










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")


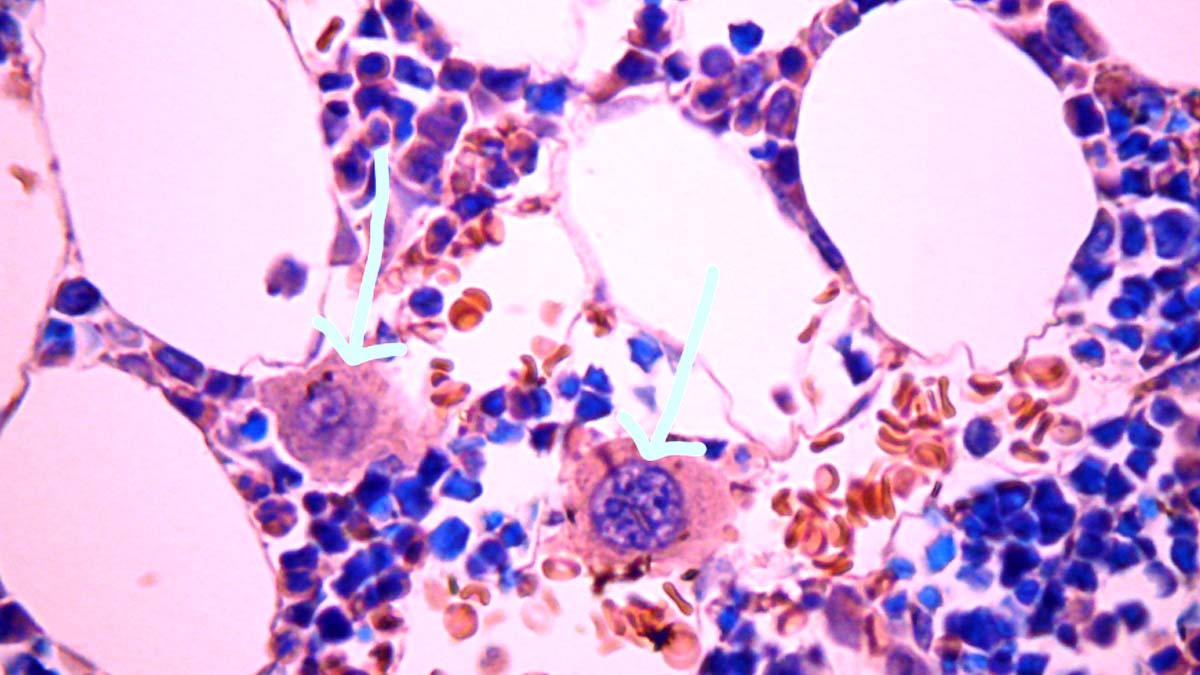
A new study by researchers from Imperial College London, Hammersmith Hospital-London, UK Charing Cross Hospital Campus, UK Chelsea and Westminster NHS Foundation Trust, United Kingdom Health Security Agency, University Hospitals Birmingham and King’s College London, has found that the SARS-Cov-2 proliferates in neurons of the myenteric plexus and also in megakaryocytes.
This discovery has lots of implications in terms of accounting for extrapulmonary involvement, such as in the gastrointestinal tract and nervous system, as well as frequent thrombotic events in SARS-CoV-3 infected individuals.
The study findings were published in the peer reviewed publication: The Journal of Pathology https://onlinelibrary.wiley.com/doi/full/10.1002/path.5878
Predominantly a respiratory disease, it has been logical for COVID-19 histopathological studies to focus on the respiratory system. Despite this, there is value in examining the histopathological correlates of SARS-COV-2 infection in extrapulmonary organs and tissues, given the increasing spectrum of extrapulmonary symptoms reported.
Our results provide evidence for peripheral neuroinvasion in the digestive tract, but critically, not in the CNS. Furthermore, we have demonstrated viral NP within platelet-producing megakaryocytes, a finding that provides insight into the well-characterised thrombotic sequelae of COVID-19.
It had been previously speculated that, due to the expression of ACE2 in neurons of the myenteric plexus, these cells are possible targets of SARS-CoV-2 infection [2], a hypothesis that we corroborate here for the first time. The long-term consequences of our findings in the myenteric plexus are yet to be established.
PD is a neurodegenerative disorder thought to result from the prion-like spread of amyloidogenic alpha-synuclein causing catastrophic motor impairment and other symptoms, such as dementia, as the spread of misfolded protein progresses throughout the brain [25].
