











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Reaching for and grasping objects brings about changes in the connections of a certain neuronal population in the red nucleus, a region of the midbrain, now reported by researchers at the University of Basel’s Biozentrum.
They have also shown how fine motor tasks promote plastic reorganization of this brain region.
The results of the study have been published in Nature Communications.
Simply grasping a coffee cup requires fine motor coordination with the highest precision.
This required performance of the brain is an ability that can also be learned and trained.
Prof. Kelly Tan’s research group at the Biozentrum, University of Basel, has investigated the red nucleus, a region of the midbrain that controls fine motor movement, and identified a new population of nerve cells that changes when fine motor coordination is trained.
The more that grasping is practiced, the more the connections between the neurons of this group of nerve cells are strengthened.
Grasping is a skill that can be trained and improved, even in adults.
For muscles to perform a movement correctly, brain commands must be transmitted through the spinal cord.
The red nucleus, which, over the years, has received little attention in brain research, plays an important role in fine motor coordination. Here, the brain learns new fine motor skills for grasping and stores what it has learned.
Kelly Tan’s team has now investigated the red nucleus in more detail in the mouse model and analyzed its structure and neuronal composition.
“We have found that this brain region is very heterogeneous and consists of different neuron populations,” says Giorgio Rizzi, first author of the study.
Improved fine motor skills through plastic changes in the brain
The research team has characterized one of these neuron populations and demonstrated that learning new grasping movements strengthens the connections between the individual neurons.
“When learning new fine motor skills, the coordination of this specific movement is optimized and stored in the brain as a code,” explains Tan.
“Thus, we have been able to also demonstrate neuroplasticity in the red nucleus.”
In a further step, the team now wants to investigate the stability of these strengthened nerve cell connections in the red nucleus and find out to what extent they regress when the learned fine motor movements are not practiced.
The findings could also provide new insights into the understanding of Parkinson’s disease, in which affected individuals suffer from motor disorders.
The team hopes to find out whether the neuronal connections in the red nucleus have also changed in these patients and to what extent fine motor training can restrengthen the neuronal network.
Anatomy
External features of the midbrain
The midbrain is the short, constricted portion, which connects the pons and cerebellum with the thalamus and cerebral hemispheres. It is the smallest part of the brainstem, not more than 2 cm in length, and most of it lies in the posterior cranial fossa [3, 5].
On the anterior surface of the midbrain are located the cerebral peduncles, separated by the interpeduncular fossa. The cerebral peduncle is two large bundles of fibers on each side of the midline. In addition, it is again divided into an anterior part, the crus cerebri, and a posterior part, tegmentum mesencephali, by a pigmented band of gray matter called substantia nigra.
The crura cerebri are superficially corrugated and emerge from the cerebral hemispheres.
They converge as they descend and meet as they enter the pons, where they form the caudolateral boundaries of the interpeduncular fossa. Two crura are separate, whereas the tegmental parts are united and traversed by the cerebral aqueduct that connects the third and fourth ventricles [1, 2].
The oculomotor nerve (CN III) emerges from the medial aspect of the cerebral peduncle of the same side. Exiting from the interpeduncular fossa near the junction of the pons and midbrain are the oculomotor nerves.
This cranial nerve supplies all but two of the extraocular muscles. The crus cerebri embraces a midline depression called the interpeduncular fossa. The basilar artery divides in the interpeduncular fossa into right and left posterior cerebral arteries at the level of the tentorial incisura.
The superior cerebellar and posterior cerebral arteries run laterally around the ventral (basilar) crural surfaces. The trochlear and oculomotor nerves lie between two arteries. In the depths of the interpeduncular fossa can be seen numerous small holes. These holes represent the entry point of the posterior cerebral artery. Because of its appearance, this region is usually referred to as the posterior perforated substance [6–8].
On the posterior surface of the midbrain are four prominent rounded elevations, the inferior and superior colliculi one on each side. Collectively, four colliculi are called as tectum (corpora quadrigemina). The superior and inferior colliculi are separated by a cruciform sulcus.
The upper limit of the sulcus expands into a depression for the pineal gland. Median frenulum veli is prolonged from its caudal end down over the superior medullary velum.
The superior colliculi are larger and darker than the inferior colliculi, and associated with visual responses.
