











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



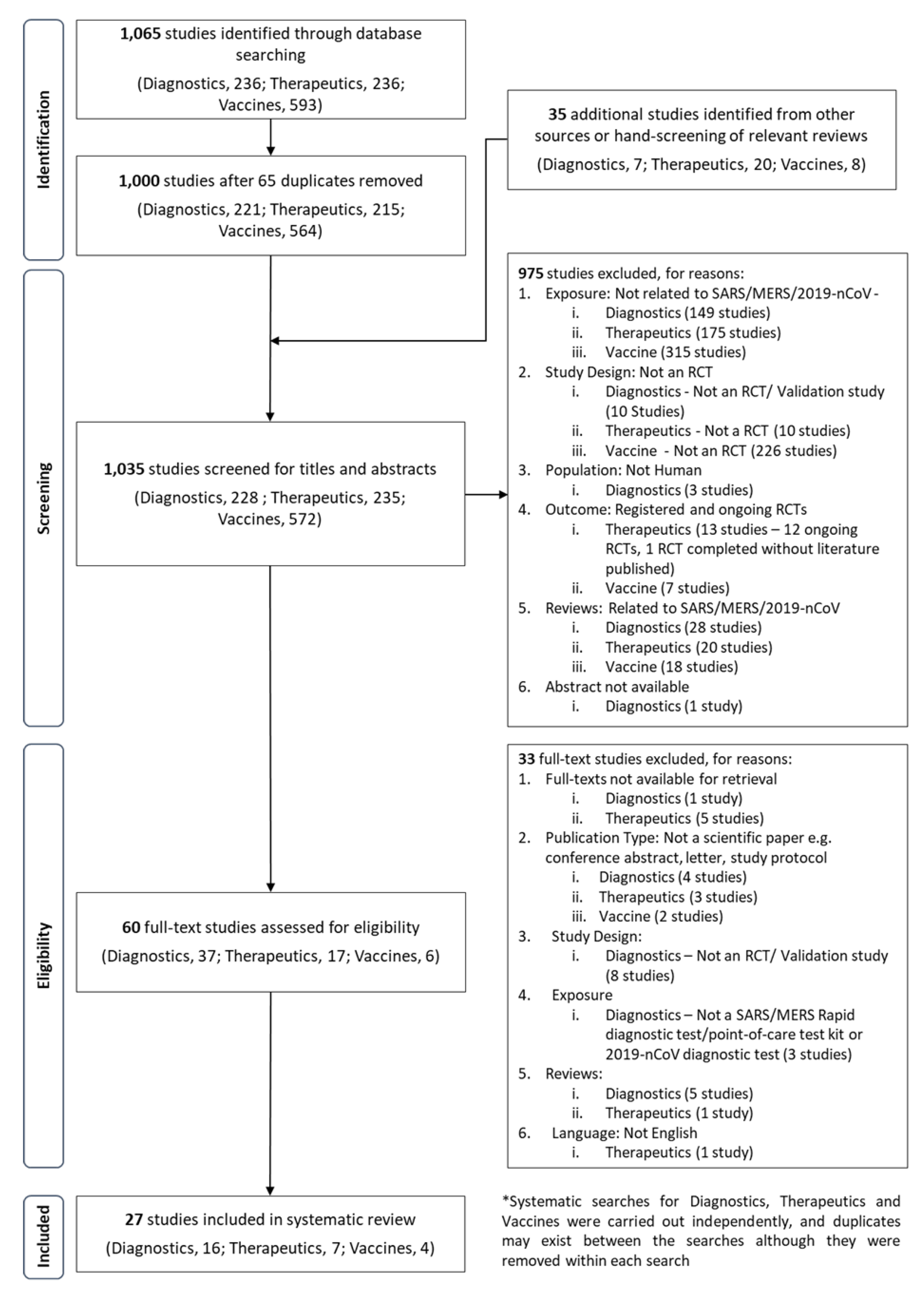
La diagnostica rapida, i vaccini e le terapie sono importanti interventi per la gestione dell’epidemia del 2019 nuovo coronavirus (2019-nCoV).
È opportuno rivedere sistematicamente il potenziale di questi interventi, compresi quelli per la sindrome respiratoria del Medio Oriente-Coronavirus (MERS-CoV) e la sindrome respiratoria acuta grave (SARS) -CoV, per guidare i responsabili politici a livello globale sulla definizione delle priorità delle risorse per la ricerca e lo sviluppo .
È stata effettuata una ricerca sistematica in tre principali database elettronici (PubMed, Embase e Cochrane Library) per identificare gli studi pubblicati in conformità con le linee guida per le segnalazioni sistematiche e le analisi sistematiche (PRISMA).
Sono state utilizzate strategie supplementari tramite Ricerca Google e comunicazioni personali. Un totale di 27 studi ha soddisfatto i criteri per la revisione.
Sono stati pubblicati numerosi protocolli di laboratorio per la conferma di casi sospetti 2019-nCoV che utilizzano la reazione a catena della polimerasi per trascrizione inversa in tempo reale (RT-PCR).
Un kit commerciale RT-PCR sviluppato dal Beijing Genomic Institute è attualmente ampiamente utilizzato in Cina e probabilmente in Asia.
Tuttavia, i test sierologici e i kit di test point-of-care non sono stati sviluppati ma sono probabili nel prossimo futuro. Diversi candidati al vaccino sono in cantiere. Il probabile primo vaccino di fase 1 sul vaccino è un candidato sintetico a base di DNA.
Numerosi nuovi composti e terapie concessi in licenza per altre condizioni sembrano avere efficacia in vitro rispetto al 2019-nCoV.
Alcuni sono stati testati in studi clinici contro MERS-CoV e SARS-CoV, mentre altri sono stati elencati per studi clinici contro 2019-nCoV. Tuttavia, attualmente non esistono efficaci antivirali specifici o combinazioni di farmaci supportati da prove di alto livello.
In risposta a questa crisi di salute pubblica, i ricercatori del Precision Vaccines Program (PVP) del Boston Children’s Hospital sono in prima linea nello sviluppo di un vaccino appositamente mirato per le popolazioni più anziane – coloro che sono maggiormente a rischio di sviluppare la malattia del coronavirus nel 2019 (COVID- 19), causato da grave coronavirus della sindrome respiratoria acuta-2 (SARS-CoV-2).
Secondo l’Organizzazione mondiale della sanità, gli anziani e le persone con condizioni mediche preesistenti sembrano essere più vulnerabili a diventare gravemente malati con COVID-19.
“Le persone anziane hanno un sistema immunitario diverso rispetto agli adulti di mezza età sani e spesso non rispondono altrettanto efficacemente all’immunizzazione, quindi un vaccino di una taglia non va bene per tutti”, ha affermato Ofer Levy, professore di pediatria presso la Harvard Medical School e direttore di il PVP.
Concentrandosi su adiuvanti
L’attuale antigene utilizzato per lo sviluppo del vaccino è la proteina del picco del coronavirus, così chiamata perché si trova in cima al picco di una particella del coronavirus. Questa è la parte del virus che “ricorda” il sistema immunitario.
La strategia di PVP è quella di combinare la proteina spike di coronavirus con adiuvanti: piccole molecole aggiunte a un vaccino per aumentare la risposta immunitaria del ricevente.