However, it is not known if the initial seeding of the misfolded protein conformation is purely a stochastic event or ignited by an internal or external factor, with the exception of 5–10% of cases in which genetic mutations have been shown to impact alpha-synuclein biology [26].
The brain pathology of PD initiates in the olfactory bulb and medulla from where it propagates in a stereotypical pattern [11], and this is anticipated by accumulation of misfolded alpha-synuclein in the ENS. This pattern of pathology progression explains anosmia and gastrointestinal (GI) symptoms such as constipation and nausea [27], appearing early in PD.
It has been proposed that an environmental agent reaches the brain from the nose through the olfactory nerves or from the gut through the ENS via retrograde axonal transport along the vagus nerve reaching the medulla [12, 28].
Viral infection of myenteric neurons, as implied by our study, could in theory be one such environmental factor. Historically, links between viral infection and Parkinsonism were seen following the 1918 influenza–encephalitis lethargica pandemic [29]. During the COVID-19 pandemic, there have been reports of concurrent neurological complications and encephalitis [20], however, there is currently only one report of postviral Parkinsonism development [30].
Diversity of genetic background, comorbidities, and previous infections all impact susceptibility and likely may only have neurodegenerative ramifications in a subset of people. As we begin to recover from the pandemic, we should nevertheless still be conscious of COVID-19 when assessing future neurological cases.
On the other hand, the extent of SARS-CoV-2’s direct invasion of the CNS is uncertain. Throughout the pandemic, neurological symptoms such as headache, ageusia, and anosmia were commonly reported, and the involvement of both the central and peripheral nervous system was hypothesised [31].
Our brain cohort of SARS-CoV-2-infected patients includes patients with clinical and pathological neurological disease, including brainstem encephalitis and cases with no neurological involvement. However, positivity within the brain appeared to be limited to endothelial cells. Crucially, despite observing viral NP in the neurons of the myenteric plexus, we found the origin of the vagus nerve in the medulla oblongata to be devoid of neuronal SARS-CoV-2 NP (supplementary material, Figure S5).
This suggests that, in the event of any neurodegenerative disease developing after SARS-CoV-2 infection, consideration should be given to the spread of misfolded protein or other pathophysiological alterations triggered by the virus in ENS neurons, rather than the direct entry of the virus into CNS cells.
Previous reports about the possibility of viral protein or RNA in the CNS are conflicting [8, 14, 16, 32, 33]. One hypothesis postulates that systemic vascular changes are likely behind reported neurological symptoms, as an indirect consequence of lung-derived hypoxia rather than direct viral damage. Additionally, endothelial cell targeting and injury are also considered to be key elements in COVID-19 pathophysiology that have been frequently alluded to [34].
COVID-19 has been established as a prothrombotic disease characterised by coagulopathy, thrombosis, and platelet activation [35, 36]. Our results parallel this with findings of platelet-producing megakaryocytes colocalising with the viral NP within the bone marrow. Several publications have also reported the presence of circulating megakaryocytes in multiple organs, including lung, heart, kidney, liver, and brain [14, 37-39].
A recent preprint report that infected calprotectin-expressing megakaryocytes additionally express the ACE2 receptor and TMPRSS2 [41]. Even more intriguing, that study also provided evidence of megakaryocytes harbouring infected pro-platelets. Together with our own observations, it is probable that entry occurs by several mechanisms. The principal molecular route of SARS-CoV-2 entry into megakaryocytes and platelets is therefore still unresolved.
Our results imply appreciable involvement of the GI tract and related digestive organs in COVID-19, correlating with the high prevalence of symptoms such as diarrhoea, abdominal pain, anorexia, and vomiting in COVID-19 symptomatology [42]. Of note, the viral cell tropism in the digestive tract overall mirrors that of the expression profile of ACE2 in cell types such as enterocytes [43].
Preexisting chronic liver diseases, such as steatohepatitis and cirrhosis, have been commonly associated with severe cases of COVID-19, with 2–11% of patients having liver comorbidities [44, 45]. With four of ten patients displaying liver disease, this is also a common finding within our own full-body postmortem cohort. In addition to endothelial cells, three cases (PM1, PM6, and PM10) exhibited a striking viral NP presence within parenchymal tissues, including bile duct cholangiocytes, as well as hepatocytes. Studies of the ACE2 receptor in normal liver asserts low expression in hepatocytes; however, mouse models and human samples suggest chronic liver injury may contribute to ACE2 receptor upregulation [43, 46]. Cholangiocytes, on the other hand, purportedly have high levels of ACE2, which may explain the strong viral protein detection seen in this study.