The inferior colliculi are smaller, and associated with auditory pathways. Each colliculus is laterally related to ridges called superior brachium and inferior brachium, coming from respective colliculi.
Superior brachium connects the superior colliculus to lateral geniculate body. Inferior brachium connects the inferior colliculus to medial geniculate body.
The trochlear nerves (CN IV) arise from the dorsal midbrain, caudal to the inferior colliculi and pass inferiorly around the lateral side of the midbrain.
The trochlear nerve is the only cranial nerve that exits from the dorsal surface of the brainstem.
The midbrain serves important functions in motor movement, particularly movements of the eye, and in auditory and visual processing [1, 2, 7].
Internal structure of the midbrain
On transverse section, the cerebral peduncles are seen to be composed of dorsal and ventral regions separated by substantia nigra. On each side, the dorsal region is tegmentum, and the ventral part is the crus cerebri. Cerebral peduncles are the major pathways of motor neurons out of the cortex.
The tegmentum is between the substantia nigra and the aquaductus mesencephali.
It also refers to the corresponding regions in the medulla and pons. Tectummesencephali, located dorsal to the aquaductus mesencephali, contains two superior colliculi and two inferior colliculi (Figures 2 and 3) [1, 2, 6].

Crura cerebri (the cerebral peduncles, pedunculus cerebri)
The most ventral part of the midbrain contains a massive band of descending corticofugal fibers, the crus cerebri. Each crus cerebri is semilunar in section. It contains corticonuclear, corticospinal and corticopontine fibres.
Corticonuclear fibers (the corticobulbar fibers) originate in the primary motor cortex.
They descend through the genu of the internal capsule, and down to the midbrain.
In the midbrain, the middle third of the crus cerebri contains the corticobulbar and corticospinal fibers.
The corticobulbar fibers end in the motor nuclei of the cranial nerves and other brainstem nuclei. The corticospinal fibers are white matter motor pathways starting at the cortex, and they travel through the posterior limb of the internal capsule.
They enter the cerebral peduncle at the base of the midbrain, then pass through the brainstem, from the pons and then to the medulla. Corticospinal neurons synapse directly onto alpha motor neurons in the spinal cord for direct muscle control.
Corticopontine fibers arise in the cerebral cortex and form two groups, both of which end in pontine nuclei.
The frontopontine fibers arise from the cells of the frontal lobe, and end in the nuclei of the pons, the temporopontine fibers, which are largely from the posterior region of the temporal lobe, traverse the internal capsule, but occupy the lateral sixth of the ipsilateral crus [1, 5, 7].
Substantia nigra
The pigmented substantia nigra, the largest single nuclear mass in the midbrain, is connected massively with the basal ganglia, but it has other projections as well it is considered to subserve a motor function.
It looks like a darkened streak in unstained brain tissue; this is where it gets its name, which is Latin for “black substance.”
Although it is often referred to as one structure, the substantia nigra is actually made up of two anatomically and functionally distinct portions: the substantia nigra pars compacta and the substantia nigra pars reticulata [2, 3].
Neurons in the pars compacta are much more densely packed together (or compact) than those in the pars reticulata.
The compact zone appears as an irregular band of closely packed, large polygonal or pyramidal cells containing granules of melanin pigment.
The reticular zone, also known as the stratum intermedium, lies close to the crus cerebri, and is composed of irregular shaped scattered cells that are rich in iron, but they do not contain melanin pigment.
Most of the dopamine neurons of the brain originate in the midbrain and are found in either the substantia nigra or the ventral tegmental area, which is located adjacent to the substantia nigra [6, 8].
These dopamine neurons, however, are found predominantly in the substantia nigra pars compacta.
The pars reticulata is instead populated largely by GABA neurons.
Lesions of the substantia nigra or dopamine deficiencies result in Parkinson’s disease.
Although it is still not clear what exactly causes neurodegeneration in Parkinson’s disease, when a significant number of these neurons have died, the individual will likely start to experience movement‐related problems such as tremor, rigidity, slowing of movements, and postural instability—all hallmark symptoms of Parkinson’s disease [9].
Afferent fibers to the substantia nigra arise mainly from the caudate nucleus and the putamen. The efferent fibers of the substantia nigra project to the striatum and certain thalamic nuclei.
