










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A diet rich in fiber may help some people being treated for melanoma respond to immunotherapy treatment by influencing the gut microbiome, according to a new study led by researchers at the Center for Cancer Research at the National Cancer Institute (NCI), part of the National Institutes of Health, and the University of Texas MD Anderson Cancer Center. Results from the study, which analyzed both people with melanoma and mouse models of the disease, appeared December 24, 2021, in Science.
In contrast, use of probiotic supplements appeared to lessen somewhat the effectiveness of immune checkpoint blocker regimens. Probiotics are live microorganisms typically consumed as a supplement to improve gut health.
“The data suggest that one can target the composition of the gut microbiota and affect the ability of the patient to respond to immunotherapy,” said Giorgio Trinchieri, M.D., chief of the Laboratory of Integrative Cancer Immunology in NCI’s Center for Cancer Research, one of the study’s coleaders. “Consuming a diet rich in fiber, like fruits, vegetables, and legumes, could improve your ability to respond to immunotherapy.”
Immunotherapy with immune checkpoint blockers helps restore the immune system’s natural ability to recognize and kill tumor cells. These drugs have been transformative in melanoma, improving how long some people with advanced disease live, sometimes by years.
“The question is, can we change the composition of the type of bacteria in the gut and improve the ability of the patient to respond?” Dr. Trinchieri said.
In a previous study, Dr. Trinchieri and a different group of collaborators showed that some people with melanoma who initially did not respond to treatment with an immune checkpoint blocker did respond after receiving a fecal transplant from a patient who had responded to the drug.
The fecal transplant, they concluded, had introduced different gut bacteria that helped make it easier for immune cells to invade and kill their tumors.
“Dietary fiber intake and use of probiotic supplements have also been shown to affect the composition of gut bacteria. More cancer patients are taking probiotic supplements in an effort to improve their gut health, but little is known about how probiotics – which basically change the ecology of the gut bacteria – impact immunotherapy response,” he said.
The connection between fiber intake and immunotherapy response has also been unclear. However, a recent study led by Romina Goldszmid, Ph.D., also of NCI’s Center for Cancer Research, showed that mice fed a diet rich in pectin, which is a fiber abundant in apples, were able to stave off tumor growth by activating immune cells and reprogramming the tumor microenvironment.
In the new study, Dr. Trinchieri and study co-leads Carrie R. Daniel, Ph.D., M.P.H., and Jennifer A. Wargo, M.D., of the University of Texas MD Anderson Cancer Center, and their collaborators looked at the composition of fecal microorganisms (the gut microbiota), dietary habits, and probiotic supplement use among patients who were being treated for advanced melanoma with immune checkpoint blockers.
Among the 128 patients whose dietary fiber intake was known, those who reported consuming at least 20 grams of dietary fiber per day (an amount the researchers designated as “sufficient” for the purposes of this study) lived longer without their cancer progressing than those who consumed less dietary fiber. Every 5-gram increase in daily dietary fiber intake corresponded to a 30% lower risk of progression of the disease.
To mimic the different diets in the melanoma patients, they fed mice either a fiber-rich or a low-fiber diet, injected the mice with melanoma cells, and then treated the mice with anti-PD-1 therapy.
Mice given the fiber-rich diet had delayed tumor growth after anti-PD-1 treatment, compared with mice given the low-fiber diet.
The researchers then repeated the experiments in germ-free mice—that is, mice that have no bacteria in their guts.
“In germ-free mice, the diet made no difference in the immunotherapy response,” Dr. Trinchieri said. “That suggests that the diet is affecting the response to immune checkpoint therapy by changing the composition of the gut microbiota.”
Dr. Trinchieri noted that one possible mechanism through which dietary fiber exerts its beneficial effect is by increasing the types of bacteria in the gut, such as Ruminococcaceae, that produce high levels of certain short-chain fatty acids that have an antitumor effect.
“We did see an increase in one of these short-chain fatty acids, propionate, in mice that were fed a high-fiber diet,” Dr. Trinchieri said. “Moreover, patients whose cancer responded to immunotherapy had a greater abundance of Ruminococcaceae bacteria in their gut microbiota compared with those who did not respond to therapy.”
The researchers also looked at the impact of probiotics on gut bacteria in the mouse model of melanoma. Mice fed probiotics had a reduced response to treatment with anti-PD-L1 drugs and developed larger tumors than control mice. Further analysis showed that mice fed probiotics had lower levels of tumor-killing immune cells, suggesting a weakened immune response.
