











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



I primi studi hanno indicato che il trattamento con anticorpi monoclonali in individui immunocompromessi potrebbe comportare un’evoluzione virale all’interno dell’ospite che colpisce preferenzialmente gli epitopi riconosciuti da questi anticorpi, sebbene non sia chiaro se ciò significhi un rischio reale di virus resistenti agli anticorpi trasmissibili.
L’emergere di mutazioni SARS-CoV-2 diffuse su più linee virali in grado di aumentare la trasmissibilità o di produrre reinfezione nonostante la vaccinazione ha causato recenti preoccupazioni [1, 2].
Per ciascuno di questi motivi, una migliore sorveglianza genomica sarà fondamentale per monitorare la diffusione di queste nuove varianti e rilevare l’emergere di nuovi lignaggi con proprietà simili.
Da marzo 2020 abbiamo monitorato l’introduzione e la diffusione di SARS-CoV-2 nell’area di servizio del Gundersen Health System, un sistema sanitario integrato con sede a La Crosse, WI e che fornisce assistenza in 21 contee del Wisconsin sudoccidentale, Iowa nordorientale e sudorientale Minnesota.
Abbiamo eseguito il sequenziamento dell’intero genoma virale su oltre 1.900 casi positivi da questa regione nell’arco della pandemia. Tra i nostri obiettivi principali c’era il monitoraggio dell’introduzione di varianti virali nella nostra area di servizio che erano state riconosciute altrove dalle misure di sorveglianza genomica come varianti preoccupanti. In natura, il nostro programma ha consentito la diagnosi precoce e il monitoraggio di polimorfismi clinicamente rilevanti in linee in cui non erano stati precedentemente identificati altrove.
Ciò suggerisce che potrebbe verificarsi un’evoluzione convergente verso alcuni fenotipi favorevoli alla trasmissione. La proteina spike svolge un ruolo chiave nell’ingresso virale [8, 9] ed è anche il bersaglio sia di anticorpi sviluppati naturalmente [10] sia di anticorpi monoclonali sintetici utilizzati come trattamento o profilassi post-esposizione [11]. Gli studi hanno indicato che i virus recanti E484K sono associati a una ridotta risposta agli anticorpi neutralizzanti indotti dal vaccino [12].
Gli anticorpi monoclonali terapeutici hanno ricevuto l’Emergency Use Authorization (EUA) in ambito ambulatoriale a seguito di studi che hanno dimostrato una modesta riduzione del rischio di ospedalizzazione [13-15]. L’autorizzazione iniziale di questi agenti ha rappresentato un calo dell’utilità clinica se i sottoceppi virali prevalenti circolanti si sono spostati per includere varianti per le quali questi anticorpi avevano una ridotta affinità, con la continua disponibilità a basarsi sul monitoraggio passivo dei dati di sequenza che si accumulano nei depositi.
Le tendenze epidemiologiche successivamente portano al ritiro di Bamlanivimab [16] (e in seguito Bamlanivimab/Etesevimab [17]) dall’uso clinico diffuso, sebbene la combinazione sia stata successivamente ripristinata per un uso diffuso e ampliata per l’uso come profilassi post-esposizione come varianti prevalenti in circolazione cambiato ancora una volta [18].
Casirivimab/imdevimab è diventato disponibile sotto EUA il 20/11/20 [19] e l’uso per la profilassi post-esposizione è stato successivamente autorizzato [20].
Il potenziale per questi trattamenti di selezionare per l’emergere di mutazioni di resistenza anticorpale è stato notato in precedenza in pazienti immunocompromessi strettamente monitorati [21-23], ma non è stata segnalata la sorveglianza post-somministrazione nei contatti stretti o nella comunità più ampia.
Tra novembre 2020 e settembre 2021, il nostro programma ha somministrato terapie con anticorpi monoclonali di Bamlanivimab, Bamlanivimab/Etesevimab o Casirivimab/Imdevimab a 1.043 pazienti positivi al COVID-19.
