










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Researchers know exercise improves mood for people with depression, but there are still questions about exactly how it makes people feel better.
The answers may be found in the endocannabinoid system.
The system is made up of the body’s own naturally occurring cannabinoid molecules and their receptors, said Jacob Meyer, an assistant professor of kinesiology at Iowa State University.
Activation of these receptors appears to strengthen connections in the brain and may influence pain and depression.
In a new study, Meyer and researchers at the University of Wisconsin-Madison, Medical College of Wisconsin and William S. Middleton Memorial Veterans Hospital, measured endocannabinoid levels and changes in mood before and after exercise sessions with varying levels of intensity.
They found a significant boost in endocannabinoids and improvements in mood following prescribed moderate-intensity exercise.
The findings are published in Medicine & Science in Sports & Exercise. Meyer says understanding the link between endocannabinoid levels, mood and exercise could lead to better treatment options.
“Finding alternatives to medication is important for the treatment of depression,” Meyer said.
“If we can figure out how exercise works with the endocannabinoid system, we could then design optimal exercise interventions.”
Preferred vs. prescribed exercise
The researchers analyzed data from 17 women with depression.
Each woman completed two, 30-minute exercise sessions on a stationary bike – one at a prescribed moderate intensity and a second of the participants’ own preferred intensity.
Both sessions improved mood, but the researchers only noted an increase in endocannabinoid levels for moderate intensity exercise.
There was no difference for preferred intensity.
Meyer says this was surprising, but the differences may stem from variation in the preferred session.
For example, some women stayed consistently at a lighter intensity, while others completed intervals. With a small sample, the variation may have obscured the relationship between the preferred intensity and change in endocannabinoids, Meyer said.
The findings are consistent with a 2016 study, in which Meyer and his colleagues looked at the effect of exercise on brain-derived neurotrophic factor or BDNF, a protein that regulates neuron growth and survival.
People with depression typically have lower levels of BDNF.
The 2016 study found that when women with depression were prescribed an exercise session similar in intensity to one they selected, their BDNF levels were significantly higher following the prescribed session.
“Having someone else prescribe the exercise could be involved in both the psychological and biological response to exercise,” Meyer said.
Effects appear to be long-lasting
For each exercise session, researchers collected blood samples immediately after exercise and also measured mood and anxiety at 10 minutes and 30 minutes post-workout.
Changes in endocannabinoids were significantly related to the improvement in mood up to 30 minutes after the moderate exercise, suggesting that the system contributes to the mood-enhancing effects of prescribed exercise, Meyer said.
“These results show us that the effect of prescribed moderate-intensity exercise on mood is generally pretty quick and lasts for a while at a relatively sustained rate,” Meyer said.
“However, given that people felt better after the preferred exercise session, even though it didn’t change endocannabinoid levels, multiple factors are likely involved in how exercise makes people feel better.”
That is the focus of Meyer’s ongoing Exercise and Brain Health Study. The goal is to develop new treatments for mental health conditions that include exercise.
Meyer is currently recruiting 20-to-60-year-old men and women to participate in the study. Click here to learn more.
What is the endocannabinoid system?
Each cell has many receptors, all of which are activated by special compounds specific to them – glutamate bind to glutamate receptors; serotonin to serotonin receptors; dopamine to dopamine receptors, endocannabinoids to cannabinoid receptors, and a whole lot more!
The endocannabinoid system is made up of the CB1 and CB2 receptors, which are activated by the two main endocannabinoids, anandamide and 2-AG.
Anandamide binds to the CB1 receptors, while 2-AG bind to both the CB1 and CB2 receptors.
The endocannabinoid system is our body’s system of endocannabinoid receptors and the endocannabinoids that interact with them.
An easy way to understand how the endocannabinoid system works is to think of it as a lock and key mechanism.
The cannabinoid receptors are the locks, and the endocannabinoids are the keys.
When the endocannabinoids bind to the cannabinoid receptors, they are able to produce physiological responses necessary for keeping cells alive and healthy.
