resistance (IR) conditions [72].
A decrease in irisin levels was associated with an increased risk of presenting metabolic syndrome and hyperglycemia in obese adults. This myokine shows negative associations with fasting insulin and glycosylated hemoglobin [73]. Previous studies also suggested its negative correlation with fasting glucose and HOMA-IR in school-age students of both genders [74], and positive associations with insulin concentration, fasting glucose, and HOMA-IR [66,75,76,77].
Irisin has other specific functions, including in the heart and liver, where it exerts antiapoptotic effects on cardiomyocytes and hepatocytes through the induction of autophagy [45], and protects cells from ischemia-reperfusion injury [78,79]. At bone level, it has a favorable effect and represents a key molecule in the crosstalk between this tissue and SkM. Specifically, irisin increases the mass and strength of the cortical bones, positively modifying their geometry by reducingthe secretion of osteoblast inhibitors, and driving the expression of bone-specific genes [80].
In immune system functioning, irisin mediates the positive effect of regular/moderate physical activity, contributing to a reduction in systemic inflammation, and consequently protecting from the development of diseases associated with chronic inflammation [81].
The functions of irisin on the brain are described in more detail in the section “Irisin: a new bridge between exercise and cognitive functions”. In brief, this myokine increases the proliferation of hippocampal neuronal cells [15], and reduces the neuronal damage mediated by pro-oxidant stimuli [14]. The FNDC5/irisin system is important for long-term potentiation and memory in mouse hippocampal region, being involved in establishing synaptic plasticity and memory [82], and may contribute to the antidepressant effect of PE together with serotonin, by the activation of the PGC-1α/BDNF pathway [83].
reference link : https://www.mdpi.com/2076-3417/11/15/7120/htm
More information: Mohammad R. Islam et al, Exercise hormone irisin is a critical regulator of cognitive function, Nature Metabolism (2021). DOI: 10.1038/s42255-021-00438-z



{kind=link}