









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")


The study team identified rare, naturally occurring T cells that are capable of targeting a protein found in SARS-CoV-2 and a range of other coronaviruses.
The study findings suggest that a component of this protein, called viral polymerase (RdRp/NSP12), could potentially be added to COVID-19 vaccines to create a longer-lasting immune response and increase protection against new variants of the virus.
Cross-reactivity and direct killing of target cells remain underexplored for SARS-CoV-2 specific CD8+ T cells. Isolation of T cell receptors (TCRs) and overexpression in allogeneic cells allows for extensive T cell reactivity profiling.
The study team identified a SARS-CoV-2 RNA-dependent RNA-polymerase (RdRp/NSP12) as highly conserved likely due to its critical role in the virus life cycle.
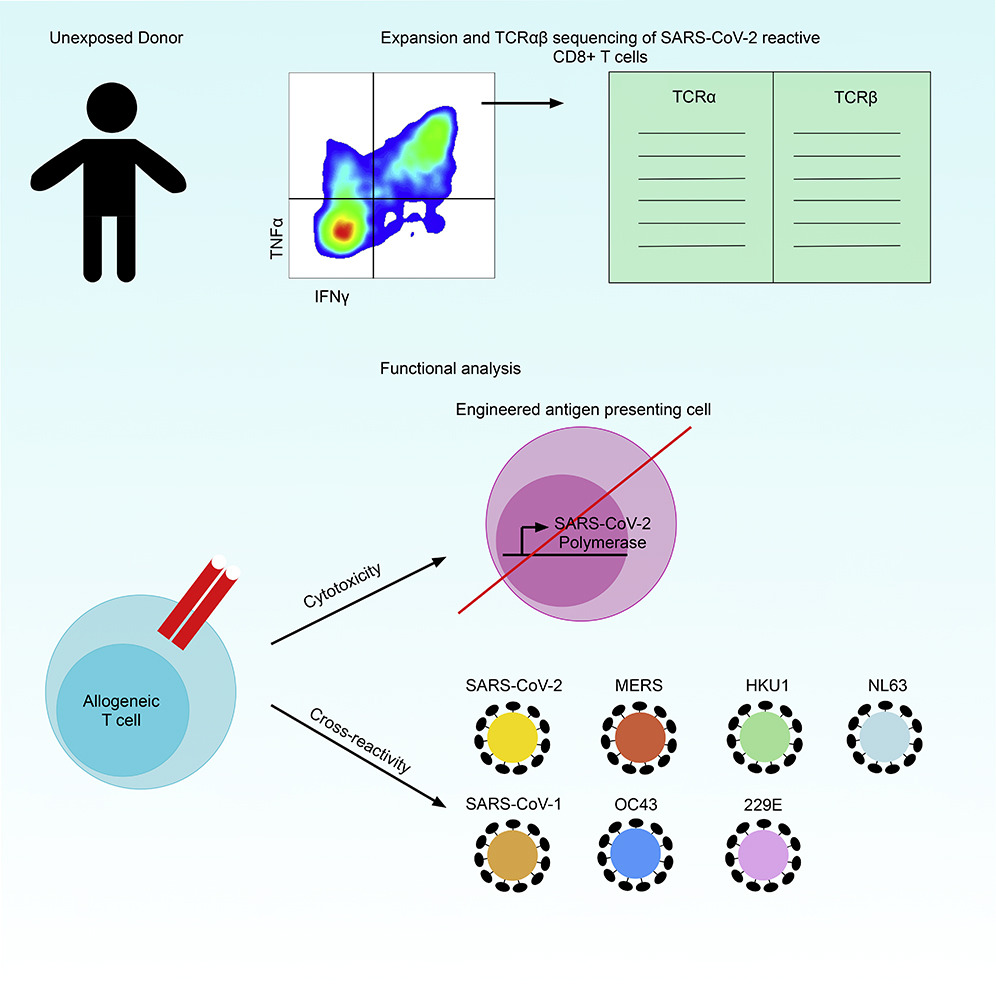
The team performed single-cell TCRαβ sequencing in HLA-A∗02:01 restricted, RdRp specific T cells from SARS-CoV-2 unexposed individuals.
The study findings showed that human T cells expressing these TCRαβ constructs kill target cell lines engineered to express full length RdRp.
Graphical Abstract
Importantly three TCR constructs recognize homologous epitopes from common cold coronaviruses, indicating CD8+ T cells can recognize evolutionarily diverse coronaviruses.
Detailed analysis of individual TCR clones may help define vaccine epitopes that can induce long term immunity against SARS-CoV-2 and other coronaviruses.
The study findings were published in the peer reviewed journal: Cell Reports (By Science Direct). https://www.sciencedirect.com/science/article/pii/S2211124721016673
Over 4 million people have died from COVID-19 as of August 2021 (World Health Organization). Many individuals are now immune as a result of successful vaccination campaigns and protection afforded by the natural infection with SARS-CoV-2 (Anand et al., 2021; Baden et al., 2021; Lumley et al., 2020; Polack et al., 2020; Sadoff et al., 2021).
The virus continues to evolve and may escape immune responses generated against the original sequence (Harvey et al., 2021; Planas et al., 2021). The BNT162b2 mRNA vaccine is 88% effective against the new Delta variant compared with 93.7% for the Alpha variant that was circulating previously (Bernal et al., 2021). Increased spread in vaccinated populations necessitates further understanding of the SARS-CoV-2 immune response.
This pandemic can only be controlled by herd immunity against contemporary strains of the virus. Vaccination against the wild type spike protein can prevent COVID-19 (Baden et al., 2021; Polack et al., 2020; Sadoff et al., 2021). SARS-CoV-2 vaccines target the spike protein by generating neutralizing antibodies that prevent host cell infection (Khoury et al., 2021; Lumley et al., 2020).
SARS-CoV-2 variants often contain multiple mutations in the spike protein and can resist antibody neutralization creating the possibility that, upon further diversification, viral variants may escape current vaccine defenses (Hoffmann et al., 2021; Kuzmina et al., 2021; Muik et al., 2021; Planas et al., 2021; Wang et al., 2021).
Cytotoxic T cells kill infected cells thereby directly limiting viral dissemination once the infection occurred (Hall et al., 1986; Harty et al., 2000; Jozwik et al., 2015; McMichael et al., 1983). T cell recognition is not limited to surface proteins like the spike protein; more conserved proteins can be targeted. Internal SARS- CoV-2 proteins are more conserved than the spike and may present a therapeutic opportunity at