In fact, you can say that the endocannabinoid system is one of the most important systems in the body because its main function is to maintain balance and homeostasis.
Where are the cannabinoid receptors located?
The cannabinoid receptors we currently know the most about are the CB1 and CB2 receptors.
In the future, researchers will likely find more cannabinoid receptors in the body.
Cannabinoid receptors are found all over the body.
They are everywhere – the brain, the spinal cord, the immune system, the internal organs, the peripheral nervous system, and even on the skin!
In fact, the endocannabinoid system is the most widespread receptor system[ii] in the human body.
However, the CB1 receptors are predominantly found in the central nervous system – your brain and spinal cord; the CB2 receptors are predominantly found in the immune system, specifically the white blood cells.
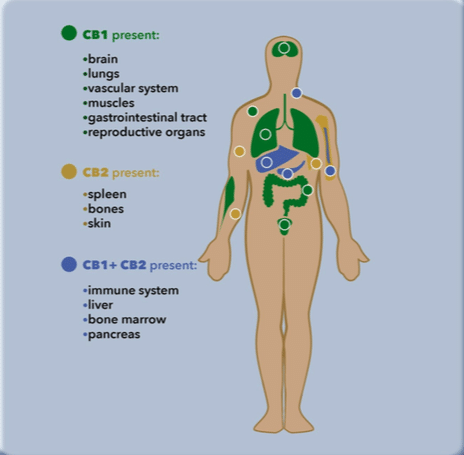
Stimulating the CB1 receptors produces a myriad of effects.
It regulates so many physiological processes – sleep, memory, emotional responses, mood, appetite, temperature, among others.
It can even regulate the sensation of pain.
Stimulating the CB2 receptors, on the other hand, produces widespread anti-inflammatory effects since they are mostly located in the immune cells.
There is the possibility that there is a third cannabinoid receptor, but its location and how it functions haven’t been clearly defined yet.
What are endocannabinoids?
There are two main types of endocannabinoids. They are anandamide and 2-AG.
Anandamide is the most studied and activates the CB1 receptors to produce their beneficial effects.
Unfortunately, anandamide is a short-order neurotransmitter.
The body only produces this endocannabinoid when it needs it.
Also, anandamide doesn’t stay long in the body because it’s easily degraded by an enzyme called fatty acid amide hydrolase or FAAH.
Whatever benefit we get from anandamide is short-lived because it is quickly broken down by these enzymes.
The same is true with the second endocannabinoid, 2-AG, which binds to both the CB1 and CB2 receptors.
It’s also produced on demand but is quickly degraded by monoacylglycerol lipase.
And just like anandamide, its actions are also short-lived.
Our endocannabinoids, although important and beneficial, can be limited by their short lifespan[iii].
What is the importance of the endocannabinoid system?
As mentioned earlier, the endocannabinoid system is a very integral system that modulates almost everything within the body.
The endocannabinoid system regulates[iv]:
- Gastrointestinal activity
- Cardiovascular activity
- Pain Perception
- Modulation of neurotransmitter release
- Maintenance of bone mass
- Protection of neurons
- Hormonal regulation
- Metabolism control
- Immune function
- Inflammatory reactions
- Inhibition of tumor cells
As you can see, your endocannabinoid system is responsible for a lot of important functions going on in your body!
The endocannabinoid system is the gatekeeper of homeostasis[v].
So how does the endocannabinoid system modulate almost everything? How does it work?
Before I tell you how the endocannabinoid system works, I’ll briefly explain to you how signals are normally transmitted by the cells.
This is important because this is one of the factors that make the endocannabinoid system unique.
Our cells communicate through chemical signals.
When a stimulus stimulates a cell, it will trigger the presynaptic portion of the cell to release chemicals, be it hormones, neurotransmitters, special proteins, signaling molecules, etc.
Now, the chemical signal will travel in a forward direction.
It will go from the presynaptic cell, cross the synapse, and bind to its specific receptor on the postsynaptic cell.
And on and on the signal will go until it reaches the brain.