“Complessivamente, speriamo che un approccio adiuvante di precisione aiuti i vari sforzi di vaccinazione in corso in tutto il mondo”, ha affermato Levy.
“Gli adiuvanti possono essere cruciali per ottenere una risposta immunitaria più forte, più duratura e più ampia, specialmente tra quelli con un’immunità indebolita, come gli anziani.”
L’approccio del team è insolito nella gamma e nella novità degli adiuvanti che vaglieranno, ha affermato Levy.
Oltre a una raccolta di adiuvanti noti del vaccino, il team testerà i nuovi adiuvanti che ha scoperto nel programma di scoperta adiuvante finanziato dal National Institutes of Health.
“Questi adiuvanti sono stati scoperti mediante screening contro le cellule umane”, ha detto Levy. “Questo approccio specifico per specie rappresenta un esempio di vaccinologia di precisione”.
Età specifica
Il PVP prevede di testare una varietà di adiuvanti e combinazioni di adiuvanti nei globuli bianchi umani provenienti da persone anziane. I ricercatori studieranno quindi le risposte immunitarie indotte da adiuvanti.
“Il nostro schermo, confrontando gli adiuvanti individuali e di combinazione con e senza l’antigene del coronavirus, identificherà una combinazione di adiuvanti che induce in modo più efficace una risposta immunitaria ottimale negli anziani”, ha detto Levy.
Queste schermate inizieranno immediatamente e continueranno per le successive 6-8 settimane.
“Speriamo di avere un segnale chiaro nei prossimi mesi con quale adiuvare il vaccino per proseguire con i test clinici”, ha aggiunto.
Tipicamente, la ricerca sui vaccini a base di adiuvante non considera le specie o l’età nelle fasi di scoperta e sviluppo iniziale.
“Il nostro uso di sistemi in vitro umani specifici per età per ridurre il rischio e accelerare un vaccino adiuvato su misura per gli anziani è nuovo”, ha affermato Levy.
“In questo modo, stiamo portando la medicina di precisione alla vaccinologia.”
Gli adiuvanti potrebbero anche essere una strategia di riduzione dei costi, hanno detto gli autori, poiché l’antigene è in genere la parte più costosa di un vaccino.
“Se vogliamo essere in grado di fornire miliardi di dosi di vaccino, gli adiuvanti possono essere la risposta in quanto migliorano il sistema immunitario, quindi è necessario molto meno antigene per ottenere una risposta immunitaria protettiva”, ha detto Levy.
Test nei topi
Il gruppo sta inoltre studiando le risposte immunitarie del coronavirus in un modello animale vivente.
I primi topi sono già stati inoculati con una simile proteina spike di coronavirus derivata dal coronavirus SARS-2003 con o senza una combinazione adiuvante di piombo per ottenere una lettura precoce sulla misurazione di una risposta anticorpale.
Numerosi laboratori leader stanno fornendo input essenziali al PVP negli sforzi per sviluppare un vaccino coronavirus efficace e mirato che funzioni bene per le persone anziane:
- Barney Graham, vicedirettore del Centro di ricerca sui vaccini presso l’Istituto nazionale di allergie e malattie infettive (NIAID), ha fornito le proteine del picco di coronavirus al PVP poche settimane fa.
- Florian Krammer della Icahn School of Medicine di Mount Sinai ha fornito il virus SARS-CoV-2 dall’attuale ceppo.
- Peter Hotez e Maria Bottazzi della Baylor University hanno contribuito all’antigene dell’epidemia di SARS del 2003.
- Lindsey Baden, direttore della ricerca clinica presso il Brigham and Women’s Hospital, ha fornito campioni di sangue di pazienti anziani.
Immaginando un vaccino stagionale per coronavirus
Guardando al futuro, il PVP sta cercando di creare una piattaforma vaccinale e un programma sistematico che faciliterebbe lo sviluppo del vaccino in eventuali futuri focolai di coronavirus.
“Dr. Anthony Fauci, direttore del NIAID, ha avvertito del potenziale di SARS-CoV-2 di diventare un virus stagionale come l’influenza ”, ha affermato David Dowling, istruttore di pediatria presso l’HMS e membro del PVP.
“In tal caso, la comunità biomedica potrebbe aver bisogno di prendere in considerazione lo sviluppo di un vaccino stagionale annuale multivalente efficace contro più coronavirus.”

Secondo l’Organizzazione mondiale della sanità, gli anziani e le persone con condizioni mediche preesistenti sembrano essere più vulnerabili a diventare gravemente malati con COVID-19. L’immagine è adattata dal comunicato stampa di Harvard.
Poiché la sorveglianza internazionale in corso identifica i ceppi di coronavirus circolanti, il PVP prevede di collaborare con altri laboratori per ottenere le corrispondenti proteine virali e modellare le risposte degli anziani a diverse combinazioni adiuvanti / antigeni per determinare quale sia il più efficace.
Tre concetti di vaccino COVID-19
Levy e Dowling stimano che oltre 24 candidati al vaccino COVID-19 sono in via di sviluppo a livello globale. Questi vaccini si dividono in tre tipi generali:
- Vaccini a base di RNA. I primi vaccini COVID-19 in sviluppo hanno utilizzato l’RNA SARS-CoV-2. “Questo approccio è innovativo e attraente, ma anche se dimostrato efficace può essere difficile creare centinaia di milioni di dosi e ogni dose può essere relativamente costosa in quanto potrebbe richiedere una discreta quantità di RNA”, ha detto Dowling. “Sebbene ci sia meno esperienza con i test sull’uomo per questo approccio, è possibile che gli adiuvanti possano aiutare a migliorare la risposta immunitaria quando tali vaccini vengono somministrati a popolazioni più anziane e possono ridurre la quantità di RNA necessaria in ciascun vaccino.”
- Tecnologia basata sul DNA. Come i vaccini a base di RNA, questo approccio promettente utilizza il materiale genetico del virus per produrre un vaccino. “Puoi fare più dosi di vaccino con il DNA che con l’RNA, ma non è chiaro se la produzione possa essere rapidamente ridimensionata per soddisfare l’enorme domanda internazionale”, ha detto Dowling. “E, per quanto ne sappiamo, questi vaccini con acido nucleico non sono stati ancora testati su pazienti anziani.”
- Basandosi sui precedenti vaccini contro il coronavirus. Questo è l’approccio che il PVP sta adottando: apprendere dai precedenti vaccini dai precedenti focolai di coronavirus e renderli più efficaci. “Se aggiungi un adiuvante scelto appositamente per l’attività ottimale in una popolazione anziana, non solo funziona meglio in quel gruppo, ma può ridurre drasticamente il costo per dose di vaccino riducendo la quantità di antigene richiesta”, ha detto Dowling.
Risposte immunitarie in COVID-19 e potenziali vaccini: lezioni apprese dall’epidemia di SARS e MERS
Il mondo ha sperimentato le epidemie di infezione da coronavirus che minacciano la pandemia globale nel 2002-2003 dalla sindrome respiratoria acuta grave (SARS) e nel 2011 dalla sindrome respiratoria del Medio Oriente (MERS).
