")










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A new QIMR Berghofer study has added weight to the potential benefits of using ultrasound treatment to deliver disease-targeting drugs to Alzheimer’s patients.
The study is the first to examine the technique on brain cells derived from human patients with Alzheimer’s disease, building on previous research on mice and other animal models.
The findings have been published today in the journal Stem Cell Reports.
Lead Australian researcher and head of QIMR Berghofer’s Cellular and Molecular Neurodegeneration group, Associate Professor Anthony White, said the researchers found using focused ultrasound coupled with microbubble treatment, could create openings in the blood-brain barrier formed by human endothelial cells.
“The blood-brain barrier is a semipermeable barrier that lines blood vessels in the brain and importantly protects brain tissue, but that protective function also prevents the uptake of drugs and therapies targeting brain diseases,” Associate Professor White said.
“We found by using focused ultrasound and microbubble treatment we could weaken the connection between blood-brain barrier cells, which would potentially allow the brain tissue to absorb a drug treatment.
“An abnormal blood-brain barrier has been associated with several neurodegenerative diseases, including Alzheimer’s disease, so it’s very important that we better understand how the barrier works and how we can safely penetrate it to combat disease.
“Our study is the first to look at how the blood-brain barrier cells from human patients can be disrupted to improve the uptake of Alzheimer’s therapies, building on previous studies that have explored if ultrasound could be used to reduce amyloid build up in the brains of mice and other animal models.”
The researchers used brain endothelial cells derived from human stem cells of people with a family history of Alzheimer’s disease to understand how those cells of the blood-brain barrier could be weakened.
They injected lipid microbubbles into the cells and then targeted the area with ultrasound which caused the cells to expand and contract, disrupting the connections between the cells, opening the blood-brain barrier.
The ultrasound and microbubble treatment technique was developed by researchers at the Queensland Brain Institute.
First author and QIMR Berghofer researcher, Dr. Lotta Oikari, said the study also showed the ultrasound and microbubble treatment had a longer lasting effect on the brain cells of Alzheimer’s patients than on healthy controls.

“The treatment generated openings in the monolayer of the blood-brain barrier of all patients, but the brain endothelial cells of healthy controls repaired themselves quicker than the Alzheimer’s patient cells,” Dr. Oikari said.
“The blood-brain barrier in Alzheimer’s patients was slower to repair, indicating they would be more receptive to drugs and treatments for longer and that brain ultrasound treatment may have to be adjusted differently depending on the type of disease the patient has.
“It also raises questions whether reduced integrity in the blood-brain barrier, following ultrasound treatment, could lead to lower amyloid levels in the brains of Alzheimer’s patients because the brain tissue could potentially expel the build-up.”
Funding: The research was funded by QIMR Berghofer.
The study involved researchers from QIMR Berghofer, the Queensland Brain Institute at The University of Queensland, Monash University, University of Helsinki and University of Eastern Finland, in Finland.
Adult neurogenesis appears to be restricted to two regions, i.e., the subventricular zone (SVZ) of the lateral ventricles and the subgranular zone (SGZ) of the hippocampal dentate gyrus (DG).
Importantly, adult hippocampal neurogenesis (AHN) was first reported over 50 years ago by Altman and Das [1], and newborn neurons are generated continuously throughout life in the mammalian brain, including the human brain [2, 3].
Since then, numerous studies have reported that AHN is implicated in cognition and endogenous repair mechanisms in normal physiological conditions such as learning and memory [4].
Interestingly, according to the recent research, the persistence of AHN appears to be decreased in aged adults and Alzheimer’s disease (AD) [5, 6].
AD is one of the major causes of age-related dementia and is characterized by cognitive impairment, amyloid-β deposition in plaques, tau hyperphosphorylation in neurofibrillary tangles, loss of synapses, loss of neuronal cells, and cholinergic dysfunction [7].
Dysfunction of the basal forebrain cholinergic (BFC) system, a significant characteristic of AD, induces neuropathological changes before clinical symptoms manifest [8,9,10]. The hippocampus and cortex receive gamma-aminobutyric acidergic, glutamatergic, and cholinergic input from the basal forebrain of the medial septum-diagonal band complex (MS/DB) [11, 12].