We also observed, predictably, a diffuse viral presence throughout the respiratory system. Consistent with other reports, detection of viral NP in the lungs appears to be limited to seromucinous glands, alveolar pneumocytes, and respiratory epithelium [47]. As previously described, all the lungs in the PM1-10 case series present with DAD, a histopathological aberration commonly affiliated with acute respiratory distress syndrome (ARDS) [8].
However, our immunohistochemical analysis revealed parenchymal positivity in only a subset of patients (four of ten). As NP has been canonised as a proxy for replicating SARS-CoV-2, this suggests that only a few patients in our cohort died with active pulmonary infection [48].
Considering the disease duration of these cases, it is likely that our findings capture the early phase of ARDS in which viral activity persists [49-51]. Similar observations have been made in other SARS-CoV-2 in situ studies, as well as in SARS-CoV, where viral replication in the lungs is constrained to the early postinfective period (~2 weeks) [47, 52, 53].
Interestingly, we found that parenchymal SARS-CoV-2 presence in the lungs was associated in all cases with an extrapulmonary viral presence, and that this was also constrained to this ~2-week period. Our data, with a universal endothelial presence in these cases, suggests that extrapulmonary dissemination could occur via haematogenous spread.
This is supported by increased SARS-CoV-2 RNA load in the blood (RNAemia) to be associated with disease severity in COVID-19 patients [54, 55] and by multiorgan damage to be related to high RNAemia [56], although other studies disagree [57]. In the case of GI viral presence, a faecal-oral route of transmission is possible [58], supported by detection of viral RNA in the faeces of COVID-19 patients, even after testing negative in respiratory tissues in a subset of patients [59].
Our data also suggest that beyond this ~2-week period, COVID-19 multiorgan complications may be the result of persistence of viral tissue damage and of pathophysiological cascades ignited by the viral infection, including inflammatory activation, rather than continued viral infection. Given that our postmortem samples are from patients who died during the initial first COVID-19 wave in the UK, our study cannot comment on any vaccine-related inflammatory processes.
A strength of this study is the strict validation performed on our immunostaining protocol. Consistently throughout our study we identified a characteristic immunostaining pattern to be indicative of SARS-CoV-2. After gaining entry into host cells, viral RNA and replicative protein components form a reticulovesicular network by reorganising the host endoplasmic reticulum [60].
We have identified that, in most cell types, genuine SARS-CoV-2 positive stain displays a cytoplasmic apical or paranuclear punctate pattern reflecting the viral cycle. In addition, the clinical relevance of this antibody is best illustrated by our tracheal biopsy series in which we used our protocol to show clinicopathological correlations.
Out of 15 biopsies, the vast majority of SARS-CoV-2 IHC results were consistent with the timeline of nasopharyngeal swab results, as well as overall symptom presentation. The outliers included one patient who persistently had negative swabs yet showed very strong IHC positivity. As the tracheostomy occurred a considerable time after the last swab, it is likely this patient had been infected during their prolonged admission to the ICU, subsequent to tracheostomy and tracheal biopsy.
Our study has limitations. Despite comprehensive sampling, our full-body cohort is of limited size, with some tissues being subjected to autolysis as well as sampling bias. This makes it difficult to speculate on any enteric neuronal loss or cytopathic damage seen in patients who died later in their disease course, although we did find evidence for at least a proportion of the neurons in the myenteric plexus surviving into the postreplicative phase (supplementary material, Figure S6).
Second, we cannot completely exclude the possibility of cross-reaction with SARS-CoV-1, although, to our knowledge, this virus is not currently in circulation in our community. Contextualising the patients analysed in this study (e.g. date and location of death), it is highly unlikely that we have misidentified another betacoronavirus, particularly as the cases had confirmed SARS-CoV-2 PCR positivity. Additionally, when considering nucleoprotein homology, the closest sequence (bat coronavirus nucleoprotein) only reveals 75% alignment (supplementary material, Figure S7).
Overall, our investigation has demonstrated the multisystemic tropism of SARS-CoV-2. Concordant with other histopathological publications, we have shown extrapulmonary involvement, with an emphasis on the digestive system, in severe cases. Most significantly, we provide evidence of neuroinvasiveness in the ENS but not the CNS. We have also identified megakaryocytes as potential targets of SARS-CoV-2. Further investigations in gene expression profiling related to these findings will help determine any long-term implications.



{kind=link}