









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



New research in mice has identified neurons in the brain that influence competitive interactions between individuals and that play a critical role in shaping the social behavior of groups.
“Social interactions in humans and animals occur most commonly in large groups, and these group interactions play a prominent role in sociology, ecology, psychology, economics and political science,” says lead author S. William Li, an MD/Ph.D. student at MGH.
“What processes in the brain drive the complex dynamic behavior of social groups remains poorly understood, in part because most neuroscience research thus far has focused on the behaviors of pairs of individuals interacting alone. Here, we were able to study the behavior of groups by developing a paradigm in which large cohorts of mice were wirelessly tracked across thousands of unique competitive group interactions.”
Li and his colleagues found that the animals’ social ranking in the group was closely linked to the results of competition, and by examining recordings from neurons in the brains of mice in real time, the team discovered that neurons in the anterior cingulate region of the brain store this social ranking information to inform upcoming decisions.
“Collectively, these neurons held remarkably detailed representations of the group’s behavior and their dynamics as the animals competed together for food, in addition to information about the resources available and the outcome of their past interactions,” explains senior author Ziv M. Williams, MD, a neurosurgical oncologist at MGH.
Manipulating the activity of these neurons, on the other hand, could artificially increase or decrease an animal’s competitive effort and therefore control their ability to successfully compete against others. “In other words, we could tune up and down the animal’s competitive drive and do so selectively without affecting other aspects of their behavior such as simple speed or motivation,” says Williams.
The findings indicate that competitive success is not simply a product of an animal’s physical fitness or strength, but rather, is strongly influenced by signals in the brain that affect competitive drive. “These unique neurons are able to integrate information about the individual’s environment, social group settings, and reward resources to calculate how to best behave under specific conditions,” says Li.
In addition to providing insights into group behavior and competition in different sociologic or economic situations and other settings, identifying the neurons that control these characteristics may help scientists design experiments to better understand scenarios in which the brain is wired differently.
“Many conditions manifest in aberrant social behavior that spans many dimensions, including one’s ability to understand social norms and to display actions that may fit the dynamical structure of social groups,” says Williams.
Original Research: Closed access.
“Frontal neurons driving competitive behaviour and ecology of social groups” by S. William Li et al. Nature
Neural Bases of Social Network Structure
Previous studies have employed various metrics to describe the social network structure, such as the number of regular relationships that a person maintains over a 7-day or 1-month period, reflecting the network size (Bickart et al., 2011, 2012), or the number of different types of these relationships to which an individual belongs, reflecting network complexity (Cohen et al., 1997; Bickart et al., 2011, 2012).
Network size and complexity are focused on the individual egocentric network, in which only direct social ties to the focal individual are involved. Recently, an increasing number of studies has started to focus on both direct and indirect social ties from a sociocentric perspective (Smith and Christakis, 2008).
The sociocentric network is constructed from all relationships between every two members in a specified group, such as a class, club, or town. Specifically, the number of outgoing ties (out-degree centrality), incoming ties (in-degree centrality), and the proportion at which an individual frequently lies on the shortest path between any other pair in the group (betweenness centrality) are usually used to reflect the whole structure of the network (see Figure 1).