Thus, lesions in, or the inactivation of, cholinergic neurons in MS/DB result in a decrease of acetylcholinesterase (AChE) and choline acetyltransferase (ChAT), consequently diminishing AHN [13,14,15,16].
Despite intensive research efforts, none of the currently available treatments for AD can completely cure or prevent the course of age-related cognitive impairment, and the pathological mechanism is not clearly understood.
Numerous pharmacological therapies have been developed to treat AD [17]. However, 98% of small-molecule drugs (< 400 Da) and 100% of large-molecule drugs (> 500 Da) cannot cross the blood-brain barrier (BBB) [18], making the prevention and treatment of brain disorders difficult.
Focused ultrasound (FUS) combined with contrast agent microbubbles is a noninvasive technique that transiently opens BBB in targeted regions, thereby enabling localized therapeutic drug, gene, or nanoparticle delivery into the brain for treating central nervous system (CNS) disorders [19,20,21].
Considering that drugs that have been, or are currently being, developed for AD are mostly large molecules, FUS may enhance the effects of these drugs especially in patients with early-stage AD who have an intact BBB [22].
Moreover, several reports suggest that FUS stimulates neuronal activity and modulates proteomes and transcriptomes, independent of any therapeutic agent [23,24,25].
Previous studies indicate that FUS-mediated BBB opening can modulate the accumulation of amyloid-β and tau hyperphosphorylation in AD transgenic mice and increase AHN in wild-type mice [26,27,28,29,30]. Recently, Moreno-Jiménez et al. reported the persistence of AHN in human DG of subjects aged over 90 years; however, the number and maturation of immature neurons in DG sharply decreased in patients with AD.
This finding has gained attention for potential therapeutic strategies as an underlying memory impairment in AD [31]. However, it remains unclear whether FUS can modulate AHN in a cholinergic-deficient condition.
In this study, we investigated the effect of FUS on AHN and the cholinergic system in a cholinergic degeneration dementia rat model, which is a key pathogenic feature of dementia. Furthermore, if FUS was effective in increasing AHN, the synergistic effects of AHN modulation and drug delivery could improve treatment outcomes of AD.
Conclusions
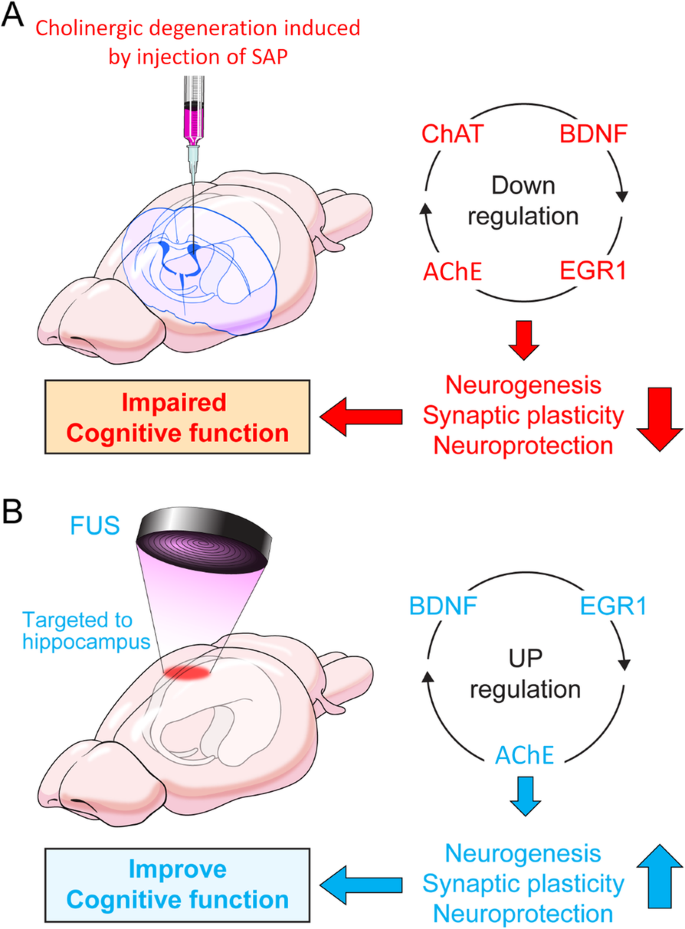
In the present study, we demonstrated that animals with BFC hypofunction causing spatial memory impairment exhibit a reduction in cholinergic activity, neurogenesis, and BDNF and ERG1 expression levels (Fig. 7a).