In parallelo, il nostro programma di sequenziamento regionale SARS-CoV-2 [24-26] ha fornito l’opportunità di rilevare i lignaggi emergenti contenenti E484K e accertare la potenziale associazione epidemiologica, se presente, con individui che ricevono una terapia con anticorpi monoclonali.
collegamento di riferimento: https://www.medrxiv.org/content/10.1101/2021.10.02.21264415v1
Evoluzione di SARS-COV-2 e impatto sul riconoscimento degli anticorpi neutralizzanti
L’emergere di varianti mutanti di SARS-CoV-2 e il loro impatto sull’efficacia di nAbs e vaccini è diventata una delle principali preoccupazioni nell’attuale progresso della pandemia (23-25). Si presume che la pressione selettiva immunitaria durante le infezioni protratte abbia contribuito all’emergere di queste varianti (26).
Sebbene i livelli di immunità di gregge aumentino attraverso l’immunità naturale e indotta dal vaccino, è ragionevole ipotizzare che ciò fornirà una maggiore pressione selettiva sul virus e potremmo vedere che altri mutanti continuano ad emergere. A causa del loro potenziale di trasformare la forza e la cinetica del legame con ACE2, le mutazioni che si verificano nell’RBD sono di particolare interesse.
La prima variante ad emergere e rapidamente diventata dominante in tutto il mondo portava una sostituzione D614G e sebbene gli studi suggerissero una maggiore trasmissibilità (27-29), questa variante è stata neutralizzata con anticorpi monoclonali esistenti e sieri convalescenti (30-32). Particolarmente preoccupante è stata l’emergere della sostituzione N501Y, che è stata segnalata per essere più trasmissibile rispetto al tipo selvatico e alla sostituzione D614G (11, 33, 34).
La sostituzione N501Y è stata osservata per la prima volta nella variante B.1.1.7 (23) e successivamente nelle varianti B.1351 (24) e P1 (35). L’altro che riguarda la sostituzione della fuga immunitaria nel RBD è E484K, che è presente anche nelle varianti B.1.351 e P.1. Le varianti B.1351 e P.1 ospitano anche la sostituzione K417N e K417T, rispettivamente (23-24, 35).
È stato riportato che la presenza della sostituzione N501Y aumenta l’affinità con ACE2 di 7 volte e la combinazione additiva di sostituzioni a 417, 484 e 501 ha mostrato un’ulteriore maggiore affinità con ACE2 (19 volte rispetto a Wuhan) (11, 36 ). Le altre sostituzioni RBD significative sono Y453F nella variante B.1.1.298 (37), L452 nelle varianti B.1.429 (25) e B.1.617 (38).
Recentemente, l’OMS ha rinominato le varianti utilizzando lettere greche per riferirsi alle varianti come: B.1.1.7 come Alpha, B.1.351 come Beta, P1 come Gamma e B.1.617.2 come Delta (39, 40) (Tabella 1). Una variante è caratterizzata come variante preoccupante (VOC) se dimostra una maggiore trasmissibilità, un aumento della virulenza, un cambiamento nella presentazione della malattia o una ridotta efficacia di: vaccini, test diagnostici e misure di trattamento (40).
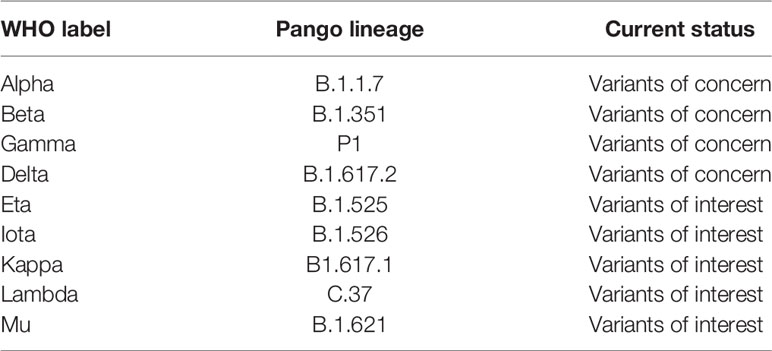
Questi COV circolanti di SARS-CoV-2, inclusi B.1.1.7 (23), B.1.351 e P.1 (35, 41), mostrano una ridotta suscettibilità ad alcuni mAbs SARS-CoV-2 (11, 42-44) , plasma convalescente (11, 45) e sieri di vaccinati SARS-CoV-2 (11, 36, 44, 46, 47). La variante Delta, B.1.617.2, è diventata la variante SARS-COV-2 dominante in tutto il mondo (48) ed è associata ad un aumento della replicazione virale che porta ad una maggiore trasmissibilità, carica virale e gravità più elevate (49, 50). L’emergere di B.1.617.2 è associato all’evasione degli mAbs e all’efficacia del vaccino (50, 51).