In entrambi i casi, gli agenti causali (SARS-CoV e MERS-CoV, rispettivamente) sono stati recentemente identificati coronavirus nel genere Betacoronavirus con origine zoonotica.
Alla fine del 2019, a Wuhan, Hubei, in Cina, è stato segnalato lo scoppio di un altro coronavirus che causa malattie respiratorie, una malattia ora ufficialmente chiamata “Malattia del Virus Corona 2019; COVID-19” .
Il coronavirus che è l’agente causale di questa malattia respiratoria è stato identificato e il suo genoma è completamente sequenziato
La sequenza genomica di SARS-CoV-2 ha mostrato una composizione genomica simile ma distinta di SARS-CoV e MERS-CoV.
Interazione ospite-patogeno: profili emergenti di infezione SARS-CoV-2
Le indagini sui modelli emergenti o sulle caratteristiche di trasmissione di SAR-CoV-2 sono aumentate dopo uno scoppio di casi confermati in tutto il mondo da dicembre 2019. Uno dei rapporti iniziali affermava che la maggior parte dei pazienti infetti confermati in laboratorio (27 su 41 casi) aveva collegamenti con il mercato ittico di Wuhan
È stata tentata l’identificazione della fonte o dell’ospite intermedio di SAR-CoV-2, concentrandosi su animali normalmente scambiati sul mercato, inclusi serpenti, uccelli e altri piccoli mammiferi.
Tuttavia, ad oggi, nessuna associazione animale specifica con SAR-CoV-2 è stata identificata in modo definitivo. Si ritiene che il candidato ospite intermedio più probabile sia pangolino poiché le sequenze genetiche del coronavirus degli animali e degli umani infetti durante l’epidemia hanno mostrato una corrispondenza del 99%, che è stata riportata dai ricercatori in una conferenza stampa il 7 febbraio 2020.3
Parallelamente all’identificazione dell’ospite intermedio, uno studio condotto su una famiglia ha rivelato che 6 pazienti che erano stati in viaggio in Romania non avevano alcun contatto diretto con il mercato. Inoltre, uno dei membri della famiglia si è infettato anche senza un viaggio a Wuhan.
La conclusione è stata che SAR-CoV-2 poteva essere trasmesso da uomo a uomo, attraverso goccioline respiratorie o contatto ravvicinato.4
Uno studio più ampio condotto su 425 pazienti ha anche confermato la trasmissione da uomo a uomo in cui la maggior parte dei pazienti (200 su 277) che sono stati diagnosticati nel periodo 1–22 gennaio 2020, non erano mai stati esposti al mercato di Wuhan o vicini contatto con soggetti con sintomi respiratori 5
Secondo la rapida diffusione di SAR-CoV-2, l’OMS ha emesso un allarme di emergenza pubblica di interesse internazionale (PHEIC) il 30 gennaio 2020. Al 20 febbraio 2020, sono stati segnalati in totale 75.725 casi confermati in almeno 29 paesi in tutto il mondo con un tasso di mortalità del 2,8% (2.126 su 75.282 casi) .6
Sebbene il tasso di mortalità sia di gran lunga inferiore a quello della SARS (9,14%) 7 e MERS (34,4%), 8 i casi confermati accumulati, entro circa 2 mesi dall’interruzione, hanno superato notevolmente la SARS (8.096 casi, dal momento che 2002) e MERS (2.494 casi, dal 2012), cfr. Figura 1 .
La natura altamente contagiosa di SAR-CoV-2 è probabilmente dovuta alla diffusione del virus attraverso un individuo infetto asintomatico che è stato riportato in Germania.9
Inoltre, i modelli matematici hanno stimato che la trasmissione che può verificarsi durante il periodo asintomatico di SARS-CoV e influenza è stata rispettivamente di circa il 5% e il 40 %.10
L’osservazione di 88 casi diagnosticati durante il 20-28 gennaio 2020 da individui con storia di viaggio a Wuhan, ha rilevato che il periodo medio di incubazione variava da 2,1 a 11,1 giorni (media = 6,4 giorni), 11 che è simile a un altro studio5 ed era nella stessa gamma con SARS-CoV e MERS-CoV.11
Sono stati inoltre segnalati periodi di incubazione più lunghi fino a 24 giorni12, ma ancora in discussione. Gli esperti dell’OMS hanno discusso durante la conferenza stampa del 10 febbraio 2020 che 24 giorni riferiti erano un’osservazione anomala o potrebbero essere dovuti alla doppia esposizione.13
È anche garantita l’identificazione del tropismo SARS-CoV-2. In accordo con la somiglianza del genoma con la SARS, è stata prevista l’analisi della sequenza di acido nucleico all’interno del dominio di legame del recettore della proteina spike (RBD) che SAR-CoV-2 potrebbe anche usare l’enzima 2 di conversione dell’angiotensina (ACE2) come recettore cellulare.14
Lo studio ha condotto esperimenti in vitro che potrebbero confermare che SAR-CoV-2 ha usato ACE2 per l’ingresso cellulare.15 Poiché la gamma selvaggia di specie animali (tranne ratto e topo) esprime ACE2, potrebbe supportare le specie incrociate osservate e da uomo a uomo eventi di trasmissione umana. I dati demografici e le caratteristiche emergenti di SARS-CoV, MERS-CoV e SAR-CoV-2 sono riassunti nella Tabella 1 .
Poiché SAR-CoV-2 è un nuovo patogeno che infetta l’uomo, studi recenti hanno anche tentato di definire una situazione di infezione più accurata e di prevedere l’epidemia nel prossimo futuro.
Utilizzando un modello matematico per calcolare il numero riproduttivo di base, R0 , che è il numero medio di persone a cui un individuo infetto trasmette il virus. Se R0 è superiore a 1, può verificarsi una trasmissione continua. R0 di SAR-CoV-2 variava da 2,2 a 2,6, con un tempo di raddoppio dell’epidemia di 6,4 giorni. 5,18
Ciò implica che, al fine di ridurre R0 al di sotto di 1, è necessario prevenire o controllare più della metà dell’infezione attuale.19 Confrontando con SARS-CoV e MERS-CoV che R0 erano <1 e 1.4-2.5, rispettivamente, implica che SAR-CoV-2 è più contagioso di MERS-CoV e può causare un’epidemia o persino una pandemia se la trasmissione non è controllata.


Immunopatologia di COVID-19
Il sito di infezione iniziale con SARS-CoV-2 è sconosciuto e la patogenesi di COVID-19 è ancora sotto inchiesta. Per la maggior parte dei pazienti, COVID-19 potrebbe interessare solo i polmoni perché è principalmente una malattia respiratoria.
La principale modalità di infezione è la trasmissione da uomo a uomo attraverso il contatto ravvicinato, che si verifica spruzzando goccioline da individui infetti attraverso la loro tosse o starnuti.
COVID-19 ha un probabile periodo di incubazione asintomatica tra 2 e 14 giorni durante i quali il virus può essere trasmesso.20
Per questo motivo, la rapida diffusione di SARS-CoV-2 si è verificata con l’ R0 di base di 2,2-2,6, il che significa che in media ogni individuo ha il potenziale per diffondere l’infezione ad altre 2,2 persone. 1,21
Sulla base dei dati dei pazienti ospedalizzati, la maggior parte dei casi di COVID-19 (circa l’80%) presentava sintomi asintomatici o lievi mentre i rimanenti sono gravi o critici.2,4 Sembra che la gravità e il tasso di mortalità di COVID-19 siano più lievi di quello di SARS e MERS.