In contrast, FUS treatment increased AHN and improved spatial memory in cholinergic degeneration conditions. This improvement may be mediated by the upregulation of BDNF, EGR1, and AChE levels in the hippocampus, which is a critical factor for regulating AHN, synaptic plasticity, and neuroprotection (Fig. 7b).
Because patients with AD have impaired cholinergic neurons and AHN starting at the early stages, FUS treatment may restore AHN and have a protective effect against neurodegeneration.
Moreover, as FUS has been shown to be effective in increasing AHN, it could also contribute to increased permeability of BBB for drug delivery, and both these effects could be potential therapeutic strategies for AD.
References
- 1.Altman J, Das GD. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J Comp Neurol. 1965;124(3):319–35.
- 2.Eriksson PS, Perfilieva E, Bjork-Eriksson T, Alborn AM, Nordborg C, Peterson DA, et al. Neurogenesis in the adult human hippocampus. Nat Med. 1998;4(11):1313–7.
- 3.Ming G-L, Song H. Adult neurogenesis in the mammalian brain: significant answers and significant questions. Neuron. 2011;70(4):687–702.
- 4.Zhao C, Deng W, Gage FH. Mechanisms and functional implications of adult neurogenesis. Cell. 2008;132(4):645–60.
- 5.Tobin MK, Musaraca K, Disouky A, Shetti A, Bheri A, Honer WG, et al. Human hippocampal neurogenesis persists in aged adults and Alzheimer’s disease patients. Cell Stem Cell. 2019;24(6):974–82.e3.
- 6.Boldrini M, Fulmore CA, Tartt AN, Simeon LR, Pavlova I, Poposka V, et al. Human hippocampal neurogenesis persists throughout aging. Cell Stem Cell. 2018;22(4):589–99.e5.
- 7.Scheltens P, Blennow K, Breteler MM, de Strooper B, Frisoni GB, Salloway S, et al. Alzheimer’s disease. Lancet. 2016;388(10043):505–17.
- 8.Ginsberg SD, Che S, Wuu J, Counts SE, Mufson EJ. Down regulation of trk but not p75NTR gene expression in single cholinergic basal forebrain neurons mark the progression of Alzheimer’s disease. J Neurochem. 2006;97(2):475–87.
- 9.Grothe M, Heinsen H, Teipel SJ. Atrophy of the cholinergic basal forebrain over the adult age range and in early stages of Alzheimer’s disease. Biol Psychiatry. 2012;71(9):805–13.
- 10.Pappas BA, Bayley PJ, Bui BK, Hansen LA, Thal LJ. Choline acetyltransferase activity and cognitive domain scores of Alzheimer’s patients☆. Neurobiol Aging. 2000;21(1):11–7.
- 11.Manseau F, Danik M, Williams S. A functional glutamatergic neurone network in the medial septum and diagonal band area. J Physiol. 2005;566(Pt 3):865–84.
- 12.Baxter MG, Chiba AA. Cognitive functions of the basal forebrain. Curr Opin Neurobiol. 1999;9(2):178–83.
- 13.Perry EK, Tomlinson BE, Blessed G, Bergmann K, Gibson PH, Perry RH. Correlation of cholinergic abnormalities with senile plaques and mental test scores in senile dementia. Br Med J. 1978;2(6150):1457–9.
- 14.Bartus RT. On neurodegenerative diseases, models, and treatment strategies: lessons learned and lessons forgotten a generation following the cholinergic hypothesis. Exp Neurol. 2000;163(2):495–529.
- 15.Coyle JT, Price DL, Delong MR. Alzheimer’s disease: a disorder of cortical cholinergic innervation. Science. 1983;219(4589):1184–90.