In the human study, nearly one-third of the patients reported they had taken a probiotic supplement within the past month. Although the researchers noted that the small sample size and variety of probiotics used by the patients made it difficult to draw definitive conclusions about the association between probiotic use and response to immune checkpoint blockers, they did observe that patients who consumed the highest levels of dietary fiber with no probiotic use survived the longest.
“The impact of dietary fiber and probiotics on the gut microbiota is only part of the bigger picture,” Dr. Trinchieri cautioned. “Many factors can affect the ability of a patient with melanoma to respond to immunotherapy. However, from these data, the microbiota seems to be one of the dominant factors. The data also suggest that it’s probably better for people with cancer receiving immunotherapy not to use commercially available probiotics.”
He noted that larger studies are warranted and should include cancers other than melanoma.
Immune checkpoint inhibitors (CPI) constitute one of the major advances in immunotherapy and cancer treatment [1]. This family of drugs is directed against immune checkpoints, such as programmed cell death 1 (PD-1), PD ligand 1 (PD-L1) and cytotoxic T-cell lymphocyte-associated protein (CTLA-4) [2] (Figure 1).
These checkpoints are part of a delicate system of stimulatory and inhibitory proteins that tightly control the T- cell immune response through the regulation of cytotoxic T-lymphocyte activation, maintenance of self-tolerance, prevention of autoimmunity, and adjustment of the duration and amplitude of the immune response in order to reduce tissue damage during a period of inflammation [3,4,5,6]. Nonetheless, tumor cells have been shown to exploit these inhibitory checkpoints in order to evade the immune system [7].
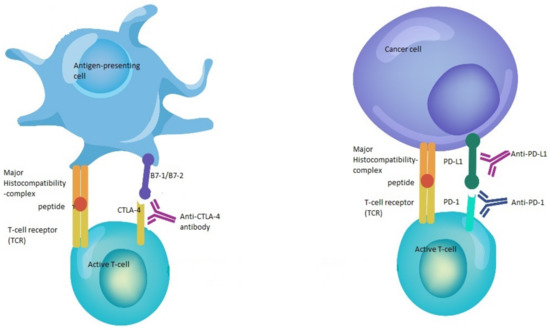
Figure 1. Mechanisms of action of immune checkpoint inhibitors. The binding of B7-1/B7-2 to CTLA-4 keeps the T cells in the inactive state so that they are not able to kill tumor cells in the body. Blocking the binding of B7-1/B7-2 to CTLA-4 with anti-CTLA-4 antibody allows the T cells to be active and to kill tumor cells. In the same way, the binding of PD-L1 to PD-1 prevents the T cells from killing cancer cells.
The interruption of this binding using anti-PD-1/PD-L1 enhances the ability of T cells to kill tumor cells.Several CPIs have been approved for use in clinical practice, demonstrating prolonged overall survival (OS) and improved safety profile in cancer patients when compared with other treatments in various types of tumors, such as metastatic melanoma, non-small cell lung cancer (NSCLC), renal cell carcinoma (RCC) and urothelial carcinoma [3,4,8,9,10].
However, despite these encouraging findings, the success rate of CPIs remains limited, with only a fraction of patients with advanced disease having long-term benefit [7,11]. As such, and given the significant financial cost associated with novel CPIs, it is of great interest to identify patients who are most likely to benefit from these therapies by establishing biomarkers that can predict a positive response to treatment and a durable clinical benefit.In this regard, a high tumor mutational burden (TMB) has been identified as an important biomarker associated with an improved response rate and survival benefit from PD1/PD-L1 blockers [12].
In addition, tumors with defective DNA repair mechanisms have a higher probability of benefiting from CPIs [13]. Moreover, tumor microenvironment (TME) has been shown to play a role in the response to treatment with CPIs. By definition, TME is a complex network of cells that surrounds tumor cells, modulates the innate and adaptive immune response, and affects tumor growth in response to treatment [14,15]. Several factors were shown to directly and indirectly modulate the TME. For example, alterations in energy metabolism appear to influence immune cells in the TME.
This is due to the fact that cancer cells increase the consumption and consequently, limit the nutrient abundance for immune-infiltrating cells, impairing effector T-cell activation and stimulating regulatory immune cells instead, resulting in CPI resistance [16,17]. In addition, obesity, gender, diet and smoking habits were also shown to influence the TME and consequently, impact the response of patients with cancer to CPIs [18,19,20,21,22,23,24].
More recently, the gut microbiota has been surfacing as a potential predictor of response to CPIs [25]. Studies have demonstrated that gut microbiota plays a major role in immunosurveillance and positively impacts the efficacy of CPIs [26,27,28,29,30].