Mesencephalic tegmentum
Mesencephalic tegmentum is between the substantia nigra and the cerebral aqueduct. It usually contains ascending fiber tracts, cranial nerve nuclei, and the reticular formation nuclei.
The structures in this section are listed below:
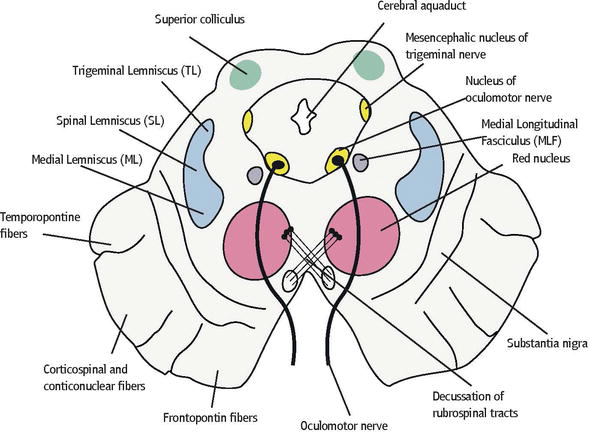
Red nucleus
The red nucleus is paired, oval shaped, and approximately 5 mm in diameter, midline structure that appears red in a freshly dissected specimen.
This unique appearance has been attributed to high vascularity of the structure in addition to the high level of iron pigments in the cytoplasms of its constituent neurons.
It blends rostrally with the nearby reticular formation and interstitial nucleus.
The nucleus consists of a large and ovoid column of cells extending from the caudal margin of the superior colliculus into the caudal diencephalon, and it appears as a circular mass which is traversed by the fibers of the oculomotor nerve [1, 3, 7].
Afferent fibers projecting to the red nucleus are derived from two principal sources, the cerebellar nuclei (approximately half of the fibers arising from the dentate nucleus pass rostrally beyond the red nucleus, cerebellorubral fibers), and the cerebral cortex (mainly from the precentral gyrus, corticorubral fibers).
Fibers of the superior cerebellar peduncle, arising from the dental, globose and emboliform nuclei, undergo a complete decussation in the caudal midbrain, and both enter and surround the contralateral red nucleus.
The efferent fibers of the red nucleus continue through the spinal cord, the brainstem and the cerebellum. Rubrospinal fibers issue from the medial margin of the red nucleus.
The fibers decussate and then run obliquely laterally in the ventral tegmental decussation (of Forel), ventral to the tectospinal decussation and dorsal to the medial lemniscus. Some efferent axons from a rubrobulbar tract to motor nuclei of the trigeminal, facial, oculomotor, trochlear and abducens nerves.
Uncrossed descending rubral efferents, from the parvocellular part of the nucleus, enter the central tegmental tract and project to the dorsal lamella of the principal inferior olivary nucleus. These fibers are referred to as rubro‐olivary fibers Physiology [3, 6, 7].
In humans, the lesions of a rubrospinal system are described as producing a syndrome characterized by contralateral motor disturbances that are variously designated as tremor, ataxia and choreiform activity and ibsilateral oculomotor palsy [1, 10].
Decussation of the superior cerebellar peduncle
All fibers of the superior cerebellar peduncle decussate at levels through the inferior colliculus. The decussation of superior cerebellar peduncle is the crossing of fibers of the superior cerebellar peduncle across the midline.
It comprises the cerebellothalamic tract, which arises from the dentate nucleus, as well as the cerebellorubral tract, which arises from the globose and emboliform nuclei and project to the contralateral red nucleus to eventually become the rubrospinal tract [2, 7, 8].
It is important as an anatomical landmark, as lesions above it cause contralateral cerebellar signs, while lesions below it cause ipsilateral cerebellar signs.
Trochlear nucleus
The nucleus of the trochlear nerve is small and nearly circular and is on a level with a plane carried transversely through the upper part of the inferior colliculus.
Root fibers emerging from the nucleus curve dorsolaterally and caudally in the outer margin of the central gray decusate completely in the superior medullary velum and from the dorsal surface of the brainstem caudal to the inferior colliculus [1, 2].
Root fibers emerging from the nucleus curve dorsolaterally and caudally in the outer margin of the central gray decussate completely in the superior medullary velum and exit from the dorsal surface of the brainstem caudal to the inferior colliculus.