- 16.Ricceri L, Minghetti L, Moles A, Popoli P, Confaloni A, De Simone R, et al. Cognitive and neurological deficits induced by early and prolonged basal forebrain cholinergic hypofunction in rats. Exp Neurol. 2004;189(1):162–72.
- 17.Cummings JL, Morstorf T, Zhong K. Alzheimer’s disease drug-development pipeline: few candidates, frequent failures. Alzheimers Res Ther. 2014;6(4):37.
- 18.Pardridge WM. Blood-brain barrier drug targeting: the future of brain drug development. Mol Interv. 2003;3(2):90.
- 19.Park J, Aryal M, Vykhodtseva N, Zhang Y-Z, McDannold N. Evaluation of permeability, doxorubicin delivery, and drug retention in a rat brain tumor model after ultrasound-induced blood-tumor barrier disruption. J Control Release. 2017;250:77–85.
- 20.Fan C-H, Lin C-Y, Liu H-L, Yeh C-K. Ultrasound targeted CNS gene delivery for Parkinson’s disease treatment. J Control Release. 2017;261:246–62.
- 21.Zhu L, Zhao H, Zhou Z, Xia Y, Wang Z, Ran H, et al. Peptide-functionalized phase-transformation nanoparticles for low intensity focused ultrasound-assisted tumor imaging and therapy. Nano Lett. 2018;18(3):1831–41.
- 22.Zlokovic BV. The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron. 2008;57(2):178–201.
- 23.Yoo SS, Bystritsky A, Lee JH, Zhang Y, Fischer K, Min BK, et al. Focused ultrasound modulates region-specific brain activity. Neuroimage. 2011;56(3):1267–75.
- 24.Kovacs ZI, Kim S, Jikaria N, Qureshi F, Milo B, Lewis BK, et al. Disrupting the blood-brain barrier by focused ultrasound induces sterile inflammation. Proc Natl Acad Sci U S A. 2017;114(1):E75–e84.
- 25.Tufail Y, Matyushov A, Baldwin N, Tauchmann ML, Georges J, Yoshihiro A, et al. Transcranial pulsed ultrasound stimulates intact brain circuits. Neuron. 2010;66(5):681–94.
- 26.Burgess A, Dubey S, Yeung S, Hough O, Eterman N, Aubert I, et al. Alzheimer disease in a mouse model: MR imaging–guided focused ultrasound targeted to the hippocampus opens the blood-brain barrier and improves pathologic abnormalities and behavior. Radiology. 2014;273(3):736–45.
- 27.Jordao JF, Thevenot E, Markham-Coultes K, Scarcelli T, Weng YQ, Xhima K, et al. Amyloid-beta plaque reduction, endogenous antibody delivery and glial activation by brain-targeted, transcranial focused ultrasound. Exp Neurol. 2013;248:16–29.
- 28.Leinenga G, Gotz J. Scanning ultrasound removes amyloid-beta and restores memory in an Alzheimer’s disease mouse model. Sci Transl Med. 2015;7(278):278ra33.
- 29.Scarcelli T, Jordao JF, O’Reilly MA, Ellens N, Hynynen K, Aubert I. Stimulation of hippocampal neurogenesis by transcranial focused ultrasound and microbubbles in adult mice. Brain Stimul. 2014;7(2):304–7.
- 30.Mooney SJ, Shah K, Yeung S, Burgess A, Aubert I, Hynynen K. Focused ultrasound-induced neurogenesis requires an increase in blood-brain barrier permeability. PLoS One. 2016;11(7):e0159892.
- 31.Moreno-Jiménez EP, Flor-García M, Terreros-Roncal J, Rábano A, Cafini F, Pallas-Bazarra N, et al. Adult hippocampal neurogenesis is abundant in neurologically healthy subjects and drops sharply in patients with Alzheimer’s disease. Nat Med. 2019. https://doi.org/10.1038/s41591-019-0375-9.
- 32.Wiley RG, Oeltmann TN, Lappi DA. Immunolesioning: selective destruction of neurons using immunotoxin to rat NGF receptor. Brain Res. 1991;562(1):149–53.