Con presentazioni cliniche simili a quelle della SARS e della MERS, i sintomi più comuni di COVID-19 sono febbre, affaticamento e sintomi respiratori, inclusi tosse, mal di gola e mancanza di respiro.
Sebbene la diarrea sia stata presentata in circa il 20-25% dei pazienti con SARS e MERS, i sintomi intestinali sono stati raramente riportati in pazienti con COVID-19.1,2,4 La maggior parte dei pazienti ha anche sviluppato linfopenia e polmonite con caratteristici cambiamenti di opacità del vetro macinato polmonare sulla TC toracica. 1,2,4
Inoltre, lo studio di 41 pazienti ospedalizzati con alti livelli di citochine proinfiammatorie tra cui IL-2, IL-7, IL-10, G-CSF, IP-10, MCP-1, MIP-1A e TNFα erano osservato nei casi gravi COVID-19.2
Questi risultati sono in linea con la SARS e la MERS in quanto la presenza di linfopenia e “tempesta di citochine” può avere un ruolo importante nella patogenesi di COVID-19.22-24 Questa cosiddetta “tempesta di citochine” può iniziare sepsi virale e induzione infiammatoria danno polmonare che porta ad altre complicazioni tra cui polmonite, sindrome da distress respiratorio acuto (ARDS), insufficienza respiratoria, shock, insufficienza d’organo e potenzialmente morte. Ulteriori studi di autopsia o biopsia sono necessari per comprendere maggiori dettagli di questa malattia.
Allo stato attuale, il tasso di mortalità di COVID-19 nel mondo è circa del 2,4%, causato da insufficienza multiorgano soprattutto negli anziani e nelle persone con patologie di base come ipertensione, malattie cardiovascolari e diabete.
Risposte immunitarie innate all’infezione da SARS-CoV-2: acquisizione di informazioni dalle strategie utilizzate da SARS-CoV e MERS-CoV
Attualmente, sono disponibili solo informazioni limitate sullo stato immunitario innato dell’ospite dei pazienti con infezione da SARS-CoV-2. In un rapporto in cui sono stati studiati 99 casi a Wuhan, sono stati osservati aumento dei neutrofili totali (38%), riduzione dei linfociti totali (35%), aumento dell’IL-6 sierico (52%) e aumento delle proteine c-reattive (84%) 25
In un rapporto separato anche di Wuhan, è emerso che in 41 pazienti, un aumento dei neutrofili totali, una diminuzione dei linfociti totali nei pazienti in terapia intensiva vs terapia non-terapia intensiva sono risultati statisticamente diversi.
L’aumento dei neutrofili e la diminuzione dei linfociti sono anche correlati alla gravità e alla morte della malattia.1 Inoltre, i pazienti che necessitano di cure in terapia intensiva avevano livelli plasmatici più elevati di molte citochine innate, IP-10, MCP-1, MIP-1A e TNFα.2
Queste caratteristiche cliniche hanno suggerito la probabilità del coinvolgimento di condizioni altamente pro-infiammatorie nella progressione e nella gravità della malattia. Questo innalzamento precoce dei livelli sierici di citochine pro-infiammatorie è stato osservato anche nell’infezione SARS-CoV e MERS-CoV, suggerendo una potenziale gravità simile a una malattia con citochine temporanee simile. 23,24
Un’efficace risposta immunitaria innata contro l’infezione virale si basa fortemente sulle risposte di tipo I dell’interferone (IFN) e sulla sua cascata a valle che culmina nel controllo della replicazione virale e nell’induzione di un’efficace risposta immunitaria adattativa.
Mentre SARS-CoV e SARS-CoV-2 sembrano condividere il recettore di ingresso di ACE2, MERS-CoV utilizza dipeptidil peptididasi (DPP) -4 come recettore specifico.25
Il recettore putativo di SARS-CoV-2, ACE2, è principalmente espresso in un piccolo sottogruppo di cellule polmonari chiamate cellule alveolari di tipo 2.26 È stato riportato che SARS-Co-V infetta direttamente i macrofagi e le cellule T, una chiave caratteristica della patogenesi mediata da SARS-CoV.27
Non è noto se SARS-CoV-2 infetti le cellule immunitarie. Solo percentuali minime di monociti / macrofagi nel polmone hanno espresso ACE2.26
Se l’ACE2 viene minimamente espresso nelle potenziali cellule immunitarie bersaglio, è possibile che esistano altri recettori o che venga utilizzata un’altra modalità di ingresso cellulare come il potenziamento anticorpale ( Figura 2 ).

da SARS-CoV-2 L’assorbimento aerosol di SARS-CoV-2 porta all’infezione di ACE2 che esprime cellule bersaglio come cellule alveolari di tipo 2 o altre cellule bersaglio sconosciute. Il virus può smorzare le risposte IFN antivirali con conseguente replicazione virale incontrollata. L’afflusso di neutrofili e monociti / macrofagi provoca l’iperproduzione di citochine pro-infiammatorie. L’immunopatologia del polmone può essere il risultato delle “tempeste di citochine”. Th1 / Th17 specifici possono essere attivati e contribuiscono ad esacerbare le risposte infiammatorie. Le cellule B / plasmacellule producono anticorpi specifici per SARS-CoV-2 che possono aiutare a neutralizzare i virus. I punti interrogativi indicavano eventi ancora speculativi o sconosciuti. La figura è realizzata con biorender (https://biorender.com/).
Per innescare una risposta antivirale, le cellule immunitarie innate devono riconoscere l’invasione del virus, spesso attraverso schemi molecolari associati ai patogeni (PAMP). Per il virus RNA come il coronavirus, è noto che i PAMP sotto forma di RNA genomico virale o gli intermedi durante la replicazione virale incluso dsRNA, sono riconosciuti dai recettori dell’RNA endosomiale, TLR3 e TLR7 e dal sensore di RNA citosolico, RIG-I / MDA5.
Questo evento di riconoscimento porta all’attivazione della cascata di segnalazione a valle, ovvero NF-κB e IRF3, accompagnata dalla loro traslocazione nucleare. Nei nuclei, questi fattori di trascrizione inducono l’espressione dell’IFN di tipo I e di altre citochine pro-infiammatorie e queste risposte iniziali comprendono la difesa di prima linea contro l’infezione virale nel sito di entrata.28
L’IFN di tipo I tramite IFNAR, a sua volta, attiva il pathway JAK-STAT, in cui JAK1 e TYK2 chinasi fosforilati STAT1 e STAT2. STAT1 / 2 formano un complesso con IRF9 e insieme si spostano nel nucleo per iniziare la trascrizione dei geni stimolati dall’IFN (ISG) sotto il controllo dell’elemento di risposta stimolato dall’IFN (ISRE) contenente promotori.28 Un montaggio riuscito di questo tipo I La risposta dell’IFN dovrebbe essere in grado di sopprimere la replicazione e la diffusione virale in una fase precoce.
Per SARS-CoV e MERS-CoV, la risposta all’infezione virale per tipo I IFN è soppressa.