And then, it curves around the lateral surface of the brainstem, passes between the superior cerebellar and posterior cerebral arteries, and enters the cavernous sinus. It innervates the superior oblique muscle that serves to: intort the eye when abducted, and depress the eye when adducted.
The trochlear nerve (CN IV) is the smallest nerve in terms of the number of axons it contains, and it has the longest length intracranially [1, 3, 5]. There are two major clinical syndromes that can manifest through damage to the trochlear nerve:
- Vertical diplopia: Injury to the trochlear nerve causes weakness of downward eye movement with consequent vertical diplopia.
- Torsional diplopia: Weakness of intorsion results in torsional diplopia, in which two different visual fields, tilted with respect to each other, are seen at the same time. To compensate, patients with trochlear nerve palsies tilt their heads to the opposite side in order to fuse the two images into a single visual field.
The clinical syndromes may originate from both peripheral and central lesions. A peripheral lesion is damage to the bundle of nerves, in contrast to a central lesion, which is damage to the trochlear nucleus [8, 10].
A lesion of the trochlear nucleus affects the contralateral eye. Lesions of all other cranial nuclei affect the ipsilateral side [10].
Parabigeminal nucleus
Ventrolateral to the inferior colliculus is a fairly well‐defined zone known as the parabigeminal area.
It is between the lateral lemniscus and the inferior colliculus and the surface of the brainstem that contains the parabigeminal nucleus. Some of the superior collicular efferents pass to the parabigeminal nucleus.
Studies have shown that the parabigeminal nucleus is active with fixed or moving objects. In addition, the parabigeminal nucleus plays a role in assessing the vision together with the colliculus superior [2, 3, 7].
Posterior commissure
The posterior commissure is located in the inferior pineal lamina and lies rostral to the superior colliculus at the place where the cerebral aqueduct becomes the third ventricle. It is one of the commissural fibers of the brain known to be important in the pupillary light reflex. Various nuclei are associated with the posterior commissure.
The best known of them is the interstitial nucleus of the posterior commissure, nucleus of Darkschewitsch; another one is the interstitial nucleus of Cajal. Fibers from the olivary nucleus cross on the opposite side and give collaterals to the visceral nuclei of the oculomotor complex (Edinger‐Westphal nucleus) [2, 3, 11].
Some fibers are believed to be derived from the posterior part of the thalamus and from the superior colliculus and to continue directly to the medial longitudinal fasciculus.
Fibers from the thalamic, pretectal, tectal region, and the habenular nuclei are known to connect with the posterior commissure, but they have not been shown anatomically.
Lesions in the nuclei of the posterior commissure, interrupting fibers from the intersititial nuclei of Cajal, produce bilateral eyelid retraction and impairment of vertical eye movements [12].
Pretectal area and pretectal nucleus
The pretectal region lies rostral to the superior colliculus at levels of the posterior commissure and rostrally by the habenular trigone, and laterally by the pulvinar thalami. This area is composed of several distinct cell groups, most of which are related to the visual system.
The nuclei of the pretectal area include pretectal olivar nucleus, medial, anterior and posterior pretectal nuclei, and optic tract nucleus [2, 13].
Afferents come from the lateral root of the optic tract from the retina, occipital cortical fields via the superior quadrigeminal brachium and the superior colliculus.
Efferents go to the ipsilateral and contralateral accessory oculomotor nucleus and superior colliculus.
They have bilateral efferent connections with the Edinger‐Westphal nucleus of the oculomotor nuclear complex by way of which they mediate the pupillary light reflex.
The effrent fibers reach both Edinger‐Westphal nucleui.
The decussate fibers pass ventral to the aqueduct or through the posterior commissure. In this way, sphincter pupillae contract in both eyes in response to impulses from either eye.
Therefore, only relatively large lesions involving multiple structures in the pretectum appear to impair the pupillary light reflex [2, 8, 13].
Medial longitudinal fasciculus
The medial longitudinal fasciculus is situated in the brainstem, and it is a set of crossed fibers with ascending and descending fibers.
The medial longitudinal fasciculus is a heavily myelinated composite tract lying near the midline, ventral to the periaqueductal gray matter.
It links three main nerves which control eye movements including saccades (rapid refixations), that is, the oculomotor, trochlear and the abducent nerves, as well as the vestibulocochlear nerve.