- 33.Lee JE, Jeong da U, Lee J, Chang WS, Chang JW. The effect of nucleus basalis magnocellularis deep brain stimulation on memory function in a rat model of dementia. BMC Neurol. 2016;16:6.
- 34.Cho JS, Lee J, Jeong DU, Kim HW, Chang WS, Moon J, et al. Effect of placenta-derived mesenchymal stem cells in a dementia rat model via microglial mediation: a comparison between stem cell transplant methods. Yonsei Med J. 2018;59(3):406–15.
- 35.Jeong DU, Lee J, Chang WS, Chang JW. Identifying the appropriate time for deep brain stimulation to achieve spatial memory improvement on the Morris water maze. BMC Neurosci. 2017;18(1):29.
- 36.Jeong DU, Chang WS, Hwang YS, Lee D, Chang JW. Decrease of GABAergic markers and arc protein expression in the frontal cortex by intraventricular 192 IgG-saporin. Dement Geriatr Cogn Disord. 2011;32(1):70–8.
- 37.Choi BY, Kim IY, Kim JH, Lee BE, Lee SH, Kho AR, et al. Administration of zinc plus cyclo-(His-Pro) increases hippocampal neurogenesis in rats during the early phase of streptozotocin-induced diabetes. Int J Mol Sci. 2017;18(1). https://www.mdpi.com/1422-0067/18/1/73.
- 38.Ellman GL, Courtney KD, Andres V Jr, Feather-Stone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol. 1961;7:88–95.
- 39.Yang B, Ren Q, Jc Z, Chen QX, Hashimoto K. Altered expression of BDNF, BDNF pro-peptide and their precursor proBDNF in brain and liver tissues from psychiatric disorders: rethinking the brain–liver axis. Transl Psychiatry. 2017;7(5):e1128-e.
- 40.Jerusalinsky D, Kornisiuk E, Izquierdo I. Cholinergic neurotransmission and synaptic plasticity concerning memory processing. Neurochem Res. 1997;22(4):507–15.
- 41.Hasselmo ME. The role of acetylcholine in learning and memory. Curr Opin Neurobiol. 2006;16(6):710–5.
- 42.Book AA, Wiley RG, Schweitzer JB. Specificity of 192 IgG-saporin for NGF receptor-positive cholinergic basal forebrain neurons in the rat. Brain Res. 1992;590(1–2):350–5.
- 43.Heckers S, Ohtake T, Wiley RG, Lappi DA, Geula C, Mesulam MM. Complete and selective cholinergic denervation of rat neocortex and hippocampus but not amygdala by an immunotoxin against the p75 NGF receptor. J Neurosci. 1994;14(3 Pt 1):1271–89.
- 44.Jeong DU, Lee JE, Lee SE, Chang WS, Kim SJ, Chang JW. Improvements in memory after medial septum stimulation are associated with changes in hippocampal cholinergic activity and neurogenesis. Biomed Res Int. 2014;2014:568587.
- 45.Jeong da U, Oh JH, Lee JE, Lee J, Cho ZH, Chang JW, et al. Basal forebrain cholinergic deficits reduce glucose metabolism and function of cholinergic and GABAergic systems in the cingulate cortex. Yonsei Med J. 2016;57(1):165–72.
- 46.Cohen JE, Zimmerman G, Melamed-Book N, Friedman A, Dori A, Soreq H. Transgenic inactivation of acetylcholinesterase impairs homeostasis in mouse hippocampal granule cells. Hippocampus. 2008;18(2):182–92.
- 47.Blitzer RD, Gil O, Landau EM. Cholinergic stimulation enhances long-term potentiation in the CA1 region of rat hippocampus. Neurosci Lett. 1990;119(2):207–10.
- 48.Shimoshige Y, Maeda T, Kaneko S, Akaike A, Satoh M. Involvement of M2 receptor in an enhancement of long-term potentiation by carbachol in Schaffer collateral-CA1 synapses of hippocampal slices. Neurosci Res. 1997;27(2):175–80.
- 49.Monteggia LM, Barrot M, Powell CM, Berton O, Galanis V, Gemelli T, et al. Essential role of brain-derived neurotrophic factor in adult hippocampal function. Proc Natl Acad Sci. 2004;101(29):10827–32.