In this review, we will talk about the role of microbiota in cancer pathogenesis and immune-system activity. We will also discuss the preclinical and clinical studies that increased our understanding about the role of microbiota in influencing the response to treatment with immune checkpoint inhibitors, focusing on the mechanisms through which this microbiota could help in overcoming the witnessed resistance to immune checkpoint inhibitors.
Role of Gut Microbiota in Cancer Development and Immunosurveillance
The microbiota of the human body includes a diversity of bacteria, viruses, fungi, and protozoa that usually exist on the different epidermal and mucosal surfaces of the body. These microbiota are essential elements of human health, as they play an important role in the control of different systemic functi
ons [31]. They mostly exert their effect by inducing the synthesis of short chain fatty acids (SCFA) from dietary fibers, as well as different vitamins, including vitamin B and vitamin K [32]. In addition, they also work on the breakdown of multiple complexes, such as sterols and xenobiotics, consequently boosting the immune system [32].
It was described that microbiota plays an important role in cancer, liver diseases, obesity, and neuropsychiatric disorders [31]. In fact, many infectious organisms were found to be causative elements in the development of different cancers. For example, Helicobacter pylori was associated with gastric cancer development [33].
In addition, the DNA of Fusobacterium nucleatum was detected in the tumor cells of colorectal adenomas and cancers that work through the Wnt signaling cascade [34]. This correlation is explained by a pathogen-induced alteration in the host environment that would facilitate the path of the host cells to becoming neoplastic [35].
However, the association between the microbiota and cancer development varies among organs [35]. Below is a list of mechanisms through which the microbiota influences immunosurveillance and carcinogenesis (Figure 2).
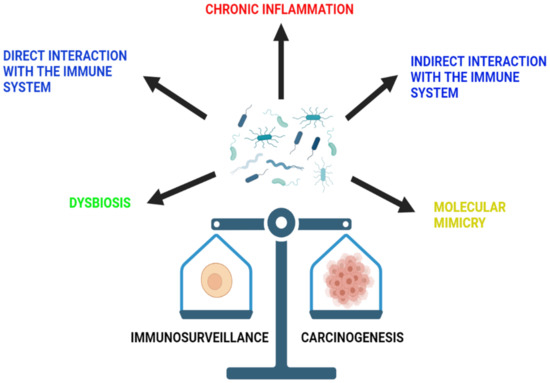
Figure 2. The role of microbiota in immunosurveillance and carcinogenesis. The microbiota of the gut is able to influence immunosurveillance and carcinogenesis through a variety of mechanisms. These include chronic inflammation, dysbiosis and direct and indirect interaction with the immune system, as well as molecular mimicry.
Dysbiosis
Dysbiosis is characterized by the disruption of gut microbial homeostasis, leading to an imbalance in microbiota diversity, composition, function, distribution, and activity [36]. This dysbiosis was shown to be associated with different mutations in the host’s genes, consequently influencing the immune system by altering innate immunity [37].
In addition, dysbiosis of the gut microbiota was clinically linked to the development of many types of cancer, including colorectal cancer (CRC) [38].It is interesting to know that dysbiosis is not only induced by pathogenic organisms [39]. Aging, antibiotics, xenobiotics, smoking, hormones, and dietary elements can also lead to this disruption and consequently, constitute risk factors for CRC [39].
It is also noticed that factors promoting inflammation through different genetic defects affecting epithelial, myeloid, or lymphoid components of the intestinal immune system are carcinogenic through the induction of dysbiosis [39]. Thus, nowadays we know that tumor driver mutations can be regulated by the microbiota [40].
Chronic Inflammation
Another mechanism through which microbiota can induce carcinogenesis is the induction of chronic inflammation. Inflammation plays an important part in the pathogenesis of cancer [40]. A chronic inflammatory state exists in multiple conditions, including inflammatory bowel disease, pancreatitis, and chronic atrophic gastritis, and was linked to the development of cancer through several mechanisms [41].
In the same way, inflammation caused by microbes also favors carcinogenesis [40]. Examples include H. pylori as a causative agent of gastric cancer and Schistosoma haematobium infection as a risk factor for bladder cancer [40].
Direct Interaction with the Immune System
More evidence is coming up to emphasize the ability of the microbiota to anticipate malignancy, manipulate the reaction to immunotherapy and even correlate with survival outcomes in certain cancers [40]. In the 19th century, it was shown that chicken sarcoma virus was able to cause sarcoma in healthy chickens [42].