The brain will then release chemicals that carry the command for how the body will react to the stimulus.
Now this is where the endocannabinoid system really gets interesting.
A stressor like a noxious stimulus will also trigger the endocannabinoid system to release endocannabinoids.
But instead of the presynaptic cell releasing the endocannabinoids, it is the postsynaptic cell that releases the chemical.
Endocannabinoids, unlike most neurotransmitters, travel in a backward direction called retrograde signaling[vi].
The postsynaptic neuron will release endocannabinoids, and the endocannabinoids will cross the synapse to attach to the presynaptic neuron’s cannabinoid receptors.
And what happens next is beautiful.
Endocannabinoids, when they bind to their receptors, have the ability to modulate the neuron’s activity.
This is how the endocannabinoid system maintains balance in the body.
Take for example a neurological disease.
The result of too many excitatory neurotransmitters in the brain will cause the receptors to become overexcited and overactive.
Because there is an imbalance, there will be a worsening of the disease. Case in point: stress and anxiety disorders.
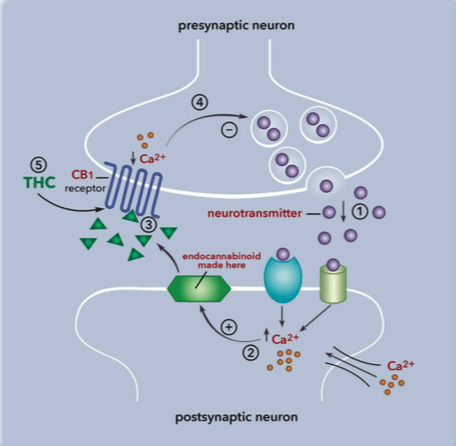
Now the endocannabinoids, when they’re released and attach to their receptors, can tell the neuron[vii] to stop producing or stop absorbing the excitatory neurotransmitters.
The neuron will listen and “calm down,” so homeostasis and balance are once more achieved in the body.
What this means is that even when a signal has already been fired by a neuron, endocannabinoids can still influence the neuron and change its response to the stimulus because of retrograde signaling.
The endocannabinoid system is a like a guard always watching over your body, ready to take action anytime there is a disturbance.
Here’s another very interesting thing about our endocannabinoid system.
Upon injury, immune cells will flock to the site of the injury and release a protein called cytokine.
The cytokine will then send the signal that it needs more immune cells in the area. On and on the cytokines will recruit more immune cells.
They only stop recruiting when anti-inflammatory proteins tell the immune cells to stop their cytokine production.
Now inflammation is good since it’s our natural and first-line-of-defense against irritants and pathogens.
But sometimes, the inflammatory process can become overactive and worsen the medical condition, which effectively delays the healing process.
In fact, there are numerous medical conditions worsened by inflammation – rheumatoid arthritis, multiple sclerosis, cancer, neuropathic pain, among others.
Now, endocannabinoids have the ability to inhibit inflammation[viii] by binding to the CB2 receptors of the cytokine-producing immune cells.
When these receptors are stimulated, they produce widespread anti-inflammatory effects by causing the apoptosis (cell death) of the immune cells.
With the immune cells inactivated, there will be no more production of cytokines.
Inflammation is controlled and proper healing can start.
The endocannabinoid system also regulates our blood pressure[ix]. Hypertensive?
The endocannabinoid system can lower your blood pressure. Hypotensive?
The endocannabinoid system can increase your blood pressure. Again, balance is promoted by the endocannabinoid system.
*-*-*-*-*-*-*
The classic “runners high” is described as the sense of well-being and mood elevation associated with moderate distance running.
Other typical indicators include a decrease in anxious thinking (anxiolytic), positive emotions/mood (euphoria), reduced pain perception (analgesia) and a feeling of increased endurance (Sparling et al., 2003; Dietrich and McDaniel, 2004; Tsatsoulis and Fountoulakis, 2006; Raichlen et al., 2012).