La disponibilità di nAbs terapeutici efficaci contro tutte le varianti di SARS-CoV-2 offrirà vantaggi per il controllo delle varianti pandemiche attuali e future, e il loro sviluppo rimane pertanto una priorità elevata (52). La presenza di ampi mAb è stata descritta in pazienti convalescenti e offre speranza che gli attuali vaccini possano essere efficaci (53). La maggior parte dei mAb isolati da pazienti convalescenti sono anticorpi di Classe 1 (18, 19).
La maggior parte degli anticorpi ampiamente neutralizzanti (bnAbs) descritti fino ad oggi rientrano anche in questa stessa classe (9-11), tuttavia, sono stati caratterizzati bnAbs appartenenti alla Classe 4 con elevata potenza e ampiezza neutralizzante (21, 22). La classe 2 comprende alcuni potenti Ab (C144, C121, COVA2-15, COVA2-37), tuttavia la loro efficacia contro le varianti emergenti non è stata descritta (19, 54). Gli anticorpi di classe 3 si legano a un epitopo conservato e generalmente non sono influenzati dalle mutazioni (19, 55).
Comprendere le diverse classi di mAb ampi che vengono generati e la loro persistenza nei pazienti convalescenti guariti e nei vaccini è importante per comprendere la robustezza dell’immunità di gregge che viene generata. Qui esaminiamo alcuni degli anticorpi ampiamente neutralizzanti più promettenti e la loro modalità di interazione ottenuta da pazienti convalescenti. Sarà importante capire quanti di questi ampi anticorpi neutralizzanti vengono conservati dopo la vaccinazione e l’infezione naturale.
Classificazione degli anticorpi in base alla loro modalità di legame con l’RBD della proteina Spike
In questa recensione utilizzeremo principalmente la classificazione di Barnes, sebbene le classificazioni di Piccoli et al. saranno occasionalmente incrociati (tabella 2).
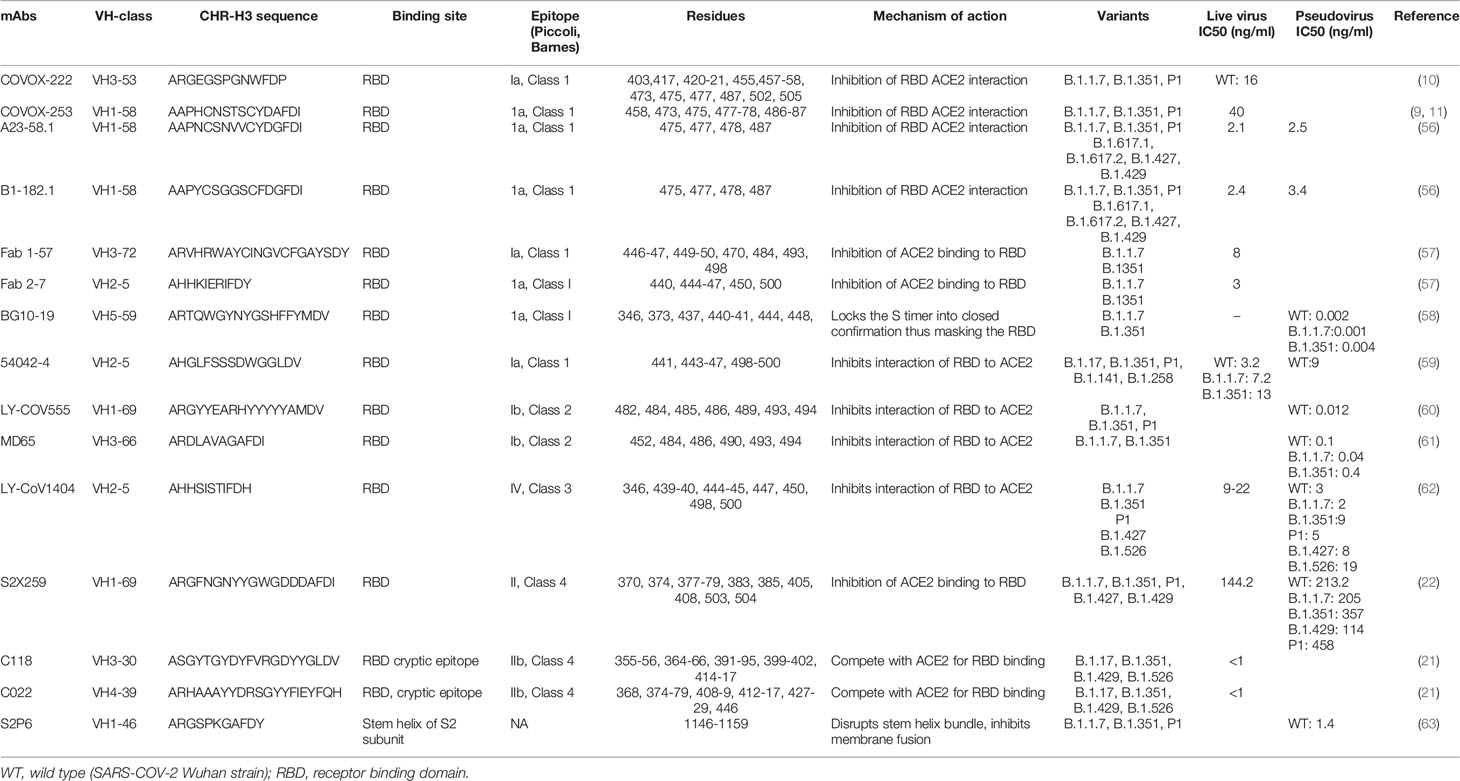
Anticorpi di classe 1
Gli anticorpi di classe 1 sono i più immunodominanti tra gli anticorpi diretti contro l’RBD e sono solitamente codificati dalle linee germinali VH3-53 e VH3-66 (5, 10, 17, 54). È stato descritto che la famiglia degli anticorpi IGHV3-53 condivide proprietà di legame comuni e si lega a un epitopo comune attorno al collo dell’RBD, con un approccio identico condiviso anche dagli Abs derivati da IGHV3-66 (5, 11, 36). Il legame e la neutralizzazione di questa classe di anticorpi sono generalmente annullati dalle mutazioni N501Y, E484K e K417N (64).