This was the first proven correlation between the host microbiota, immune system, and cancer [40]. Germ-free animals were the first to be used to investigate this connection [40]. These models lack mucosal immunity because of deficits in Peyer’s patches, mesenteric lymph nodes, lymphoid follicles in the lamina propria, pattern recognition receptors (PRRs), such as Toll-like receptors (TLRs), and the excessive activation of anti-inflammatory T helper (Th) type 2 cytokines [43].
With microbiota implantation in these models, such deficits were replaced, consequently helping in the development of an innate and adaptive immune system [40]. In fact, IL-4 and TGF-ß promote the differentiation of a subset of CD4-T cells called T-helper (Th) 9 cells and induce the secretion of IL-9, which is a key element in antitumor immunity [40]. It was noticed that the GF models lacked Th9, which wereinduced and restored after the implementation of microbiota [44].
One of the major microbiota that was noticed to increase this CD4+ maturation in GF mice was Bacteroides fragilis [40]. Similarly multiple microorganisms were noticed to induce IFNg+ CD8 T-cell3 [40]. Moreover, the specificity and diversity of B cell collections in GF mice was also shown to be determined by the site of microbial exposure [40].
IgA production was mainly induced by transient mucosal exposure, whereas IgM and IgG depended on systemic exposure so they can be able to face invasive infections [45]. This shows that each immune system of each host is unique and influenced by their microbiota exposure [40]
This correlation between microbiota, immune system and carcinogenesis was further investigated with time. The microbiota was proven to induce carcinogenesis through the direct interaction with immune cells. For example, Fusobacterium promotes the growth of colorectal cancer through variable direct mechanisms, including the suppression of the immune system. This is possible through the inhibition of T-cell activity and NK cell cytotoxicity and the induction of the activity of myeloid suppressor cells and tumor-associated macrophages (TAMs), in addition to the inhibition of the tumor-infiltrating lymphocytes in MSI-high types of colorectal cancer [46].
Here rises the relationship between the microbiota, the immune system and cancer progression through immune cells interactions [47].The first cells to fight microbes are the dendritic cells that lay in lamina propria [47]. Through their pattern recognition receptors, they recognize microbes and present them to the adaptive immune system through antigen-presenting cells [47].
Gut microbiota tend to produce butyrate, which in turn can inhibit the dendritic cells’ antigen presentation, consequently decreasing the CD8+ T-cell response [47].Natural killer T-cells, which also carry a cytotoxic activity as part of the innate immune system, are also influenced by the gut bacteria.
In fact, by inducing the metabolism of primary bile acids into secondary bile acids, gut microbiota is able to inhibit the immune-system surveillance of liver tumors by decreasing the CXCR6+ NKT cells through the pattern of C-X-C motif chemokine ligand -16(CXCL)-16 [48].In addition, tumor growth and micrometastasis can also be induced by TAMs [49]. M2 polarized macrophages aim to suppress the TME through the secretion of chemokines and cytokines [49].
Through dysbiosis, discussed above, and through Cathepsin K (CTSK)-mediated TLR4 signaling, microbiota can induce the proliferation of this M2 phenotype of macrophages and consequently, induce tumor growth and metastasis [50]. Following the same pathway, the intra-tumoral bacteria also provoke the proliferation of the M2 phenotype through the TLR2 and TLR5 in pancreatic ductal adenocarcinoma (PDAC) [50].
Through the TLR5 pattern and mainly through the increase in systemic IL-6 secretion, bacteria were able to provoke malignant progression, mainly at extra mucosal sites [51]. This is how Fusobacterium plays a role in colorectal cancer progression. In fact, this species can induce the suppression of the T-cell receptors, as previously discussed [52].Furthermore, follicular T helper cells (TFH) mainly reside in the mucosal lymphoid tissue and function through IL-1R and IL-12 to decrease tumor size [52].
The microbiota in the ileum can have variable effects in this regard, with some acting as stimulators for TFH cells and others as tolerogenic yet not as tumorigenic [52]. In addition, gut microbiota is important for the development of IL-9-secreting cells, which play an important role in decreasing tumor growth [52].Moreover, CD4+ and CD8+T cells, which tend to secrete interferon (INF)-γ, can be specific for microbial epitopes and consequently, may lead to an increase in INF- γ and a decrease in IL-10 and IL-17 upon microbiota exposure modulating the immune response [46]. γδ T cells are a subset of T cells that do not depend on MHC molecules for antigen presentation, yet they express TCRs and consequently, they are able to induce an immune response [53]. The microbiota was shown to induce the production of Vγ6+ Vγ1+ γδ T cells through the Myd88 signaling pathway, provoked by IL-17 production, and hence activate the immune system in the TME [53].
Indirect Interaction with the Host Cells and Immune System
In addition to the direct interaction mentioned above, an indirect interaction also allows the microbiota to influence the immune response and cancer progression (Figure 3).