To explain these positive effects post-exercise, attention was directed to the endocannabinoid (eCB) system, and a number of groups have found significant correlations between physical activity, mood and elevated eCB levels.
Interestingly, the majority of studies have only observed significant rises in the first identified eCB, anandamide (AEA; Sparling et al., 2003; Heyman et al., 2012; Raichlen et al., 2013), whilst the reports analyzing 2-arachidonylglycerol (2-AG) levels post-exercise have been less clear.
Heyman et al. (2012) reported no change in circulating 2-AG levels after cycling. However, Brellenthin et al. (2017) showed that 2-AG and AEA were significantly increased in a study analyzing the effects of preferred (self-selected) and prescribed (70%–75% of max) exercise on eCB levels and mood.
The eCB system consists of the cannabinoid receptors 1 and 2 (CB1 and CB2), eCBs, and the enzymes that are responsible for their synthesis and breakdown (Devane et al., 1992; Mechoulam et al., 1995; De Petrocellis and Di Marzo, 2009). AEA and 2-AG are partial agonists of CB1 and CB2, whilst palmitoylethanolamine (PEA) and oleoylethanolamine (OEA) share similar synthesis and degradation mechanisms, without directly interacting with these receptors themselves (Hansen et al., 2000; Okamoto et al., 2004).
Instead, these molecules interact with other receptors, primarily peroxisome proliferator-activated receptor alpha (PPAR-α) and transient receptor potential cation channel subfamily V member 1 (TRPV1; Ahern, 2003; Fu et al., 2003; Lo Verme et al., 2005a,b; Karwad et al., 2017). eCB signaling mediates a number of physiological and psychological processes including emotional responses, cognition, memory, motor behavior, feeding and energy consumption (Berger and Motl, 2000; Cota et al., 2003; Cota, 2007; Brellenthin et al., 2017).
Studies have also established prominent roles of eCB signaling in the positive reinforcement in reward driven activities such as masturbation, arousal, binge-eating and social interactions in humans (Klein et al., 2012; Monteleone et al., 2015, 2017; Fuss et al., 2017).
Singing and dancing, especially as a group activity, are associated with positive mood in humans (Zajenkowski et al., 2015; Pearce et al., 2016; Tarr et al., 2016; Schladt et al., 2017).
However, little has been studied to elucidate how these positive emotions are mediated.
Recently, Hahn et al. (2017) studied the relationship between song practice and the eCB system in European starlings.
They found a significant positive correlation between conditioned place preference (a measure of reward and song production), the number of songs a bird produced and the expression of CB1 in areas of the brain associated with reward, primarily the ventral tegmental area.
Therefore suggesting a role for eCB signaling in singing and reward (Hahn et al., 2017; Riters et al., 2017).
In humans, singing has been studied as a therapy for long-term disorders such as Alzheimer’s (to improve cognition, memory and long-term pain), chronic obstructive pulmonary disease, as well as to improve mood in conditions such as anxiety and depression (Reagon et al., 2016; Kang et al., 2017).
Similarly, dancing has been explored as a potential therapy for cognitive and emotional dysfunction in conditions such as depression, dementia and Parkinson’s.
In a systematic review of 11 studies, Kiepe et al. (2012) found that depression and psychological distress were reduced by dance therapy in patients suffering from Parkinson’s, diabetes, breast cancer or heart failure.
Dance therapy in a group of 60 students also significantly reduced depression over a period of 12 weeks (Akandere and Demir, 2011).
To date, no study has assessed singing or dancing and whether they modulate eCB levels in humans and whether that correlates to an improved mood.
Given that mood is central in the measure of overall psychological well-being, low intensity activities that can positively modulate mood could be useful therapeutic tools in numerous conditions such as depression, anxiety and stress, especially if a patient cannot undertake moderate/higher intensity exercise.
The purpose of this study was to investigate whether activities other than running can give you a measurable “high” through changes in circulating eCBs levels.
We examined activities that are associated with euphoria (singing and dancing) as well as an exercise regime other than running (cycling), with the hypothesis that these activities would increase plasma eCB levels.