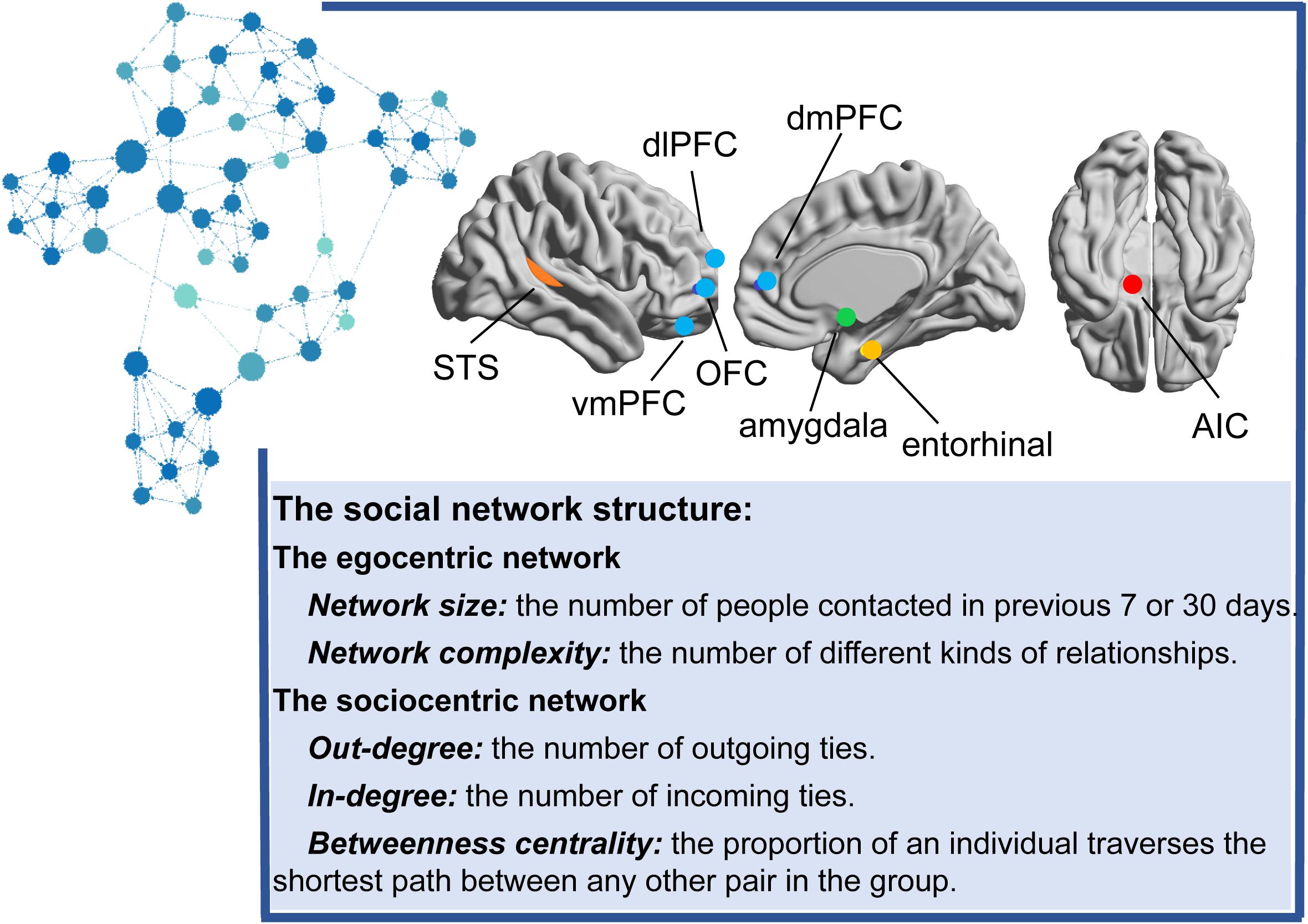
Another important feature of living in a social group is that individuals adapt and utilize their different social status/hierarchies to acquire relative social resources. Social status forms based on genetic or cultural background and plays a key role in determining the relationships among the group members (Dávid-Barrett and Dunbar, 2012). An individual’s social status can affect how people navigate their social world, including effects on their social cognitive processes or social contacts (Muscatell et al., 2012). In contrast to the social network indicators mentioned above, which mainly focus on the roles of social integration (e.g., the degree to which people are embedded in social relationships), social status emphasizes the role of people’s social power in controlling the flows of information and the access to social resources through social relationships (Hussong et al., 2019). However, social status and social network structure are often highly and positively correlated, and they are different dimensions of social position (Hussong et al., 2019; Rueden et al., 2019). For example, low-status individuals may have high-quality but fewer social relationships, whereas relatively high-status individuals usually engage in fewer close relationships but have more social relationships (Berkman et al., 2000; Kraus and Keltner, 2009). Thus, social status is important to consider as a part of the social network structure. However, social status is not a typical index of social network analysis. Because it differs from social network centrality, which is a more objective measure acquired from an individual’s actual social relationships, social status is more like an intrapersonal indicator that is usually measured with a subjective questionnaire, socioeconomic status (SES) (e.g., educational attainment and occupational status) or performance ranking in virtual competition scenarios in human studies and measured less frequently using peer nomination or peer ranking (Hu et al., 2016; Okamoto et al., 2017; Rueden et al., 2019). In the present study, we review the social network and social status studies to obtain a deeper understanding of social context.