It interconnects the oculomotor, troclear, abducens, Edinger‐Westphal, vestibular, reticular and spinal accessory nuclei, coordinating conjugate eye movements and associated movements of the neck and head, including semicircular‐ and otolith‐mediated ocular motor reflexes [1, 12, 14].
The medial longitudinal fasciculus provides a neural mechanism for simultaneous contraction of the lateral rectus muscle on the one side, and the medial rectus muscle on the opposite side, required for conjugate lateral gaze. It also forms a major component of the optokinetic and vestibule‐ocular reflexes.
The most commonly recognized syndrome that results from the medial longitudinal fasciculus damage is internuclear ophthalmoparesis, which is characterized by slowing or limitation of adduction (on the same side as the medial longitudinal fasciculus lesion) during horizontal eye movements.
In patients with internuclear ophthalmoparesis, the contralateral abducting eye will usually exhibit a disassociated horizontal nystagmus, although this does not always occur [13, 15].
Oculomotor nucleus
The oculomotor nucleus is on the level of colliculus superior.
The upper end of the nucleus approaches the bottom of third ventricle in the vicinity of commissura caudalis and the lower end the top of colliculus inferior, lying adjacent to the inner ventral edge of stratum griseum centrale surrounding aquaeductus mesencephali.
The nucleus of the oculomotor nerve is about 10‐mm long.
The oculomotor nuclear complex containing subnuclei that give rise to the axons of the occulomotor nerve, both motor and parasympathetic fibers, is situated at the midline, at the level of the superior colliculus in the midbrain tegmentum [8].
The complex including the somatic portion is formed by multipolar motor neurons, and the parasympathetic portion is formed by oval or fusiform preganglionic cells, on each side of the median raphe.
The somatic portion consists of the lateral somatic cell column and the caudal central nucleus. It is divided into the principal, intrafascicular and extrafascicular parts.
The principal part is subdivided into the dorsal, intermediate and ventral portions. Isolated multipolar neurons are also found in the periaqueductal gray matter, the interstitial nucleus of Cajal, the Edinger‐Westphal nucleus, and the fiber bundles of the oculomotor nerve.
These cells most likely represent the displaced motor neurons of the oculomotor nerve. The Edinger‐Westphal nucleus consists of the rostral, ventral and dorsal parts [16]. The lateral somatic cell columns innervate the extraocular muscles.
The dorsal column innervates the inferior rectus muscle.
The intermediate cell column innervates the inferior ollique muscle, and the ventral cell column supplies fibers to the madial rectus muscle. A cell column medial to both dorsal and intermediate cell column is referred to as the medial cell column, and it innervates the superior rectus muscle.
The caudal central oculomotor nucleus is a midline somatic cell group found only in the caudal third of the complex, and it innervates the levator palpebrae muscle.
As a result, the oculomotor nuclear complex innervates all extraocular muscles except the lateral rectus and the superior oblique, supplies the levator palpebrae muscle, and provides preganglionic parasympathic fibers to the ciliary ganglion [17].
Visceral nuclei of the oculomotor nuclear complex consist of two distinct nuclear groups which are in continuity rostrally, and often are collectively referred to as the Edinger‐Westphal nucleus. Its afferent inputs come from the pretectal nuclei bilaterally and mediate the pupillary light reflex. In addition, they come from the visual cortex, mediating accommodation.
Efferent fibers relay through the ciliary ganglion and synapse upon postganglionic neurons, which give rise to the short ciliary nerves. The postganglionic fibers innervate the ciliary body, concerned with the mechanism of accommodation, and the sphincter of the iris [1, 17, 18].
Oculomotor nerve palsy results in weakness of the medial rectus, inferior rectus, superior rectus, inferior oblique, and levator palpebrae, leading to an eye that is “out and down “.
The oculomotor nerve also carries parasympathetic innervation to the pupil, responsible for pupil constriction. Oculomotor palsy may therefore leave the pupil dilated. Nuclear lesions usually occur due to small regions of infarction, and often there are no other neurological symptoms.
In ventral midbrain, lesions are due to HYPERLINK “https://radiopaedia.org/articles/benedikt-syndrome” Benedikt syndrome and HYPERLINK “https://radiopaedia.org/articles/weber-syndrome” Weber syndrome [2, 5, 19, 20].
Pupillary light reflex consists of a simultaneous and equal constriction of the pupils in response to illumination of one or the other eye.