Quiet reading was used as a control condition. A secondary objective of this study was to establish whether there was a link between cycling, dancing, singing and reading with regards to mood and hunger ratings.

Endocannabinoid system (ECS) influences on mood and behavior. Complex relationships between internal and environmental factors converge on the ECS and modulate mood and behavior. The eCBs modulate different neurotransmitter systems including dopaminergic (DA) and serotoninergic (5-HT) projections onto the reward circuit. IFN-ɤ, interferon ɤ; IL-1β, interleukin-1β; TNF, tumor necrosis factor.
Inflammation and mood
Recent research has highlighted the relationship between the immune response and mood disturbances. In different inflammatory conditions, proinflammatory mediators could induce anxiety, anhedonia, social withdrawal, fatigue, and sleep disturbances, defined as “sickness behavior” (Raison et al., 2006; Dantzer et al., 2008; Miller et al., 2009).
Accordingly, systemic administration of proinflammatory agents is able to promote the activation of microglial cells in the hippocampus and stimulate the release of proinflammatory cytokines in the CNS (van Dam et al., 1992; Breder et al., 1994; Layé et al., 1994; Riazi et al., 2008), and is associated with depression and anxiety (Reichenberg et al., 2001; Miller et al., 2009).
Different inflammatory molecules, including interleukin (IL)-1β and tumor necrosis factor (TNF), have been implicated in the behavioral manifestations. In particular, central administration of these cytokines induces sickness behavior in animals (Dantzer et al., 2008; Haji et al., 2012; Rossi et al., 2012).
In addition, it has been proposed that inflammation may be implicated in the pathophysiology of different psychiatric syndromes, including major depression (Maes et al., 1995; Capuron et al., 2002).
Accordingly, elevated biomarkers of systemic inflammation, as C-reactive protein, have been associated with depressive symptoms (Morris et al., 2011).
It has been evidenced that depressed patients show higher peripheral blood levels of proinflammatory cytokines (Zorrilla et al., 2001; Dowlati et al., 2010; Haapakoski et al., 2015), and elevated inflammatory markers predict reduced response to antidepressant treatment (Strawbridge et al., 2015).
Accordingly, increased prevalence of depression has been observed in patients with autoimmune disorders (Zeher et al., 2010), and blocking cytokine signaling can exert beneficial effects on mood (Tyring et al., 2006).
Moreover, post-mortem studies in patients with major depression evidenced that also the innate immune response is altered, possibly contributing to the pathogenesis of depression (Martín-Hernández et al., 2018).
Finally, it has been suggested that also the efficacy of different psychoactive treatments may be mediated by immunomodulatory properties (Maes et al., 1999; Cattaneo et al., 2013; Horowitz et al., 2015).
The effects of neuroinflammation on mood have been extensively investigated in neurological diseases and particularly in multiple sclerosis (MS).
Anxiety and depression, are more frequent in MS patients compared to both general population (Patten et al., 2003) and other neurological patients (Schiffer and Babigian, 1984; Schubert and Foliart, 1993; Thielscher et al., 2013). In particular, mood disturbances may be independent of other neurological deficits and may occur as a presenting symptom (Haussleiter et al., 2009; Lo Fermo et al., 2010; Suh et al., 2010). It has been shown that in animal models of MS (i.e., experimental autoimmune encephalomyelitis, EAE) sickness behavior is observed during the acute phase before the onset of motor signs (Pollak et al., 2000) and comes from altered neuronal activity produced by the inflammatory milieu (Gentile et al., 2015a).
Experimental studies showed that in EAE specific proinflammatory cytokines, particularly TNF and IL-1β, are critically involved in the induction of mood alterations (Pollak et al., 2003; Gentile et al., 2015b).
Notably, the synaptic alterations have been found in the striatum, a key structure involved in mood regulation both in rodents and humans (Mathew and Ho, 2006; Nestler and Carlezon, 2006; Kim et al., 2008; Zhang et al., 2008).