Remaining social roles in the network depend on the capacity of social cognition, including the identification of social emotion, understanding of social cues, and theory of mind. Evidence from social neuroscience has suggested that the volume and activities of brain regions (e.g., amygdala and mentalizing network) involved in social cognition processes are also related to the social network structure.
Amygdala
The amygdala, a core brain region involved in the emotional network, plays an extremely prominent role in processing and handling social information such as emotion-related social signals, social values, motivation, and identity (Adolphs, 2001; Gothard et al., 2018). +The earliest study focusing on the relationship between the amygdala and social behavior used primates as study subjects and found that after amygdalectomy, the individuals who formerly held a high social status in the group had a decreasing social rank and became extremely compliant (Rosvold et al., 1954; Watanabe and Yamamoto, 2015). Accordingly, Munuera et al. (2018) further reported that the primate amygdala encodes the hierarchical rank and reward value.
Similar to primate research, a growing body of literature on humans has also shown that the amygdala is associated with rank in one’s social network. Bickart et al. (2011) examined the total number of regular contacts that a person maintains (social network size) and the number of embedded networks (social network complexity). They reported that these two social network indexes positively predicted the amygdala volume. Likewise, Kanai et al. (2012) tested a larger sample and found that the gray matter density in the amygdala correlated with both online and real-world network sizes, whereas the gray matter density in the right superior temporal sulcus (STS), left middle temporal gyrus and entorhinal cortex was specifically associated with online social network size.
In another study using three social network indicators to further confirm the relationship between the amygdala and social network structure, researchers found that the online network size (the number of Facebook friends), offline network size (the number of regular contacts of an individual in the last 30 days), and social support network size (the number of friends who could provide social support) were positively correlated with the gray matter density of the amygdala and the right entorhinal/ventral anterior temporal cortex, regardless of whether the individuals were in a resting state or viewing the faces of friends or strangers (Heide et al., 2014).
The important role of the amygdala in social network construction and maintenance has also been confirmed by Jones et al. (2020). According to the authors, the amygdala is presumed to track visual signals in social interactions, such as face stimuli, gestures, and expressions (Bickart et al., 2011, 2012). A larger amygdala provides an individual with advantages in processing non-verbal social signals (Bickart et al., 2011, 2012). An amygdala with a larger volume and higher gray matter density enables an individual to search, decode, and match multilayered and complex social signals for processing more complex social network information (Bickart et al., 2011, 2012). In addition, the amygdala tracks the reward value brought by social interaction. Individuals with a larger volume, higher gray matter density, or higher activation level of the amygdala tend to perceive social interaction as more interesting and of higher reward value, which in turn prompts them to develop more social connections (Bickart et al., 2012; Zerubavel et al., 2015; Liu et al., 2019).
Brain structure and function are not independent of each other. Instead, structural changes in the cerebral cortex usually cause corresponding functional changes (Falk and Bassett, 2017). Therefore, a number of studies searched for more evidence at the level of brain functional connectivity to determine the importance of amygdala networks in social network construction. The functional connectivity between the amygdala and orbitofrontal cortex (OFC) is crucial for facial expression recognition, social strategy development, social reward processing, prosocial behavior, etc. (Hampton et al., 2016; Kwak et al., 2018).
Researchers generally agree that amygdala-OFC functional connectivity stably and positively predicts the differences in social network size among individuals (Hampton et al., 2016; Kwak et al., 2018). According to a previous study, smell is a type of social signal that conveys information about an individual, such as sex, disease and emotional state; therefore, individuals with high olfactory sensitivity are able to identify social signals from the body odor of others, which is conductive to social interaction (Zou et al., 2016). Individuals with higher functional connectivity between the amygdala and OFC, which are the brain regions related to olfactory sensitivity and theory of mind, are more sensitive to olfactory signals and have a larger social network size (Zou et al., 2016).