Il loro impegno con il RBD è dettato da CDR-H1 e CDR-H2, mentre il CDR-H3 è breve e fa poche interazioni (5, 10, 65). Tuttavia, CDR-H3 interagisce con K417 e CDR-L1 con N501 e, pertanto, l’attività di neutralizzazione di molti anticorpi di classe I VH3-53 è compromessa dalla sostituzione N501Y nei virus varianti B.1.1.7, B.1.351, e P.1, mentre la sostituzione aggiunta a 417 in P.1 e B.1.351 ha un effetto negativo additivo sull’attività neutralizzante (10).
Quindi, la potenza neutralizzante della maggior parte degli mAbs di Classe I è notevolmente ridotta rispetto alle varianti (66). La classe 1, però, contiene anche diversi nAb in grado di neutralizzare le varianti emergenti. In questa sezione discutiamo molti di questi anticorpi e descriviamo la loro interazione con il SARS-CoV-2 Spike.
Dejnirattisai et al. isolato il mAb di classe I, COVOX-222, che è derivato da VH3-53, che neutralizza tutte e tre le varianti nonostante il legame con due delle sostituzioni del sito di legame ACE2 (10, 36) (Tabella 2). Nel virus originale, il residuo 417 crea una debole interazione sale-ponte con il residuo CDR3 a catena pesante E99 che viene abolito a causa di una sostituzione con asparagina o treonina (10).
Il CDR-H3 di COVOX-222 (13 residui) è leggermente più lungo della maggior parte degli anticorpi VH3-53; tuttavia, sembra improbabile che questa sia l’unica ragione per la resilienza di COVOX-222 (10, 65). C’è poca energia di legame dal CDR3-H3, poiché la maggior parte dell’energia di legame della catena pesante proviene da CDR-H1 e CDR-H2, che interagisce debolmente con il residuo RBD 417. Pertanto, molti degli anticorpi VH3-53 sono probabili essere volatile alle mutazioni al residuo 417 (K417N in B.1.351 e K417T in P1).
Il COVOX-22 CDR L1 interagisce con il residuo 501 del RBD attraverso il suo residuo P30. L’interazione è ulteriormente rafforzata dalla sostituzione N501Y, che alla fine si aggiunge alla resilienza di questo anticorpo (10). Quindi, COVOX-222, un mAb di classe 1, famiglia di geni VH3-53, nonostante il suo breve CDR-H3 e il legame con i residui RBD mutanti,
È stato anche riportato che un altro gruppo di mAbs di classe I, indicato come 55, 165, 253 e 318, mantiene l’attività di nAb contro le varianti (67). Questi quattro mAb sono tutti di classe IGHV1-58, hanno meno mutazioni non silenti (2-5) e una CDR3 a catena pesante più lunga (12-16 residui). mAb 55, 165 e 253 si legano all’epitopo dominante del collo dell’RBD mentre 318 si lega a un epitopo sulla spalla destra.