In particular, striatal synaptic degeneration and dendritic spine loss have been found already during the early phases of EAE, independently of demyelination or clinical disability (Centonze et al., 2009).
This data suggest that TNF-induced striatal synaptic dysfunction may be responsible of the behavioral manifestations.
In line with these findings, intracerebroventricular injection of TNF in control mice was associated with anxious behavior (Haji et al., 2012).
Conversely, intracerebroventricular administration of etanercept, a TNF inhibitor, reduced anxious behavior and prevented the synaptic alteration in EAE mice (Haji et al., 2012). In addition, elevated levels of IL-1β and IL-1β mRNA have been evidenced in the striatum of EAE mice, together with reduced dopaminergic transmission (Gentile et al., 2015b). Accordingly, blocking IL-1β signaling in EAE mice can reduce mood alterations and restore DA release in the striatum (Gentile et al., 2015b).
Inflammation and ECS
A complex bidirectional interaction exists between the ECS and the immune system. eCBs signaling has immunosuppressant actions, in particular, CB2R stimulation reduce both inflammation (Pandey et al., 2009) and circulating proinflammatory mediators (Croxford and Miller, 2003).
Conversely, lipopolysaccharide administration modulates eCB signaling (Klein et al., 2003). In particular, microglial cells are recently emerging as key elements mediating the relationship between inflammation and the ECS. Microglia express CBRs, particularly CB2Rs, and is able to release both inflammatory mediators and eCBs (Stella, 2009; Boorman et al., 2016).
Notably, CB2R stimulation in microglial cells promote the release of anti-inflammatory cytokines (Ma et al., 2015).
Therefore, reduced CB2R stimulation may contribute to the pathogenesis of mood disorders. Accordingly, it has been evidenced that a polymorphism of the CB2R gene, producing reduced activation of the CB2R receptor, was associated with increased incidence of depression (Onaivi et al., 2008).
Moreover, it has been demonstrated that treatment with both minocycline and antidepressant drugs was associated to reduced release of proinflammatory molecules by microglial cells (Burke et al., 2014; Su et al., 2015).
Several studies evidenced that the ECS plays a critical role in mediating the effects of inflammation on synaptic functioning and mood. Accordingly, in EAE mice, CB1Rs stimulation may counteract the TNF-mediated synaptic alterations (Rossi et al., 2011). In particular, administration of a CB1R agonist, prevented the alterations of striatal transmission produced by TNF on brain slices.
Several evidences suggest that specific proinflammatory mediators, including IL-1β and IFNɤ, modulate striatal CB1R sensitivity (Rossi et al., 2012; Gentile et al., 2016; Mandolesi et al., 2017).
In particular, intracerebroventricular administration of IL-1β in control mice induced anxious behavior and was associated with reduced sensitivity of striatal GABAergic synapses to CB1Rs stimulation (Rossi et al., 2012).
It has been observed that IL-1β mediates the effects of inflammation in EAE mice (Gentile et al., 2016). Accordingly, blocking IL-1β signaling restored striatal CB1Rs sensitivity and reduced anxious behavior (Gentile et al., 2016).
Interestingly, it has been shown that inflammatory signals and environmental stimuli interact with the ECS to produce mood changes. In particular, the effects of IL-1β administration on striatal transmission were similar to the alterations induced by social defeat stress (Rossi et al., 2008).
Furthermore, administration of IL-1β receptor antagonist (IL-1ra) was able to revert the synaptic and behavioral effects induced by both IL-1β injection and social defeat stress. These results suggest that IL-1β is involved in inflammation-induced mood alterations and also play a key role in mediating responses to environmental stress (Koo and Duman, 2008; Goshen and Yirmiya, 2009; Norman et al., 2010).
More information: Jacob D. Meyer et al. Serum Endocannabinoid and Mood Changes after Exercise in Major Depressive Disorder, Medicine & Science in Sports & Exercise (2019). DOI: 10.1249/MSS.0000000000002006
Journal information: Medicine & Science in Sports & Exercise
Provided by Iowa State University



{kind=link}