Furthermore, Bickart et al. (2014) divided the amygdala into three subareas and investigated the relationships between the social network size and the intrinsic anatomical connectivity levels of the three subareas with other brain regions. The functional connectivity levels of the perception network, which consists of the ventrolateral amygdala and OFC and is responsible for decoding social signals, and the social affiliation network, which consists of the medial amygdala and ventromedial prefrontal cortex (vmPFC) and is responsible for processing social reward information, are positively correlated with social network size.
In addition, the functional connectivity level of the aversion network, which is composed of the dorsal amygdala, insula and hypothalamus and is responsible for processing aversive stimuli, is not significantly correlated with social network size (Bickart et al., 2012, 2014). This finding suggests that the amygdala does not work independently. Instead, it works synergistically with other brain regions to enhance an individual’s ability to process social information. However, the perception network, the social affiliation network and the social aversion network were defined by a data-driven approach, and the intrinsic functional connectivities among them have not been verified in further studies (Bickart et al., 2012).
A relatively stable correlation has been identified between social networks and the gray matter structure and activation level of the amygdala, a region of the brain that has attracted attention from researchers for many years. The aforementioned studies support the hypothesis that the volume and activation level of the amygdala may affect the social function and social network structure of human individuals by affecting their abilities to process social signals and perceive social reward value.
The amygdala comprises multiple nuclei that are distinct in morphology, histochemistry, cytoarchitecture, and functional connectivity (Kedo et al., 2018; Jones et al., 2020). Jones et al. (2020) further found that the central nucleus, basal and accessory basal nuclei of the basolateral amygdala are significantly associated with social network size, but no association with the lateral amygdala nucleus was found. However, the participants in this study were homeless and precariously housed people, not the general population. In addition, the social network indicators of this study are based on a sociocentric network, which is different from the studies described above that focused on an individual’s egocentric network.
The egocentric network focuses on the number and types of all direct social relationships associated with the core individuals, while the sociocentric network focuses on the social position of individuals in a specific collective based on both their direct and indirect relationships. The egocentric network and sociocentric network describe the network structure from micro level and macro level, respectively. Therefore, these two different network indicators may reflect different brain areas and brain activities. Future studies should explore the sociocentric network to supplement the neural basis of social network at the macro level.
Indeed, some studies failed to replicate the important role of the amygdala in a social network (Lewis et al., 2011; Powell et al., 2012; Noonan et al., 2018; Spagna et al., 2018). According to the authors, social networking is complicated and depends more on social cognition abilities, which may be related to frontal and temporal structures, rather than to non-verbal social signal identification. These studies did not obtain consistent results for the amygdala, possibly because they assessed the social network index using distinctly different methods. Thus, the relationship between the amygdala and the social network must be verified in further studies using consistent measurements and populations. In addition, the studies described above discussed only resting-state networks, and therefore, more studies are needed to clarify the causal relationship among the amygdala, social function, and social network structure in the future.
The Mentalizing Network
The amygdala network is involved in the perception and understanding of non-verbal social signals. However, the social cognitive ability required for establishing and maintaining social networks is beyond the superficial processing of social signals; instead, the deep processing of the psychological state of other people is also key to successful social interactions (Kanai et al., 2012). Premack and Woodruff (1978) proposed the concept of “theory of mind” to describe an individual’s social cognitive ability, which refers to an individual’s ability to infer the personality characteristics, psychological state, and intentions of other people.
Studies have found that the core brain regions involved in theory of mind include the medial prefrontal cortex (mPFC), the temporal-parietal junction (TPJ), the posterior cingulate cortex (PCC), the medial OFC (mOFC), and the precuneus, which are commonly known as the “mentalizing network” (Mitchell, 2009; Muscatell et al., 2012). According to these findings, the network structure of an individual is postulated to be limited by mentalizing competence. In recent years, researchers have also begun to explore evidence supporting the hypothesis that theory of mind reflects the structure of an individual’s social networks from the perspective of the mentalizing network.
