











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A new study by researchers from Harvard University has revealed compartmentalized SARS-CoV-2 infection in the human body according to cell and tissue types and with the virus itself evolving and mutating to sustain its existence in these sub-micro environments.
The study findings besides showing the existence of viral reservoirs and also validating viral persistence, also has numerous implications in terms of changing what we know about Long COVID issues and also the dynamics of variant emergence.
The study findings were published in the peer reviewed journal: Nature Communications.
https://www.nature.com/articles/s41467-022-34256-y
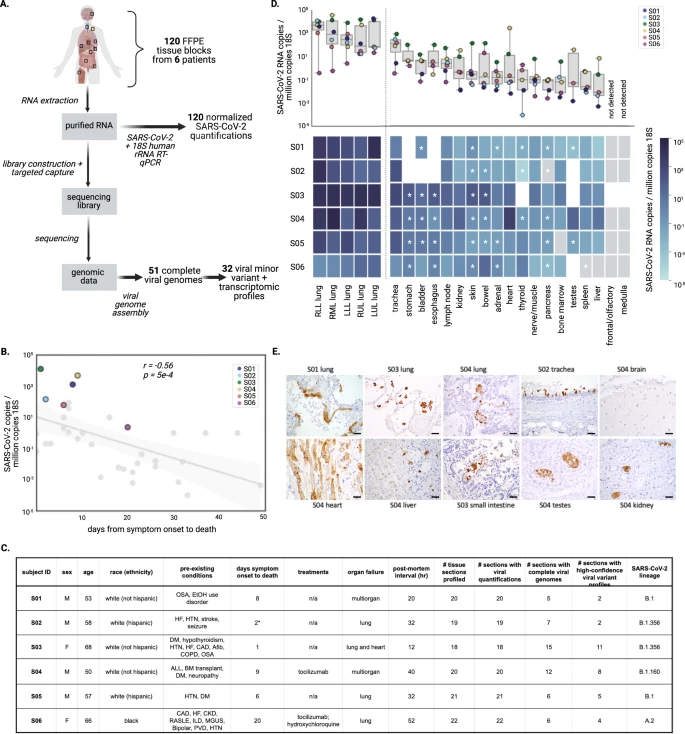
Quantification of SARS-CoV-2 viral load across tissues
Quantification of viral RNA by RT-qPCR enabled robust comparisons across the sample set, revealing substantial variation in viral load across different tissues.
To minimize systematic variation in comparisons across subjects, tissues, and experimental batches, we normalized viral loads by specimen cellularity (reported viral load constitutes SARS-CoV-2 RNA copies per million human 18S ribosomal RNA copies) (Supplementary Fig. 1A).
This normalization was particularly essential in this study given vast differences in cellularity across tissue types and specimens. Across the entire sample set, normalized viral loads ranged over eight orders of magnitude (1.1 × 10−4 to 3.2 × 104 viral copies per million 18S) (Fig. 1D). With the exception of S03, all subjects had at least one tissue where the viral load was not detected, or detected below the assay limit of detection (Fig. 1D and Supplementary Fig. 1b).
Lung specimens generally had high viral loads relative to other tissues, while varying substantially within and across subjects. The maximum viral loads detected for each subject always occurred in a lung specimen, but loads spanned three orders of magnitude across the six subjects (1.9 × 101 to 3.2 × 104 viral copies per million 18S).
Extrapulmonary viral load profiles across tissues were largely unique to each subject. Pancreas, liver, spleen, bone marrow, and peripheral nerve/skeletal muscle specimens were frequently associated with the lowest viral loads within subjects (Fig. 1D and Supplementary Fig. 1B). Specimens from the lymph node and GI tract had high normalized viral loads in several subjects. Other tissues, including the heart and kidney, were variable (Fig. 1D).
In general, extrapulmonary viral loads were more subject-specific, as they were highest in S03 (one reported day between symptom onset and death) and S04 (immunocompromised following a bone marrow transplant). We did not observe clear trends in extrapulmonary tissue viral loads over time between symptom onset and death (Supplementary Fig. 1D).
We identified SARS-CoV-2 nucleocapsid protein in 3/6 kidney, 2/6 liver, 2/6 lymph node, 2/6 GI tract, 1/6 heart, 0/6 spleen, 0/5 brain, and 1/3 testis specimens using this method (Supplementary Data File 2). We observed SARS-CoV-2 protein in the cardiomyocytes in the heart, hepatocytes in the liver, small intestine epithelial cells, rete testis, and tubular epithelium in kidneys (Fig. 1E).
To demonstrate that this signal was due to specific staining, we also stained the heart specimen where we had detected viral antigen with the nucleocapsid IHC assay (from S04), and one heart specimen where we did not (from S06), for SARS-CoV-2 spike protein, and confirmed that the results from this assay were consistent (Supplementary Fig. 1C).



{kind=link}