Entrambi i coronavirus utilizzano molteplici strategie per interferire con la segnalazione che porta alla produzione di IFN di tipo I e / o alla segnalazione a valle dell’IFNAR.
Questa strategia di smorzamento è strettamente associata alla gravità della malattia.29
Nella fase di induzione dell’IFN di tipo I, la SARS-CoV interferisce con la segnalazione a valle dei sensori di RNA direttamente o indirettamente come l’ubiquitinazione e il degrado delle molecole dell’adattatore del sensore di RNA MAVS e TRAF3 / 6 e inibendo la traslocazione nucleare dell’IRF3.30
MERS-CoV utilizza anche alcune di queste strategie con meccanismi aggiuntivi come la modifica repressiva dell’istone.30 Una volta che l’IFN di tipo I viene secreto, questi due virus sono dotati di un meccanismo che inibisce la segnalazione dell’IFN come la diminuzione della fosforilazione STAT1.28
Le proteine virali coinvolte nella modulazione di questa risposta IFN di tipo I ospite sono sia proteine strutturali (come M, N) sia proteine non strutturali (proteine ORF).
Sulla base del confronto della sequenza genomica, SARS-CoV condivide la somiglianza genomica complessiva con SARS-CoV o MERS-CoV, rispettivamente circa il 79% e il 50%. Il genoma di SARS-CoV-2 contiene anche regioni geniche aggiuntive (10b, 13, 14).
Inoltre, le sequenze di aminoacidi di alcune proteine putative di SARS-CoV-2 mostrano solo il 68% di somiglianza con quella di SARS-CoV .14
Pertanto, un attento confronto di sequenze di ciascuna regione genica può fornire una migliore previsione del modo in cui SARS-CoV-2 interferisce con la risposta immunitaria innata dell’ospite.
È parzialmente speculativo che SARS-CoV-2 utilizza strategie simili per modulare la risposta immunitaria innata dell’ospite, in particolare per smorzare la risposta IFN di tipo I, ma possono essere scoperti nuovi meccanismi aggiuntivi ( Figura 2 ).
Nei casi gravi o letali di infezione da SARS-CoV o MERS-CoV, si osserva costantemente un aumento dell’afflusso di neutrofili e monociti-macrofagi.27,31
In un modello murino di infezione da SARS-CoV, l’IFN disregolata di tipo I e i macrofagi infiammatori dei monociti sono la principale causa di polmonite letale.29
Pertanto, l’eccesso di IFN di tipo I con le cellule mielidi infiltrate è la principale causa di disfunzione polmonare e influisce negativamente sull’esito dell’infezione.
Si ipotizza che in caso di infezione da SARS-CoV o MERS-CoV, l’IFN ritardato di tipo I comprometta il controllo virale precoce, portando all’afflusso di neutrofili iperinfiammatori e monociti-macrofagi.
L’aumento di queste cellule immunitarie innate produce conseguenze deterioranti per l’ospite infetto che si è manifestato nell’immunopatologia polmonare, inclusa la polmonite o la sindrome da distress respiratorio acuto.
Nell’infezione SARS-CoV-2, ci si aspetta uno scenario simile con vari gradi di interferenza immunitaria. È interessante notare che la trasmissione del virus si verifica anche in individui infetti asintomatici. Ciò può essere indicativo di una risposta precoce ritardata della risposta immunitaria innata.
Sulla base dei dati accumulati per la precedente infezione da coronavirus, la risposta immunitaria innata svolge un ruolo cruciale nelle risposte protettive o distruttive e può aprire una finestra per l’intervento immunitario. La replicazione virale attiva in seguito porta all’iperproduzione di tipo I IFN e all’afflusso di neutrofili e macrofagi che sono le principali fonti di citochine pro-infiammatorie.
Con cambiamenti simili nei neutrofili e nei linfociti totali durante COVID19, SARS-CoV-2 induce probabilmente un IFN di tipo I ritardato e la perdita di controllo virale in una fase precoce dell’infezione. Gli individui sensibili a CoVID19 sono quelli con patologie di base, tra cui diabete, ipertensione e patologie cardiovascolari.2
Inoltre, non sono stati segnalati casi gravi nei bambini piccoli, quando la risposta immunitaria innata è altamente efficace. Questi fatti indicano fortemente che la risposta immunitaria innata è un fattore critico per l’esito della malattia.
Sulla base del presupposto che l’immunità innata svolge un ruolo chiave, possono essere proposti diversi interventi. IFN di tipo I, antagonisti di alcune citochine proinfiammatorie chiave e agenti antivirali sono alcuni di questi esempi.
Quando si utilizza l’IFN di tipo I per il trattamento, in un modello murino di infezione da SARS-CoV o MERS-CoV, i tempi di somministrazione sono fondamentali per ottenere una risposta protettiva.29
Risposte immunitarie adattive: un indizio per lo sviluppo futuro del vaccino?
In generale, la risposta immunitaria di tipo Th1 svolge un ruolo dominante nell’immunità adattativa alle infezioni virali. Il microambiente di citochine generato dalle cellule presentanti l’antigene determina la direzione delle risposte delle cellule T.
Le cellule T helper orchestrano la risposta adattativa complessiva, mentre le cellule T citotossiche sono essenziali per uccidere le cellule infette virali.
La risposta immunitaria umorale, in particolare la produzione di anticorpi neutralizzanti, svolge un ruolo protettivo limitando l’infezione in una fase successiva e previene la reinfezione in futuro. In SARS-CoV, entrambi gli epitopi delle cellule T e B sono stati ampiamente mappati per le proteine strutturali, le proteine S, N, M ed E.32
L’infezione da SARS-CoV induce sieroconversione già dal 4 ° giorno dopo l’insorgenza della malattia ed è stata riscontrata nella maggior parte dei pazienti entro 14 giorni. IgG specifiche di lunga durata e anticorpi neutralizzanti sono riportati fino a 2 anni dopo l’infezione.33
Infezione da F o MERS-CoV, si osserva sieroconversione alla seconda o terza settimana di insorgenza della malattia. Per entrambi i tipi di infezione da coronavirus, la risposta anticorpale ritardata e debole è associata a un esito grave.32
Sono stati riportati alcuni dettagli sierologici limitati di SARS-CoV-2. In uno studio preliminare, un paziente ha mostrato un picco specifico di IgM al giorno 9 dopo l’insorgenza della malattia e il passaggio a IgG entro la settimana 2.25 È interessante notare che i sieri di 5 pazienti con COVID-19 confermato mostrano una cross-reattività con SARS-CoV, ma non altri coronavirus . F
inoltre, tutti i sieri dei pazienti sono stati in grado di neutralizzare la SARS-CoV-2 in una placca in vitro , suggerendo un possibile montaggio delle risposte umorali con successo.25
Resta da verificare se il cinetico / titolo dell’anticorpo specifico sia correlato alla gravità della malattia.
La risposta delle cellule T in SARS-CoV è stata ampiamente studiata. In uno studio condotto su 128 campioni convalescenti, è stato riportato che le risposte delle cellule T CD8 + erano più frequenti con maggiore ampiezza rispetto alle risposte delle cellule T CD4 +.