Among the core brain regions in the mentalizing network, the covariation in the PFC and social network structure have been emphasized in most studies. The PFC is the core brain region in the mentalizing network. The vmPFC and OFC are involved in the emotional part of the theory of mind and are mainly responsible for understanding the emotional state of others (Abu-Akel and Shamay-Tsoory, 2011). The dorsomedial PFC (dmPFC) and the dorsolateral PFC (dlPFC) participate in the cognitive part of the theory of mind and are mainly responsible for inferring the beliefs and intentions of others (Abu-Akel and Shamay-Tsoory, 2011).
Existing studies have revealed that both emotional and cognitive components are reflected by the social networks of individuals. Lewis et al. (2011) have found that individuals with a larger social network size in the previous 30 days understand the psychological state of others more accurately and generally have larger gray matter volumes in the mOFC and vmPFC (Kanai et al., 2012). In another study, researchers further explored the possible causal relationship among social network size, theory of mind, and the PFC in human individuals using a path analysis. They found that theory of mind plays an intermediary role between the OFC volume and social network size. Individuals with larger OFC volumes have higher mentalizing competence and thus more complex social relations (Powell et al., 2012). Kwak et al. (2018) constructed sociocentric networks with town as the unit and found that the volumes of the dmPFC, OFC, and TPJ were generally larger in the individuals with higher in-degrees. Despite the discrepancy in results, all the aforementioned studies emphasize the significant role of the OFC. Compared with the cognitive component of social cognition, the emotional component may be more important for social network construction and maintenance in humans. However, this hypothesis should be confirmed by performing additional research.
Researchers have explored the relationship between mentalizing network functional activity and social networks. A study conducted by Pillemer et al. (2017) included elderly populations. The elderly participants were asked to report the number of social ties with high contact frequency (network quality) and the number of social ties that the participant recently contacted (network quantity). The researchers found that the functional connectivity level between the frontal and parietal lobes, mainly including the dmPFC, dlPFC, PCC, and precuneus, was positively correlated with the two social network indicators listed above.
Network quality was positively correlated with the functional connectivity level of the lateral part of the frontoparietal network, and network quantity was positively correlated with the functional connectivity level of the medial part of frontoparietal network (Pillemer et al., 2017). Noonan et al. (2018) also found that the functional connectivity levels between dmPFC and dlPFC with the anterior cingulate cortex (ACC) predict the social network size of an individual (Noonan et al., 2018).
In another study on elderly individuals, those with greater functional connectivity between the frontal and temporal lobes were shown to have larger social networks. Frontotemporal functional connectivity is closely related to an individual’s ability to perceive the agreeableness of surrounding people, suggesting that individuals with larger social networks may perceive greater interpersonal closeness (Bang et al., 2019). Thus, although some brain regions play important roles in social behavior, extensive brain functional connectivity is likely more important for social network construction and maintenance.
Theory of mind is the basic mechanism through which individuals successfully establish social relationships. By examining the relationship between social networks and the core brain regions of the mentalizing network, researchers have shown that social networks and theory of mind share the same neural circuits. However, Bickart et al. (2012) found that the functional connectivity of the mentalizing network, including the dmPFC, precuneus and TPJ, was not related to either the social network size or complexity, and the authors speculated that the contribution of the affective processing by the amygdala to social networking was greater than social cognition (Bickart et al., 2011, 2012). Nevertheless, they could not ignore the important role of the mentalizing network. The authors emphasized that the functional connectivity between the amygdala and the vmPFC and OFC predicted social network structure.
The Mirror Neuron System
The mirror neuron system, another important neural system involved in social cognition, is also related to social network structure. The mirror neuron system is mainly responsible for supporting imitation and understanding other people’s actions (Ikeda et al., 2019). Brain regions such as the inferior frontal gyrus (IFG), inferior parietal lobule (IPL), and STS are involved in the mirror neuron system. However, the identification of biological motion supported by the mirror neuron system is a basic and vital function for individuals’ sociality, and only a few network neurosciences studies have focused on the mirror neuron system. They all emphasized the association between social network properties and the posterior STS (pSTS).
The pSTS is specialized for understanding and imitating the non-verbal social signals of others, such as body movements, eye gaze, and mouth movements (Deen and Saxe, 2019). Researchers reported a positive correlation between the online network size (number of friends on Facebook) with the pSTS gray matter density, and real-world social network size did not have a significant relationship with the pSTS (Kanai et al., 2012).