The afferent axons of retinal ganglion cells pass into the optic nerve and decussate in the chiasm, and pass with the optic tract to the midbrain. The pupillary fibers do not synapse with the visual fibers in the lateral geniculate body, but pass to the pretectal nuclei at the level of the superior colliculus with intercalated fibers that pass as the efferent pupillary pathway to the Edinger‐Westphal nucleus of the oculomotor nerve on both sides.
Preganglionic parasympathetic fibers run in the oculomotor nerve as it leaves the brainstem.
The fibers pass downward to lie inferiorly in the inferior division of the third nerve as it enters the orbit. These fibers synapse in the ciliary ganglion and give rise to postganglionic parasympathetic myelinated short ciliary nerves [1,19, 20, 21].
The accommodation‐convergence reaction occurs when gaze is shifted from a distant object to a near one. The accommodation reflex has its afferent input from the primary visual pathway; sequentially retina, optic nerve, optic chiasm, optic tract, lateral geniculate body, optic radiation, visual cortex and area 17.
The peristriate area 19 interprets accommodation, and sends signals via the Edinger‐Westphal nucleus and the oculomotor nerve to the ciliary muscle, the medial rectus muscle and the sphincter pupillae muscle [20, 21].
Mesencephalic reticular formation
The mesencephalic reticular formation is less extensive than the pontine reticular formation caudal to it.
It is a neuronal structure located in the core of the brainstem, its caudal boundary crosses of the superior cerebellar peduncle, and extends rostrally to the thalamic reticular nucleus.
It is reciprocally interconnected with the superior colliculus. Even though detailed studies have indicated that the red nucleus is recognized as a distinctive part of the reticular formation, the principal reticular nuclei of the mesencephalon are:
1. The pedinculopontine nucleus,
2. The nucleus cuneiformis, and
3. The nucleus subcuneiformis.
Original function is defined as a part of the reticular activating system. There is clear evidence showing that there are subgroups of cells that participate in the control of saccadic and vergence eye movements.
The mesencephalic reticular formation has two major subdivisions. The posterior commissure in the sub‐human primate serves to separate the mesencephalic reticular formation into rostral and caudal regions.
The cells of the rostral portion of the mesencephalic reticular formation are associated with the control of vertical eye movements, while neurons in the caudal region also called as the central mesencephalic reticular formation are more closely associated with the control of horizontal eye movements [1, 7, 22].
Interpedincular nuclei
It is a ventral nucleus of the midbrain tegmentum lying between the right and left substantia nigrae, which cap the two cerebral peduncles.
The interpeduncular nucleus receives axons from the habenula, and it sends axons dorsally, to the midbrain raphe nuclei. The pathways described above constitute a part of the complex system by which impulses related to the limbic system are projected to midbrain levels [1–4].
The mesencephalic nucleus of the trigeminal nerve
The mesencephalic nucleus of the trigeminal nerve that is composed of large unipolar neurons forms a slight cell column near the lateral margin of the central gray of the upper part of the cerebral aqueduct.
The nucleus extends from the level of the motor nucleus into the rostral midbrain.
The cell bodies located in mesencephalic nucleus of the trigeminal nerve are actually connected to primary sensory fibers primarily coming from masticatory muscles.
This is the only place in the nervous system where the cell bodies of primary afferent fibers are found in the central nervous system rather than in ganglia outside of it. It houses proprioception for all muscles of the head and face. This nucleus has connections to the motor nucleus of the trigeminal nerve [1, 3, 13].
Afferent fibers of the mesencephalic nucleus of the trigeminal nerve convey proprioceptive impulses (kinesthesis and pressure) from the teeth, periodontium, hard palate, muscles of mastication and joint capsules. The mesencephalic nucleus of the trigeminal nerve receives afferent impulses from stretch receptors in the muscles of mastication.
The jaw proprioception pathway consists of sensory pseudounipolar neurons.
Their peripheral processes consist of stretch receptors that terminate in the muscles of mastication.
Their central processes bifurcate to send a branch to the principal sensory nucleus, and another branch to the rostral portions of the spinal nucleus.
Second‐order neurons from these nuclei project to the ventral posteromedial nucleus of the thalamus, which in turn projects to Brodmann’s area 3 a on the medial surface of the primary somatosensory cortex in the parietal lobe where position sense of the mandible enters conscious awareness.