Inoltre, le cellule T specifiche del virus del gruppo grave tendevano ad essere un fenotipo di memoria centrale con una frequenza significativamente più alta di cellule T polifunzionali CD4 + (IFNγ, TNFα e IL-2) e cellule T CD8 + (IFNγ, TNFα e stato degranulato) , rispetto al gruppo lieve-moderato.
Le forti risposte delle cellule T erano significativamente correlate con un anticorpo neutralizzante più elevato, mentre nel gruppo fatale sono state rilevate più citochine Th2 sieriche (IL-4, IL-5, IL-10 ).34 Per la mappatura degli epitopi, la maggior parte delle risposte (70%) sono state trovate contro le proteine strutturali (spike, enopeope, membrane e nucleocapsid).
Nell’infezione da MERS-CoV, l’aumento precoce delle cellule T CD8 + è correlato alla gravità della malattia e alla fase convalescente, si osservano cellule T helper di tipo Th1 dominanti.35 In un modello animale, le cellule T CD4 + di memoria delle vie aeree specifiche per l’epitopo conservato sono protettive contro la letalità sfida e può reagire in modo incrociato con SARS-CoV e MERS-CoV.36
Poiché i neutrofili svolgono un ruolo distruttivo in tutte le infezioni, il ruolo protettivo o distruttivo del Th17 nell’infezione da coronavirus umano rimane senza risposta.
Le evidenze attuali indicano fortemente che la risposta di tipo Th1 è la chiave per un controllo efficace di SARS-CoV e MERS-CoV e probabilmente vera anche per SARS-CoV-2.
La risposta delle cellule T CD8 +, sebbene cruciale, deve essere ben controllata per non causare patologie polmonari. Poiché la maggior parte degli epitopi identificati per entrambi i virus si concentrano sulle proteine strutturali virali, sarà informativo mappare quegli epitopi identificati con SARS-CoV / MERS-CoV con quelli di SARS-CoV-2.
Se si possono identificare epitopi sovrapposti tra i tre virus, sarà utile per l’applicazione nell’immunizzazione passiva usando siero convalescente da pazienti con SARS o MERS recuperati. Per gli epitopi delle cellule T, aiuterà in futuro la progettazione di vaccini cross-reattivi che proteggeranno da tutti e tre i coronavirus umani.
Potenziali meccanismi di evasione immunitaria
Le attuali osservazioni indicano che i coronavirus sono particolarmente adattati per eludere il rilevamento immunitario e smorzare le risposte immunitarie umane. Ciò spiega in parte il motivo per cui tendono ad avere un periodo di incubazione più lungo, in media 2-11 giorni rispetto all’influenza, 1-4 giorni.37
Il periodo di incubazione più lungo è probabilmente dovuto alle loro proprietà di evasione immunitaria, che sfuggono efficacemente alla rilevazione immunitaria dell’ospite nella fase iniziale dell’infezione. Come membro del genere Betacoronavirus, il meccanismo di evasione immunitaria è potenzialmente simile a quelli di SARS-CoV e MERS-CoV.
I meccanismi del modo in cui SARS-CoV e MERS-CoV modulano le risposte immunitarie dell’ospite sono stati ampiamente rivisti e discussi ( Figura 3 ) .30,38,39

I coronavirus interferiscono con più passaggi durante la risposta immunitaria innata iniziale, incluso il rilevamento dell’RNA (1 e 2), la via di segnalazione della produzione IFN di tipo I (3), l’attivazione STAT1 / 2 a valle dell’IFN / IFNAR (4) come indicato da segni di soppressione. Queste risposte IFN ritardate o attenuanti di tipo I incidono sull’attivazione immunitaria adattativa. La persistenza virale prolungata aggrava le risposte infiammatorie che possono portare all’esaurimento immunitario e alla soppressione immunitaria come meccanismo di regolazione del feedback.
La risposta distorta di tipo Th2 favorisce anche il cattivo esito della malattia.
In breve, la maggior parte dei meccanismi si basa sull’inibizione delle risposte immunitarie innate, in particolare il riconoscimento e la segnalazione dell’interferone di tipo I. Le proteine virali incluse le proteine di membrana (M) o non strutturali (NS) (ad es. NS4a, NS4b, NS15) sono le molecole chiave nella modulazione immunitaria dell’ospite.
In accordo con lo studio di cui sopra, l’analisi di due individui con infezione da MERS-CoV con gravità diversa ha rilevato che la risposta di interferone di tipo I nel paziente con esito negativo (decesso) era notevolmente inferiore rispetto al paziente guarito.40
Per l’evasione immunitaria adattativa, la presentazione dell’antigene tramite MHC classe I e MHC classe II è stata regolata verso il basso quando i macrofagi o le cellule dendritiche sono stati infettati con MERS-CoV, il che avrebbe notevolmente ridotto l’attivazione delle cellule T.38
Vaccini profilattici: è possibile?
A causa del rapido aumento delle infezioni SAR-CoV-2 e dei paesi colpiti, gli sforzi per sviluppare un vaccino SAR-CoV-2 efficace sono stati accesi in molti paesi.
Acquisendo le conoscenze dal percorso di sviluppo dei vaccini SARS e MERS, diversi gruppi di ricerca sono stati in grado di iniziare lo sviluppo del vaccino SAR-CoV-2 entro poche settimane dall’esplosione.
La selezione dell’antigene target e la piattaforma vaccinale sono probabilmente basati sugli studi sui vaccini SARS-CoV e MERS-CoV, riassunti nella Tabella 2 . Il picco a lunghezza intera (S) o S1 che contiene il dominio di legame del recettore (RDB) potrebbe essere considerato un buon antigene del vaccino perché potrebbe indurre anticorpi neutralizzanti che impediscono l’attaccamento e l’infezione delle cellule ospiti.
La Tabella 2 descrive gli antigeni e le piattaforme selezionati che sono stati testati per SARS-CoV e MERS-CoV in studi clinici e preclinici.
È interessante notare che, come riassunto nella Tabella 2 , il vaccino a base di acido nucleico, il vaccino a DNA, ha mostrato la piattaforma più avanzata in risposta ai patogeni emergenti.
Inoltre, durante l’epidemia del virus Zika, il vaccino a DNA è stato il primo candidato al vaccino ad entrare nella sperimentazione clinica (NCT02809443) 44 (meno di 1 anno dopo l’epidemia). Secondo l’attuale progresso tecnologico, il vaccino contro l’mRNA, un altro vaccino a base di acido nucleico, è stato considerato una tecnologia del vaccino dirompente.
I recenti progetti di vaccino contro l’mRNA hanno migliorato la stabilità e l’efficienza della traduzione proteica, quindi potrebbe indurre risposte immunitarie robuste.45,46 Sistema di consegna come nanoparticelle lipidiche, LNP è stato anche ottimizzato.47
Entro due mesi lo scoppio SAR-CoV-2, almeno 37 aziende biofarmaceutiche o settori accademici sono in gara per sviluppare il vaccino profilattico utilizzando diverse piattaforme tra cui mRNA, DNA, vettore adenovirale e proteina ricombinante .48

Al fine di rendere possibile il vaccino SAR-CoV-2, la raccolta di informazioni importanti per lo sviluppo e la valutazione del vaccino dovrebbe essere ben definita. Ciò include la ricerca di antigeni target, percorso di immunizzazione, protezione immunitaria correlata, modelli animali, scalabilità, struttura di produzione, profilo del prodotto target (TPP), previsione dell’epidemia e popolazione target.