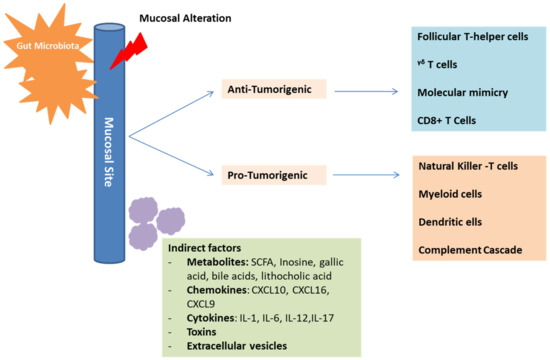
Figure 3. Mechanisms of gut microbiota in cancer development and immunosurveillance. The gut microbiota plays an essential role in influencing the immune-system composition and function. This is possible through direct, as well as indirect interactions with the immune-system components. These interactions can have positive anti-tumorigenic effects on some cells and negative immunosuppressive pro-tumorigenic effects on other cells, depending on the established microbiota profile. Mediators of indirect interactions include metabolites, chemokines, cytokines, toxins, and extracellular vesicles.This is possible through different mediators [29]. For example, Escherichia coli can produce a product called colibactin, which in turn can induce the alkylation of the DNA on the adenine residue, inducing double-strand breaks [54]. This will provoke a carcinogenic effect similar to that found in colorectal cancer [54]. In addition, there is an emerging evidence indicating that, through the microbial metabolite gallic acid, the microbiota can harbor oncogenic properties of genetic mutations similar to Tp53 [55]. Other metabolites that harbor oncogenic properties as well include lithocholic acid, SCFA, cadaverine and de-conjugated estrogens, which mainly contribute to the pathogenesis of breast cancer [56]. As for SCFA, they tend to inhibit the histone deacetylase (HDAC), which plays a role in regulating the innate immune system, controlling myeloid cell differentiation, and modulating inflammatory responses controlled through TLR- and INF- gene expressions [56]. In fact, it was shown that the stools of patients responding to immunotherapy have a good amount of SCFA in contrast to those showing no response [57]. This can emphasize the fact that the inhibition of HDAC can play a role in the upregulation of the expression of the PDL-1 and PDL-2 and thus, work in line with anti-PDL1 therapy [56]. In addition, the butyrate limits CD80/CD86 regulation and thus, increases the effectiveness of the anti-CTLA-4 treatment [58]. As a secondary bile acid, deoxycholic acid is also incorporated in the pathogenesis of hepatocellular carcinoma [40]. Moreover, lipoteichoic acid, coming mainly from gram-positive bacteria, activates the production of TLR-2 dependent prostaglandin E2 that aims at suppressing the immune system in the TME [59]. Inosine, a purine metabolite mainly secreted by Bifidobacterium pseudolongum, Lactobacillus johnsonii and Olsenella species was also found to play an important role in immune checkpoint blockade [60]. Inosine and its metabolite hypoxanthine translocate into the systemic circulation and provoke the differentiation of the Th1 cells through the inosine-A2AR-cAMP-PKA pathway [60]. It also functions as a carbon source for T-cells supporting their differentiation and proliferation and consequently, enhances the response to CPIs [40,60]. Mannose binding lectin (MBL), a metabolite of the Malassezia species, leads to the activation of complement C3 and provokes an oncogenic effect [61]. Finally, another mode of indirect interaction between the host and the gut microbiota takes place through extracellular vesicles. These vesicles mainly send signaling molecules, metabolites or other antigenic proteins that would induce an anti-inflammatory response or a pathological process [62].
Molecular Mimicry
Molecular mimicry (or direct antigenicity) is also an important mechanism through which the microbiota influences immune-system surveillance and potentially contributes to carcinogenesis [29]. It was shown that multiple microbial epitopes are similar either to host antigens or to tumoral antigens, impacting the development of autoimmune diseases, as well as the development and treatment of different malignancies [29,40]. This cross-reactivity affects the efficacy of anticancer immunotherapy, as the CD4+ and CD8+ T cells that are specific for the microbial epitopes are important for the effectiveness of anti-PD1 and anti-CTLA4 treatments [29].
reference link : https://www.mdpi.com/1422-0067/22/15/8036/htm
More information: Christine N. Spencer et al, Dietary fiber and probiotics impact the gut microbiome and melanoma immunotherapy response, Science (2021). DOI: 10.1126/science.aaz7015. www.science.org/doi/10.1126/science.aaz7015



{kind=link}