generating T cell responses that can recognize many coronavirus strains (Grifoni et al., 2020a). T cell vaccine strategies, targeting the nucleocapsid protein, are being explored to generate long term immunity against SARS-CoV-2 (Dutta et al., 2020; Gauttier et al., 2020; Sieling et al., 2021). It remains unknown which epitopes elicit the most effective antiviral responses (Chen and John Wherry, 2020).
Initial evidence for T cell control of respiratory infections was provided by children with genetic defects in T cell development(Hall et al., 1986). Resident memory T cells, which are permanently localized in non-lymphoid tissues, including the lung, are thought to mediate anti- viral responses(Jozwik et al., 2015).
In a human RSV infection disease severity was inversely correlated with the preexisting T cells in the lung(Jozwik et al., 2015). Adoptive transfer of highly functional T cell clones can reduce severity of viral diseases as well(Einsele et al., 2002; Feuchtinger et al., 2010a). The mechanism of respiratory viral infection T cell control is thought to happen through FAS and perforin mediated lysis of infected cells(Topham et al., 1997). The efficiency of lysis correlates with the ability to clear an infection(McMichael et al., 1983).
Both convalescent donors and unexposed individuals have SARS-CoV-2 specific T cell responses (Le Bert et al., 2020; Braun et al., 2020; Grifoni et al., 2020b; Mateus et al., 2020; Peng et al., 2020; Tarke et al., 2021; Weiskopf et al., 2020). CD8+ T cell responses have been identified as correlates of protection in SARS-CoV-2 infection (Chen and John Wherry, 2020; Liao et al., 2020; McMahan et al., 2021). Unexposed individuals may have T cell responses that were generated by common cold coronaviruses (HCoVs) and may be partially protective against SARS-CoV-2 encounter(Lipsitch et al., 2020; Mallajosyula et al., 2021; Mateus et al., 2020). T
cells interact with target antigens through the T cell receptor (TCR), which is a heterodimer of alpha and beta chains. TCRs are inherently cross-reactive to maximize the breadth of ligand recognition, however a single TCR is not guaranteed to recognize related antigens (Sewell, 2012). Several cross-reactive CD8+ T cell responses are known, but specific TCRαβ clones that can drive such reactivity are not defined (Lineburg et al., 2021; Lipsitch et al., 2020; Mallajosyula et al., 2021; Mateus et al., 2020). T cell memory is most often defined as ability to recognize synthetic peptide epitopes in functional assays or peptide-MHC multimer staining.
Recognition of processed epitopes derived from full length intracellular antigens is underexplored in SARS-CoV-2. Isolation of specific TCR clones permits unambiguous determination of reactivity and detailed characterization of immune responses such as cytotoxic potential and measurement of cross-reactivity against related viruses.
We employ recent technological advances in single-cell sequencing, DNA synthesis and gene transfer to recover antigen specific TCRαβ and subsequently characterize them in allogeneic T cells. The viral polymerase (NSP12/RdRp) was identified as highly conserved within SARS- CoV-2 and other human coronaviruses. RdRp reactive CD8+ T cells were then selected for
TCRαβ droplet-based sequencing based on the intracellular level of TNFα and IFNγ via the CLInt-seq, which allows for antigen specific TCR sequencing via commercially available Drop- Seq in cells that are stained for intracellular cytokines (Nesterenko et al., 2021). TCRs were initially screened for single epitope recognition in a cell line system via the NFAT-GFP reporter system. Reactive TCRs were overexpressed in human PBMCs and killed antigen presenting cells that expressed the full length RdRp. Three TCR constructs were broadly reactive and cross- reacted with epitope homologues from HCoVs.



{kind=link}