La collaborazione internazionale e il trasferimento tecnologico tra esperti aiuteranno anche lo sviluppo del vaccino SARS-CoV-2 a progredire rapidamente.
Lezioni apprese da Zika, al fine di accelerare il vaccino disponibile durante l’epidemia in corso, potrebbe essere necessario eseguire studi preclinici sui candidati al vaccino SAR-CoV-2 parallelamente agli studi clinici.
Tuttavia, prima di accedere ai test clinici, le agenzie di regolamentazione devono valutare il processo di produzione e le informazioni precliniche per garantire la sicurezza dei volontari.49
Osservando le somiglianze e le differenze tra l’attuale SARS-CoV-2 e il precedente scoppio di SARS e MERS, emerge una sorprendente somiglianza con alcune caratteristiche uniche. Poiché il COVID-19 causa gravi preoccupazioni per la salute pubblica in tutta l’Asia e in un batter d’occhio influisce sulla popolazione mondiale,
un’indagine sulle caratteristiche di SARS-CoV-2, la sua interazione con le risposte immunitarie dell’ospite può aiutare a fornire un quadro più chiaro di come l’agente patogeno provoca malattie in alcuni individui mentre la maggior parte delle persone infette mostra solo sintomi lievi o assenti.
Inoltre, lo studio dei correlati immunitari di protezione e della memoria immunitaria a lungo termine da parte di individui convulsi può aiutare a progettare misure profilattiche e terapeutiche per lo scoppio futuro di coronavirus simili.
Conflitto d’interesse
Gli autori dichiarano assenza di conflitto di interesse
Riconoscimento
Gli autori desiderano ringraziare il professor Kiat Ruxrungtham per i suoi preziosi contributi scientifici e la lettura critica del manoscritto.
Riferimenti
- Wu F, Zhao S, Yu B, Chen YM, Wang W, Song ZG, et al. A new coronavirus associated with human respiratory disease in China. Nature [Preprint]. 2020 [cited 2020 Feb 16]: [19 p.]. Available from: https://doi.org/10.1038/ s41586-020-2008-3.
- Huang C, Wang Y, Li X, Ren L, Zhao J, Hu Y, et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet. 2020;395:497–506.
- Cyranoski D. Did pangolins spread the China coronavirus to people [Internet]. Heidelberg: Springer Nature; 2020 [cited 2020 Feb 16]. Available from: https://www.nature.com/articles/d41586-020-00364-2
- Chan JF, Yuan S, Kok KH, To KK, Chu H, Yang J, et al. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: a study of a family cluster. Lancet. 2020; 395: 514-23.
- Li Q, Guan X, Wu P, Wang X, Zhou L, Tong Y, et al. Early Transmission Dynamics in Wuhan, China, of Novel Coronavirus-Infected Pneumonia. N Engl J Med [Preprint]. 2020 [cited 2020 Feb 16]: [9 p.]. Available from: https://doi.org/10.1056/NEJMoa2001316
- Center for Systems Science and Engineering [Internet]. Baltimore: Johns Hopkins; c2020 [cited 2020 Feb 16]. Coronavirus COVID-19 Global Cases by Johns Hopkins CSSE 2020; [about 1 screen]. Available from: https://gisanddata.maps.arcgis.com/apps/opsdashboard/index.html#/b da7594740fd40299423467b48e9ecf6
- World Health Organization [Internet]. Geneva; World Health Organization; c2020 [cited 2020 Feb 16]. Summary of probable SARS cases with onset of illness from 1 November 2002 to 31 July 2003; [about 4 screens]. Available from: https://www.who.int/csr/sars/country/table2004 _04_21/en/.
- World Health Organization [Internet]. Geneva; World Health Organization; c2020 [cited 2020 Feb 16]. Middle East respiratory syndrome coronavirus (MERS-CoV) 2019; [about 4 screens]. Available from: https://www.who. int/emergencies/mers-cov/en/.
- Rothe C, Schunk M, Sothmann P, Bretzel G, Froeschl G, Wallrauch C, et al. Transmission of 2019-nCoV Infection from an Asymptomatic Contact in Germany. N Engl J Med [Preprint]. 2020 [cited 2020 Feb 16]: [2 p.]. Available from: https://www.nejm.org/doi/pdf/10.1056/NEJMc2001468? articleTools=true
- Fraser C, Riley S, Anderson RM, Ferguson NM. Factors that make an infectious disease outbreak controllable. Proc Natl Acad Sci U S A. 2004; 101(16):6146-51.
- Backer JA, Klinkenberg D, Wallinga J. Incubation period of 2019 novel coronavirus (2019-nCoV) infections among travellers from Wuhan, China, 20-28 January 2020. Euro Surveill. 2020;25(5).
- Guan W, Ni Z, Hu Y, Liang W, Ou C, He J, et al. Clinical characteristics of 2019 novel coronavirus infection in China. medRxiv [Preprint]. 2020 [cited 2020 Feb 10]: [30 p.]. Available from: http://medrxiv.org/content/ early/2020/02/09/2020.02.06.20020974.abstract
- World Health Organization holds news conference on coronavirus outbreak – 2/11/2020 [Internet]. New Jersey: CNBC Television; 2020 Feb 11 [cited 2020 Feb 16]. Video:1:12:45 hr. Available from: https://www. youtube.com/watch?v=a0Nu5MUR&feature=youtu.be&t=2166
- Lu R, Zhao X, Li J, Niu P, Yang B, Wu H, et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. Lancet. 2020; 395: 565-74.
- Hoffmann M K-WH, Krüger N, Müller M, Drosten C, Pöhlmann S. The novel coronavirus 2019 (2019-nCoV) uses the SARS-coronavirus receptor ACE2 and the cellular protease TMPRSS2 for entry into target cells. bioRxiv [Preprint]. 2020 [cited 2020 Feb 16]: [23 p.]. Available from: https://www. biorxiv.org/content/10.1101/2020.01.31.929042v1
- Wei M, Yuan J, Liu Y, Fu T, Yu X, Zhang ZJ. Novel Coronavirus Infection in Hospitalized Infants Under 1 Year of Age in China. JAMA [Preprint]. 2020 [cited 2020 Feb 16]:[2 p.]. Available from: https://jamanetwork.com/ journals/jama/fullarticle/2761659
- The Novel Coronavirus Pneumonia Emergency Response Epidemiology Team. Vital Surveillances: The Epidemiological Characteristics of an Outbreak of 2019 Novel Coronavirus Diseases (COVID-19) — China, 2020. China CDC Weekly. 2020; 2(8): 113-22.