Anche i titoli FRNT50 per mAbs 55, 165, 253 e 318 sono relativamente uguali tra il ceppo Victoria (A.1) (67) e B.1.351, indicando che i loro epitopi non sono influenzati dalle mutazioni K417N, E484K e N501Y (9 , 11). Questi mAbs, nonostante il diverso utilizzo del gene VH rispetto al COVOX-222, sono ancora in grado di neutralizzare il SARS-COV-2 di tipo selvaggio e le varianti esistenti.
Un gruppo separato di ricercatori ha isolato due anticorpi, A23-51.1 e B1-182.1, da soggetti convalescenti infettati dal ceppo Washington-1 (WA-1), che ha una sequenza S identica a Hu-1 (56). Questi anticorpi hanno mostrato la capacità di mantenere un’elevata potenza di neutralizzazione contro 10 varianti di proteine spike tra cui B.1.1.7, B.1.351, P1 e la variante altamente trasmissibile B.1.617.2.
I due anticorpi, A23-58,1 e B1-182.1, condividono un uso simile della famiglia di geni nelle loro catene pesanti e leggere; entrambi utilizzano catene pesanti IGHV1-58 e catene leggere IGKV3-20/IGKJ1 e bassi livelli di SHM. Entrambi questi mAb hanno una modalità simile di legame con RBD. Mentre si lega, il residuo RBD 486 si immerge nel cratere formato dai CDR e forma un motivo uncinato che è stabilizzato da un legame disolfuro intra-ansa tra i residui 480 e 488 mentre i residui aromatici, inclusi 456, 473, 486 e 489 forniscono 48% (299 Å2) dell’epitopo.
Rispetto agli epitopi di altri anticorpi, il supersito definito dai contatti comuni di questi anticorpi derivati da IGHV1-58 ha avuto meno interazioni con i residui negli hotspot mutazionali. Il motivo a uncino e il cratere CDR sono essenziali per la rilegatura all’interno della classe pubblica VH1-58 (56). Questa combinazione di famiglie di geni anticorpali è stata proposta come un clonotipo pubblico poiché è stata identificata in altri soggetti convalescenti COVID-19 (6, 9, 54).
Un altro studio ha recentemente caratterizzato sei anticorpi potentemente neutralizzanti da cellule B di soggetti convalescenti (58). Tra questi, BG10-19 ha neutralizzato potentemente i virus SARS-CoV-2 wild-type, B.1.1.7 (23), B.1.351 e i virus pseudotipizzati SARS-CoV eterologhi (58). BG10-19 utilizza cinque dei sei anelli CDR per connettersi con un epitopo proteoglicano diretto sulle eliche RBD α-1 (338-347) e α-2 (364-374), con contatti aggiuntivi con 436-450.
I loop CDRH2 e CDRH3 moderano la maggior parte dei contatti RBD (~760Å2), stabilendo forti interazioni con i residui RBD. L’interazione dei loop CDRH1-3 e CDRL2 stabilisce l’epitopo primario riconosciuto da BG10-19, che non si sovrappone al motivo di legame del recettore ACE2. Nel complesso, BG10-19 impiega un meccanismo di neutralizzazione che maschera l’impegno dell’ACE2 da parte dell’RBM, bloccando il trimero Spike in una conformazione chiusa (58).
Un altro mAb di Classe I di interesse sono gli anticorpi monoclonali Fab 1-57 e Fab 2-7 che si legano all’epitopo RBD al di fuori dell’hotspot della pressione evolutiva (57). Il riconoscimento di RBD wild-type da parte di Fab 1-57 è dominato dalla catena pesante, che seppellisce 533,7 di superficie, con un contributo minore di 223,3 da parte della catena leggera. Rispetto alle mutazioni nelle varianti B.1.1.7, B.1.351 e P1, solo il residuo 484 era vicino al sito di legame di Fab1-57.