Other studies were based on task-state functional magnetic resonance imaging (fMRI) when identifying biological motion. Dziura and Thompson (2014) investigated the effects of the social network size and complexity on the perception of basic, non-verbal social stimuli (e.g., gestures and expressions) and found that the activation of the STS and amygdala induced by biological motion recognition was positively correlated with social network indicators. Based on this result, the individual at the center of a network must have a strong ability to identify obscure social signals in order to play a complex social role (Dziura and Thompson, 2014).
Accordingly, Kirby et al. (2018) investigated neural sensitivity to biological motion related to social network properties in middle childhood. Consistent with the study by Dziura, the children’s social network size (the number of people the child regularly sees or talks to), diversity (the number of social roles the child plays) and embeddedness (the number of social domains in which the child is active) predicted activation in the pSTS, which significantly correlated with the neural sensitivity to biological motion when the children viewed biological motion.
Surprisingly, few social network neuroscience studies emphasize the function of the mirror neuron system in social networking. However, the existing research has provided evidence that the ability to construct and maintain social networks is limited by neural sensitivity to biological motion. Little direct evidence is available on how brain structure and functional connectivity among regions in the mirror neuron system influence social networking. This influence is important because the ability to understand biological motion, including facial expressions and body movements, can affect an individual’s social skills and the quality of social interaction (Oberman et al., 2007). Recognizing and clarifying the relationship between the mirror neuron system and the social network (not only social cognition) might improve our ability to predict group behaviors (Freiwald, 2020). Thus, research in this area is needed for further and deeper explorations of the mirror neuron system and social networks in the future.
Other Brain Regions
In addition to the involvement of the amygdala, the mentalizing network and the mirror neuron system in social network construction and maintenance, previous studies have found that the entorhinal cortex and anterior insular cortex (AIC) are also closely related to the social network structure in humans. By comparing two different samples from New York and Beijing, researchers found that social network size and complexity positively correlate with AIC volume (Pillemer et al., 2017; Spagna et al., 2018). One of the important roles of the AIC is to process interpersonal emotional information, including sympathy, empathy and understanding the feelings of others (Pillemer et al., 2017; Spagna et al., 2018). Previous studies also revealed a linear correlation between the online social network size and entorhinal cortex gray matter density (Kanai et al., 2012; Heide et al., 2014). The entorhinal cortex is related to memorizing names and faces, but unrelated to social cognition (Sperling et al., 2001; Kanai et al., 2012). Thus, memory ability is also clearly one of the indispensable abilities for individuals to maintain a large social network. In other words, social cognitive ability is not the sole factor limiting the social network structure of an individual. Accordingly, Joo et al. (2017b) examined the global structure of the sociocentric social network in an entire village and suggested that older adults’ social network embeddedness (number of social groups to which an individual belongs), but not social size, was positively associated with functional connectivity between anterior-posterior regions, including the right inferior frontal gyrus and the occipital lobe (right lateral occipital cortex), the cingulate gyri and parietal (left superior parietal lobule and precuneus cortex) and occipital lobes (right lateral occipital cortex). Older adults with higher network embeddedness may need more cognitive resources to deal with complicated social relationships, which may subsequently activate functional connectivity among anterior-posterior regions.
In summary, social network construction and maintenance involve various related brain regions, including the amygdala network, the mentalizing network, the mirror-neuron system, the entorhinal cortex, and the AIC (see Figure 1). Although the aforementioned studies used different social network indicators and measured the indicators for different periods of time, ranging from 1 week to 1 month, and from an individual egocentric network to a sociocentric network, they reported relatively reproducible findings. However, Lin et al. (2019) repeated previous studies and did not identify a brain structure that was significantly related to social network structure. The authors postulated that the relationship between dynamic indicators of social network structure and brain function, instead of brain structure, should be emphasized because social networks are dynamic, as social relations might naturally increase or decrease over time, but the brain structure is relatively stable (Lin et al., 2019). Although many of the studies described above have discussed the functions of brain regions, they only focused on resting-state networks, and few of them mention how social networks affect the processing of social information by the brain. More in-depth studies should be conducted to investigate task-state networks and to explore and verify the regulatory effect of network structures on neural function.