- Wu JT, Leung K, Leung GM. Nowcasting and forecasting the potential domestic and international spread of the 2019-nCoV outbreak originating in Wuhan, China: a modelling study. Lancet [Preprint]. 2020 [cited 2020 Feb 15]: [9 p.]. Available from: https://doi.org/10.1016/S0 140-6736(20)30260-9
- Thompson R. Pandemic potential of 2019-nCoV. Lancet Infect Dis [Preprint]. 2020 [cited 2020 Feb 15]: [1 p.]. Available from: https://doi. org/10.1016/S1473-3099(20)30068-2
- Center for Disease Control and Prevention [Internet]. Atlanta: CDC; c2020 [cited 2020 Feb 10]. Symptoms of Novel Coronavirus (2019-nCoV); [about
1 screen]. Available from: https://www.cdc.gov/coronavirus/2019-ncov/ about/symptoms.html - Chen N, Zhou M, Dong X, Qu J, Gong F, Han Y, et al. Epidemiological and clinical characteristics of 99 cases of 2019 novel coronavirus pneumonia in Wuhan, China: a descriptive study. Lancet. 2020; 395:507-13.
- Nicholls JM, Poon LL, Lee KC, Ng WF, Lai ST, Leung CY, et al. Lung pathology of fatal severe acute respiratory syndrome. Lancet. 2003; 361(9371):1773-8.
- Mahallawi WH, Khabour OF, Zhang Q, Makhdoum HM, Suliman BA. MERS-CoV infection in humans is associated with a pro-inflammatory Th1 and Th17 cytokine profile. Cytokine. 2018;104:8-13.
- Wong CK, Lam CW, Wu AK, Ip WK, Lee NL, Chan IH, et al. Plasma inflammatory cytokines and chemokines in severe acute respiratory syndrome. Clin Exp Immunol. 2004;136(1):95-103.
- Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature [Preprint]. 2020 [cited 2020 Feb 15]: [15 p.]. Available from: https://doi. org/10.1038/s41586-020-2012-7
- Zhu N, Zhang D, Wang W, Li X, Yang B, Song J, et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N Engl J Med. 2020; 382(8):727-33.
- Perlman S, Dandekar AA. Immunopathogenesis of coronavirus infections: implications for SARS. Nat Rev Immunol. 2005;5(12):917-27.
- de Wit E, van Doremalen N, Falzarano D, Munster VJ. SARS and MERS: recent insights into emerging coronaviruses. Nat Rev Microbiol. 2016;14(8):523-34.
- Channappanavar R, Perlman S. Pathogenic human coronavirus infections: causes and consequences of cytokine storm and immunopathology. Semin Immunopathol. 2017;39(5):529-39.
- Kindler E, Thiel V, Weber F. Interaction of SARS and MERS Coronaviruses with the Antiviral Interferon Response. Adv Virus Res. 2016;96:219-43.
- Zumla A, Hui DS, Perlman S. Middle East respiratory syndrome. Lancet. 2015;386(9997):995-1007.
- Liu WJ, Zhao M, Liu K, Xu K, Wong G, Tan W, et al. T-cell immunity of SARS-CoV: Implications for vaccine development against MERS-CoV. Antiviral Res. 2017;137:82-92.
- Liu W, Fontanet A, Zhang PH, Zhan L, Xin ZT, Baril L, et al. Two-year prospective study of the humoral immune response of patients with severe acute respiratory syndrome. J Infect Dis. 2006;193(6):792-5.
- Li CK, Wu H, Yan H, Ma S, Wang L, Zhang M, et al. T cell responses to whole SARS coronavirus in humans. J Immunol. 2008;181(8):5490-500.
- Shin HS, Kim Y, Kim G, Lee JY, Jeong I, Joh JS, et al. Immune Responses to Middle East Respiratory Syndrome Coronavirus During the Acute and Convalescent Phases of Human Infection. Clin Infect Dis. 2019;68(6): 984-92.
- Zhao J, Zhao J, Mangalam AK, Channappanavar R, Fett C, Meyerholz DK, et al. Airway Memory CD4(+) T Cells Mediate Protective Immunity against Emerging Respiratory Coronaviruses. Immunity. 2016;44(6):1379-91.
- Lessler J, Reich NG, Brookmeyer R, Perl TM, Nelson KE, Cummings DA. Incubation periods of acute respiratory viral infections: a systematic review. Lancet Infect Dis. 2009;9(5):291-300.
- Shokri S, Mahmoudvand S, Taherkhani R, Farshadpour F. Modulation of the immune response by Middle East respiratory syndrome coronavirus. J Cell Physiol. 2019;234(3):2143-51.
- Kikkert M. Innate Immune Evasion by Human Respiratory RNA Viruses. J Innate Immun. 2020;12(1):4-20.
- Faure E, Poissy J, Goffard A, Fournier C, Kipnis E, Titecat M, et al. Distinct immune response in two MERS-CoV-infected patients: can we go from bench to bedside? PLoS One. 2014;9(2):e88716.
- Al-Amri SS, Abbas AT, Siddiq LA, Alghamdi A, Sanki MA, Al-Muhanna MK, et al. Immunogenicity of Candidate MERS-CoV DNA Vaccines Based on the Spike Protein. Sci Rep. 2017;7:44875.
- Du L, He Y, Zhou Y, Liu S, Zheng BJ, Jiang S. The spike protein of SARS-CoV–a target for vaccine and therapeutic development. Nat Rev Microbiol. 2009;7(3):226-36.
- Du L, Zhao G, He Y, Guo Y, Zheng BJ, Jiang S, et al. Receptor-binding domain of SARS-CoV spike protein induces long-term protective immunity in an animal model. Vaccine. 2007;25(15):2832-8.
- Tebas P, Roberts CC, Muthumani K, Reuschel EL, Kudchodkar SB, Zaidi FI, et al. Safety and Immunogenicity of an Anti-Zika Virus DNA Vaccine – Preliminary Report. N Engl J Med [Preprint]. 2017[cited 2020 Feb 10]:[16 p.]. Available from: https://doi.org/10.1056/ NEJMoa1708120
- Pardi N, Hogan MJ, Porter FW, Weissman D. mRNA vaccines – a new era in vaccinology. Nat Rev Drug Discov. 2018;17(4):261-79.
- Maruggi G, Zhang C, Li J, Ulmer JB, Yu D. mRNA as a Transformative Technology for Vaccine Development to Control Infectious Diseases. Mol Ther. 2019;27(4):757-72.
- Reichmuth AM, Oberli MA, Jaklenec A, Langer R, Blankschtein D. mRNA vaccine delivery using lipid nanoparticles. Ther Deliv. 2016;7(5):319-34.
- Koch S, Pong W, Editors. The count of companies developing vaccines for coronavirus rises 2020 [Internet]. Redwood: BioCentury; 2020 [cited 2020 Feb 16]. Available from: https://www.biocentury.com/article/304412
- Thomas SJ, L’Azou M, Barrett AD, Jackson NA. Fast-Track Zika Vaccine Development – Is It Possible? N Engl J Med. 2016;375(13):1212-6.
- Song Z, Xu Y, Bao L, Zhang L, Yu P, Qu Y, et al. From SARS to MERS, Thrusting Coronaviruses into the Spotlight. Viruses. 2019;11(1).
- Yong CY, Ong HK, Yeap SK, Ho KL, Tan WS. Recent Advances in the Vaccine Development Against Middle East Respiratory Syndrome
-Coronavirus. Front Microbiol. 2019;10:1781. - Schindewolf C, Menachery VD. Middle East Respiratory Syndrome Vaccine Candidates: Cautious Optimism. Viruses. 2019;11(1).
first part of Source: Harvard



{kind=link}