Nonostante la sua vicinanza all’epitopo, tuttavia, 484 non interagiscono in modo significativo con Fab 1-57. La modellizzazione strutturale della mutazione E484K ha mostrato che il residuo K484 era geometricamente compatibile con il legame di Fab 1-57 alla serina 29 con un legame idrogeno (57). L’interazione di RBD wild-type da parte dell’anticorpo Fab 2-7 è dominata dalla connessione prossimale alle anse RBD formate dai residui 438-451 e 495-502. I residui di CDR H2 54, 52 e 58 hanno formato legami idrogeno con i residui di RBD 450, l’ammina principale di 445 e 447, rispettivamente.
Il residuo 32 della catena leggera ha anche interagito con il residuo 440 per formare un legame idrogeno. Per quanto riguarda i tre residui mutati, l’anticorpo Fab 2-7 si è legato solo vicino a N501, ma la catena laterale di N501 puntava lontano dall’anticorpo.
Anche se si verificherebbe qualche cambiamento conformazionale dell’ansa 495-502 a causa della mutazione N501Y, questo anello contribuisce solo con 225 2 su 736 Å2 e conteneva pochi residui che formano interazioni significative con il Fab (57). L’uso terapeutico di Abs, come Fab 1-57 e 2-7, che colpiscono epitopi meno diffusi, potrebbe mitigare la preoccupazione della fuga di mAb.
Oltre ai metodi esistenti, a questo scopo vengono impiegate anche nuove tecnologie (68). Kramer et al. (59) hanno utilizzato la tecnologia LIBRA-seq (69) e hanno identificato un potente anticorpo monoclonale 54042-4 di un paziente convalescente COVID-19 che ha legato e neutralizzato i virus SARS-CoV-2 vivi, comprese le varianti preoccupanti.
Hanno concluso che l’anticorpo 54042-4 si legava a queste varianti RBD (B.1.17, B.1.351, P1, B.1.141, B.1.258) a un livello paragonabile al legame con l’RBD dell’isolato Wuhan-1. La struttura crio-EM ha mostrato che 54042-4 forma una vasta interfaccia con l’RBD attraverso tutti e tre i CDR della catena pesante, CDRL1 e CDRL3 per formare un morsetto sull’apice della sella RBM.
La catena pesante interagisce con i residui RBD 443-447, mentre la catena leggera interagisce con i residui 445 e 498-500. CDR H1 si lega a 441, CDR H2 a 444 e CDR H3 a 443. CDR L1 e L3 formano un legame idrogeno con 445 e 498-500. La complessa interazione ha indicato che è improbabile che le sostituzioni degli spike negli attuali VOC influenzino l’affinità di legame di 54042-4.
Il residuo RBD N501 si trova al di fuori dell’epitopo 54042-4, mentre gli atomi di Cα di E484 e K452 sono rispettivamente di 18 e 14 Å dal residuo 54042-4 più vicino. L’uso della tecnologia moderna come LIBRA-seq consente la determinazione simultanea ad alto rendimento della sequenza del recettore delle cellule B e della reattività dell’antigene a livello di singola cellula, facilitando il processo di selezione e caratterizzazione del candidato e accelera anche lo sviluppo di anticorpi ampiamente neutralizzanti.
La classe 1 comprende una vasta gamma di mAb con una forte ampiezza e potenza di neutralizzazione. Nonostante la variazione nel loro utilizzo del gene VH, tutti i suddetti mAbs possono neutralizzare efficacemente le varianti emergenti di SARS-COV-2. Questi anticorpi potrebbero essere utilizzati da soli o in combinazione con altre classi di anticorpi nella produzione di anticorpi terapeutici e guidare anche la formulazione di vaccini di prossima generazione.
Anticorpi di classe 2
La classe 2 comprende alcuni potenti Abs (C144, C121, COVA2-15, COVA2-37) che possono legarsi all’RBD sia nella conferma up che down. In generale, la loro efficacia contro le varianti emergenti non è stata descritta (19, 54). Tuttavia, sono stati ben descritti due mAb di Classe 2 e l’impatto di mutazioni note. LY-CoV555 (nome clinico bamlanivimab), attualmente utilizzato in ambito clinico in combinazione con etesivimab, è un potente anticorpo neutralizzante anti-spike (70, 71).