- 50.Taliaz D, Stall N, Dar DE, Zangen A. Knockdown of brain-derived neurotrophic factor in specific brain sites precipitates behaviors associated with depression and reduces neurogenesis. Mol Psychiatry. 2009;15:80.
- 51.Zuccato C, Cattaneo E. Brain-derived neurotrophic factor in neurodegenerative diseases. Nat Rev Neurol. 2009;5(6):311.
- 52.Cooper-Kuhn CM, Winkler J, Kuhn HG. Decreased neurogenesis after cholinergic forebrain lesion in the adult rat. J Neurosci Res. 2004;77(2):155–65.
- 53.Rossi C, Angelucci A, Costantin L, Braschi C, Mazzantini M, Babbini F, et al. Brain-derived neurotrophic factor (BDNF) is required for the enhancement of hippocampal neurogenesis following environmental enrichment. Eur J Neurosci. 2006;24(7):1850–6.
- 54.Wrenn CC, Wiley RG. The behavioral functions of the cholinergic basal forebrain: lessons from 192 IgG-saporin. Int J Dev Neurosci. 1998;16(7–8):595–602.
- 55.Gil-Bea FJ, Solas M, Mateos L, Winblad B, Ramirez MJ, Cedazo-Minguez A. Cholinergic hypofunction impairs memory acquisition possibly through hippocampal Arc and BDNF downregulation. Hippocampus. 2011;21(9):999–1009.
- 56.Waterhouse EG, An JJ, Orefice LL, Baydyuk M, Liao G-Y, Zheng K, et al. BDNF promotes differentiation and maturation of adult-born neurons through GABAergic transmission. J Neurosci. 2012;32(41):14318–30.
- 57.Mousavi K, Jasmin BJ. BDNF is expressed in skeletal muscle satellite cells and inhibits myogenic differentiation. J Neurosci. 2006;26(21):5739–49.
- 58.Reichardt LF. Neurotrophin-regulated signalling pathways. Philos Trans R Soc London B: Biol Sci. 2006;361(1473):1545–64.
- 59.Bramham CR, Messaoudi E. BDNF function in adult synaptic plasticity: the synaptic consolidation hypothesis. Prog Neurobiol. 2005;76(2):99–125.
- 60.Scharfman H, Goodman J, Macleod A, Phani S, Antonelli C, Croll S. Increased neurogenesis and the ectopic granule cells after intrahippocampal BDNF infusion in adult rats. Exp Neurol. 2005;192(2):348–56.
- 61.M-m P. Neurotrophins as synaptic modulators. Nat Rev Neurosci. 2001;2(1):24.
- 62.Impey S, Obrietan K, Wong ST, Poser S, Yano S, Wayman G, et al. Cross talk between ERK and PKA is required for Ca2+ stimulation of CREB-dependent transcription and ERK nuclear translocation. Neuron. 1998;21(4):869–83.
- 63.Veyrac A, Besnard A, Caboche J, Davis S, Laroche S. The transcription factor Zif268/Egr1, brain plasticity, and memory. Prog Mol Biol Transl Sci. 2014;122:89–129.
- 64.Duclot F, Kabbaj M. The role of early growth response 1 (EGR1) in brain plasticity and neuropsychiatric disorders. Front Behav Neurosci. 2017;11(35). https://www.frontiersin.org/articles/10.3389/fnbeh.2017.00035/full.
- 65.Choi SH, Bylykbashi E, Chatila ZK, Lee SW, Pulli B, Clemenson GD, et al. Combined adult neurogenesis and BDNF mimic exercise effects on cognition in an Alzheimer’s mouse model. Science. 2018;1(6406). https://science.sciencemag.org/content/361/6406/eaan8821.long.
- 66.Shin J, Kong C, Cho JS, Lee J, Koh CS, Yoon MS, et al. Focused ultrasound-mediated noninvasive blood-brain barrier modulation: preclinical examination of efficacy and safety in various sonication parameters. Neurosurg Focus. 2018;44(2):E15.



{kind=link}