Although proprioceptive information is generally processed by the mesencephalic neurons, their receptors, and their central connections, some proprioception is relayed from the temporomandibular joint, and extraocular muscle spindles [2–5, 13].
Sensory tracts of the tegmentum
Medial lemniscus: It is formed by the crossings of internal arcuate fibers, composed of axons of nucleus cuneatus and nucleus gracilis.
It ascends from the lemniscal decusstio on each side, as a flattened tract near the median raphe.
This large ascending fiber bundle can be readily followed through the brainstem to its termination in the ventral posterolateral nucleus of the thalamus.
On the transverse section of the midbrain, the medial lemniscus is dorsal in tegmentum.
Medial lemniscus is important for somatosensation from the joints and skin, and therefore, lesions of the medial lemnisci cause an impairment of vibratory and touch‐pressure sense [1, 2, 5].
Lateral lemniscus: The lateral lemniscus projects to the contralateral side from the dorsal cochlear nucleus.
The ventral cochlear nuclei project to the superior olivary nuclei on both sides as well as into the lateral lemniscus on each side.
Thus, the lateral lemniscus contains axons originating from cells in the dorsal and ventral cochlear nuclei and in the superior olivary nucleus.
The lateral lemniscus terminates in the central nucleus of the inferior colliculus. The principal ascending auditory pathway in the brainstem courses rostrally in the lateral part of the tegmentum [3, 6, 13].
Tectum
The tectum is located in the dorsal part of the midbrain, below the diencephalon. The name comes from the Latin word for “roof.”
Tectum is composed of a set of colliculi superior and colliculi inferior, which resemble small lumps and are responsible for initial processing of sensory information from the eyes and ears. Tectum is responsible for visual and auditory reflexes [3, 5, 13].
Superior colliculi
The superior colliculus refers to the rostral bump on the lateral side of the midbrain. It receives afferents from a number of sources including the retina, spinal cord, inferior colliculus and occipital and temporal cortices.
Collicular efferents pass to retina, lateral geniculate nucleus, pretectum, parabigeminal nucleus, thalamus and spinal cord.
They pass through the pulvinar relay to primary and secondary visual cortices
The tectobulbar and tectospinal tracts start from neurons in the superior colliculi and sweep ventrally round the central gray matter to decussate ventral to the oculomotor nuclei and medial longitudinal fasciculi as part of the dorsal tegmental decussations (of Meynert) (Figure 2). The superior colliculus is not restricted to a visual role alone.
It also helps orientation of the eyes and head. Part of the colliculus sticks out in the direction of the spinal cord region. This key projection helps the head to respond to different sensory stimuli.
The superior colliculus is concerned primarily with the detection of the direction of movement of object in the visual fields, and in this way, it facilitates visual orientation, searching and tracking [1, 2, 8].
The collicular stimulation produces contralateral head movement as well as movements involving the eyes, trunk and limbs, which implicates the superior colliculus in complex integration between vision and widespread body activity.
Inferior colliculi
The inferior colliculus is a part of the midbrain that serves as a main auditory (sound) center for the body (Figure 3). It consists of a compact nucleus of gray substance containing large and small multipolar nerve cells, and more or less completely surrounded by white fibers derived from the lateral lemniscus. Its primary roles are signal integration, frequency recognition, and pitch discrimination. It also processes sensory signals from the superior colliculi, located above it. The inferior colliculi is the relay station for Auditory Pathway. It receives fibers from the lateral lemniscus, the opposite inferior colliculus, the ipsilateral medial geniculate body and the auditory cortex. Most efferent fibers travel via the inferior brachium to the ipsilateral medial geniculate body. Some colliculogeniculate fibers do not relay in the geniculate body, but continue, with those that do, via the auditory radiation to the auditory cortex area. In few, they pass to the opposite inferior colliculus and the superior colliculus [2, 5, 8].
In experimental animals, lesions of either the inferior colliculus or its brachium produce defects in tonal discrimination, sound localization and auditory reflex. The effects of such lesions are poorly documented in humans (Table 1) [23].
More information: Giorgio Rizzi et al. Excitatory rubral cells encode the acquisition of novel complex motor tasks, Nature Communications(2019). DOI: 10.1038/s41467-019-10223-y
Journal information: Nature Communications
Provided by University of Basel



{kind=link}