L’analisi strutturale ha rivelato che si lega sia nella conferma verso l’alto che verso il basso dell’RBD e il legame di questo mAb all’RBD non è influenzato dalla mutazione N501Y; tuttavia, la mutazione E484K abolisce il legame (60). Contro B.1.351 sono abolite anche le attività del bamlanivimab e di un altro mAb terapeutico REGN10933 appartenente agli anticorpi di classe 1 (66).
Un altro anticorpo di classe 2, MD65, ha un pattern di legame simile a LY-CoV555 ma, a differenza di LY-CoV555, la mutazione E484K non ha influenzato la sua efficacia di legame (61).
La valutazione in vivo su topi transgenici K18-hACE2 ha suggerito che MD65 ha mostrato efficacia contro la variante B.1.1.7, B.1.351 e P1 (61). Quindi, sembra che gli mAbs di Classe 2 ampiamente reattivi possano essere sviluppati nonostante le carenze di bamlanivimab rispetto a B.1.351.
Anticorpi di classe 3
Gli anticorpi che prendono di mira l’epitopo di Classe 3 possono legarsi con l’RBD in entrambi gli stati “su” e “giù” (18, 19). L’epitopo di classe 3 è altamente conservato nei cladi 1, 2 e 3 di Sarbecovirus, il che indica che è un buon bersaglio per ampi anticorpi neutralizzanti e suggerisce che è conservato funzionale e ha meno probabilità di essere associato al sistema immunitario (55, 72). Gli anticorpi di classe 3 si legano al di fuori della regione di legame ACE-2 e, quindi, forniscono il potenziale per effetti sinergici quando combinati con nAbs che intercettano il legame ACE2.
L’anticorpo monoclonale di classe 3 LY-CoV1404 è stato isolato da uno schermo ad alto rendimento di cellule mononucleate del sangue periferico ottenuto da un soggetto convalescente 60 giorni dopo l’insorgenza dei sintomi (62).
Utilizzando saggi di neutralizzazione autentici e pseudovirus, i ricercatori hanno dimostrato che LY-CoV1404 mantiene una potente attività neutralizzante contro più varianti tra cui B.1.1.7, B.1.351, B.1.427, P.1 e B.1.526.
È interessante notare che LY-CoV1404 condivide il 92% dell’identità della sequenza di amminoacidi nelle regioni variabili delle sue catene pesanti e leggere, per l’anticorpo Fab 2-7 (57), sebbene siano stati scoperti indipendentemente da diversi pazienti suggerendo che forse un repertorio condiviso pubblicamente ( 62).
Entrambi gli anticorpi si attaccano anche al RBD in modo simile. LY-CoV1404 si lega a una regione che si sovrappone al sito di interazione ACE2 del picco che è accessibile nello stato aperto e chiuso del RBD. Sebbene questa proprietà suggerisca che si tratta di un anticorpo di classe 2 (5, 18), la posizione dell’epitopo è identica a mAb S309, un legante di classe 3 (55). Sebbene l’epitopo legante LY-CoV1404 includa residui N501 e N439, possono legare le varianti B.1.1.7 e B.1.351 e sono neutralizzati con la stessa forza del virus di tipo selvatico (62).
I risultati di cui sopra suggeriscono che LY-CoV1404 rappresenta un potente mAb con un ampio intervallo di neutralizzazione, possiede la proprietà degli anticorpi di classe 2 e 3, ha un epitopo relativamente conservato e potrebbe essere impiegato per affrontare il problema delle varianti emergenti.
La potente neutralizzazione di SARS-CoV-2 da parte di LY-CoV1404 consente l’esplorazione di dosi cliniche più basse, che possono supportare la somministrazione sottocutanea e ha il potenziale per fornire un complemento a lungo termine ai vaccini nel probabile caso in cui COVID-19 diventi endemico. I nAb di classe 3 integrano il repertorio di anticorpi anti-SARS-CoV-2 e possono essere efficacemente utilizzati in combinazioni terapeutiche con nAb di classe 1 o 2 (18).
Un altro gruppo di anticorpi della classe 3 sono VIR-7831 e VIR-7832; mAb a doppia azione derivati dall’anticorpo genitore S309, un anticorpo ottenuto da un sopravvissuto alla SARS-CoV (55). Questi mAb sono stati progettati per avere un’emivita estesa e una migliore biodisponibilità polmonare (73) e mirare a un epitopo situato intorno al residuo N343 che è altamente conservato tra i Sarbecovirus e neutralizzare virus vivi e pseudotipizzati contro B.1.1.7, B varianti .1351 e P1. Esibiscono anche una potente funzione effettrice e conferiscono citotossicità cellulare anticorpo dipendente e fagocitosi cellulare anticorpo dipendente in vitro (74). Il mAb VIR-7831 ha ricevuto autorizzazioni all’uso di emergenza (EUA) dalla FDA statunitense per il trattamento di COVID-19 da lieve a moderato (75).