Brain Connectivity Dynamics During Social Information Processing Reflect the Social Network Structure
The position of an individual in a social network represents the individual’s social resources, preferences, and status. Individuals with larger or more complex networks have more opportunities to influence others or be affected by others, which provides those individuals with abundant social experience that guides them to cope more effectively with problems in different social situations (O’Donnell et al., 2017). Therefore, social network structure may further affect the activation level of the related brain regions during social information processing. In recent years, researchers have used various social cognitive tasks to explore how social networks regulate brain activity in the process of social cognition.
In a study investigating social conformity, O’Donnell et al. (2017) presented a series of software applications to study subjects and asked them to what extent they would recommend the apps to their friends. Additionally, the authors investigated the whole network formed by each individual and his/her friends on Facebook. Compared with the individuals with lower betweenness centrality (individuals with fewer opportunities to pass information and those on the outer edges of the social network), the individuals with higher betweenness centrality exhibited greater activation of the mentalizing network when they disagreed with their peers. Thus, when providing suggestions to others, the individuals in the center of the network care more about other people’s opinions; i.e., they are more sensitive to the opinions of others (O’Donnell et al., 2017).
Researchers asked participants to complete a classic Cyberball social exclusion task and used social network density as a network indicator to evaluate interpersonal communication. Friends of individuals with a higher network density are usually friends with each other. Compared with individuals with a higher network density, the functional connectivity between the left and right TPJ was higher in individuals with a lower network density when they were rejected. The researchers postulated that the individuals with a lower network density thought more about the causes and consequences of rejection and conducted more self-reflection when they were rejected by others, while those with a higher network density were less affected by social exclusion because their close social connections helped them buffer the negative effect of social exclusion (Schmälzle et al., 2017). Based on this observation, social networks affect interpersonal interaction patterns. Nevertheless, social networks and interpersonal interaction patterns might have a mutual relationship, which requires further investigation.
Previous studies emphasized the important role of social experience in processing social reward value. As shown in the study by Meshi et al. (2013), the activation of the ventral striatum when participants gain reputation from others positively predicted the degree of Facebook use (e.g., the number of friends on Facebook or the minutes per day spent on Facebook). Meanwhile, the activity in the ventral striatum during monetary gain did not predict Facebook use. The ventral striatum is a well-established structure involved in processing rewards such as food, money, and social stimuli (Meshi et al., 2013). The social experience/context created by our social network may influence our perception of social rewards based on positive evaluations from others (Fareri and Delgado, 2014).
In a recent study, researchers revealed that social network structure also moderates neural function in a non-social task (Go/No-Go response inhibition task). Adolescents with larger online social networks who are involved in more communities in their social network showed a stronger relationship between response inhibition and functional connectivity in self-referential systems (e.g., vmPFC) and response inhibition systems (e.g., superior temporal gyrus), suggesting that social context might facilitate self-control in adolescents. The real-life social network might have accounted for the compensatory role of social experience for better task performance (Tompson et al., 2020). However, adolescents are more sensitive to social influence than adults. Thus, the compensatory role of social experience in self-control does not extend to adults.
The abovementioned findings lay an important foundation for studies investigating how social network structure relates to neural sensitivity to social information processing. In addition, some studies found that the activation of an individual’s brain regions related to the processing of another person’s information is affected by the position of the other person in his/her social network. For example, using two small groups (13 members) as research subjects, a study based on whole networks found that the activation levels of the dmPFC, precuneus, and TPJ in the mentalizing network increased when the individuals saw the face of a highly popular member in their group, in which the reward system (the vmPFC, amygdala, and striatum) played an intermediary role. In other words, the individuals perceived that the popular member would bring them higher reward value, which in turn prompted them to infer the psychological state of the popular member (Zerubavel et al., 2015). In another study, participants were asked to watch short self-introduction videos of their friends, friends’ friends, and friends of friends’ friends in their whole network. Watching videos of their immediate friends, the participants had high activation levels in the lateral superior temporal cortex (STC), mPFC, and IPL, which are the brain regions generally responsible for processing physical distance and abstract distance (e.g., social distance) information from the individuals themselves. Thus, individuals will spontaneously process information related to the relationship between others and themselves in a network, regardless of whether the task of social distance judgment is required (Parkinson et al., 2017).