Anticorpi di classe 4
Gli Abs di classe 4 si legano all’epitopo criptico altamente conservato sull’RBD al di fuori dell’RBM (18). La maggior parte degli mAb descritti in precedenza sono cross-reattivi ma debolmente neutralizzanti (1, 17, 76-78). Tuttavia, Jette et al. (21) hanno caratterizzato due anticorpi anti-RBD di classe 4, C118 e C022, che sono stati ottenuti da donatori di COVID-19 e hanno rivelato un ampio riconoscimento e una potente neutralizzazione delle varianti di SARS-CoV-2 (21).
Hanno scoperto che C118 e C022 Abs hanno neutralizzato quattro varianti di SARS-CoV-2 (B.1.17, B.1.351, B.1.429, B.1.526). L’analisi della struttura ha mostrato che entrambi i mAbs hanno riconosciuto un epitopo che è altamente conservato alla base del RBD, che è divulgato solo nelle conformazioni “up”. C118 e C022 utilizzano quattro dei sei anelli CDR per legarsi all’epitopo che si estende verso la cresta RBD vicino alla regione di legame ACE2, e C022 include un residuo di interazione sovrapposto aggiuntivo a 417.
I loop CDRH3 e CDRL2 e porzioni della regione quadro L3 di entrambi gli anticorpi dominano i contatti RBD e sviluppano interazioni polari e di van der Waals che rappresentano il 71% della superficie sepolta dell’epitopo (21). C022 e C118 formano notevoli interazioni di backbone con RBD, con 9 e 10 legami H formati con RBD, che contribuiscono rispettivamente alla loro neutralizzazione incrociata tra varianti e ampiezza, poiché queste interazioni mediano il legame nonostante le sostituzioni della catena laterale.
L’isolamento e la caratterizzazione di nAbs mirati agli epitopi di RBD conservati che possiedono un duplice vantaggio di ampiezza tra i Sarbecovirus e una maggiore resistenza alla fuga di neutralizzazione sono necessari per combattere questa pandemia (79). A causa della conservazione dell’epitopo di classe IV, gli anticorpi diretti a questo sito sono di interesse. S2X259, è un altro anticorpo di classe 3 isolato (22) da un paziente convalescente che neutralizza ampiamente l’ingresso di SARS-CoV-2 comprese le varianti B.1.1.7, B.1.351, P.1 e B.1.427/B.1.429. Questo anticorpo agisce attraverso l’inibizione del legame di ACE2 al RBD.
Hanno anche eseguito diversi esperimenti per dimostrare che questo anticorpo è efficace contro un ampio spettro di sarbecovirus umani e zoonotici e mantiene un’elevata barriera alla comparsa di mutanti resistenti (22). S2X259 prende di mira un epitopo criptico privo di glicani all’interno del sito antigenico IIa e si lega all’RBD utilizzando catene pesanti e leggere che contribuiscono rispettivamente a due terzi e un terzo della superficie del paratopo sepolta al momento del legame (1, 22).
S2X259 utilizza CDR H1-H3, L1 e L3 per interagire con i residui 369-386 formando due α-eliche e un -foglio intermedio. L’epitopo S2X259 è altamente conservato nei virus SARS-CoV-2 e non contiene residui mutanti prevalenti, come L452R, S477N o N439K. Il mAb si lega alla spina dorsale del residuo N501 e non alla sua catena laterale, aggira i residui 417 e 484 il che potrebbe spiegare la potenza neutralizzante conservata contro B.1.1.7, P1, B.1.351 e B.1.429 (22).
A causa del suo epitopo conservato e criptico, gli anticorpi di Classe 4 sono risorse migliori per la neutralizzazione attraverso più varianti e quindi potenzialmente proteggono dai Sarbecovirus emergenti (21, 22).
link di riferimento: https://www.frontiersin.org/articles/10.3389/fimmu.2021.752003/full?fbclid=IwAR23Dbry666btcHAdEsk0kMrI-XejO6XXsX44DkNEa2A-DF5Nkqh91Y88aE



{kind=link}