Studies that used different social cognitive tasks all confirmed that an individual’s social network structure affects his or her ability to process social information. The individuals at the core of a network have rich social experience and use the mentalizing network differently from those on the edge of the network in social cognitive tasks (Falk and Bassett, 2017). However, these studies only used social network indicators as moderating variables and did not manipulate them. Future research can use social networks as independent or dependent variables for in-depth investigations to further clarify the relationship between social networks and social cognition.
Neural Bases of Social Status
High-status individuals typically have more opportunities to receive primary resources and are engaged in communicating with others for longer periods (Fiske, 1992; Chiao et al., 2009). Navigation of the social world with a higher hierarchical status requires higher level of capacities in expressing and recognizing social status signals, as well as processing cognitive information (Chiao et al., 2009; Hanson et al., 2013). MRI studies have found that a lower SES is associated with decreased gray matter or volumes in prefrontal, temporal, and parietal cortices, the hippocampus and amygdala (Hanson et al., 2013; Noble et al., 2013; Brito and Noble, 2014; Finn et al., 2016; see Willard and Shively, 2016 for review). Researchers have attempted to discover the neurocognitive function of social status using behavioral tasks, including social information processing, working memory, language processing, social information processing, etc. For instance, Muscatell et al. (2012) revealed that college students’ subjective social status in the university community was negatively correlated with neural activity in the dmPFC, mPFC, and precuneus/PCC (posterior cingulate cortex) when processing social information in response to negative feedback. Adolescents with a lower SES showed greater neural activity in the dmPFC and amygdala under social threats. Some other fMRI studies focused on SES revealed similar results. For instance, children and adolescents with a higher SES showed less overall activity in several areas associated with executive function, including the inferior frontal gray and the dorsal anterior cingulate cortex (dACC), when performing tasks related to working memory, mathematics achievement, language processing, etc. (Sheridan et al., 2012; Finn et al., 2016; see Farah, 2017 for review). Individuals with a low social status may require more attention on cognitive information processing, which would be reflected in greater activities in the brain regions discussed above, compared to people with a higher SES. In addition, fMRI studies also reported differences in reward-related brain regions (e.g., ventral striatum and caudate) between individuals with high and low SES, and the latter were more sensitive to reward associated learning and actions (Yaple and Yu, 2019). However, individuals with a low social status do not always display hyperactivity in executive network regions and reward-related regions (Finn et al., 2016; Yaple and Yu, 2019). In another fMRI study, social status was measured by adolescents’ popularity and acceptance in their class. Their popularity and acceptance were analyzed by asking adolescents which classmate they like most and least and which classmate they perceive as most and least popular. Adolescents who were nominated as most likable and popular were central and influential in a peer group with a high social status. Participants’ acceptance was positively correlated with activity in the dorsal anterior cingulate cortex (dACC) during exclusion, while participants’ popularity was positively associated with activity in the ventral striatum and mPFC during exclusion (Water et al., 2017). Therefore, when individuals with different social statuses process information, the social status will be reflected in the activities of different brain regions. The relationship between social status and the activities of these brain regions may be affected by the types of processed information and the measurements of social status. Numerous studies have focused on the neural mechanism of social status, but few directly compare the similar and distinct neural mechanisms between social status and social network. According to the results mentioned above, social status shares overlapping neural representations with social network structure, such as the mPFC and amygdala.
However, much of the work on social status and brain function has focused on SES. Although SES is an objective indicator measured by the income or education level of an individual/family, it does not involve the description of social relationships. SES is quite different from the index of social network structure, making a direct comparison of the neural basis of these two social structure-related indicators difficult. Social status has also been measured using peer ranking or nomination, depending on the social network (Water et al., 2017; Rueden et al., 2019). Future research should increase the exploration of the relationship between the neural mechanisms underlying social network structure and social status, such as whether the correlation between social status and brain activities will be moderated by social network indicators.
reference link :https://www.frontiersin.org/articles/10.3389/fpsyg.2021.626337/full



{kind=link}