










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



When we fall asleep, our brains are not merely offline, they’re busy organizing new memories–and now, scientists have gotten a glimpse of the process.
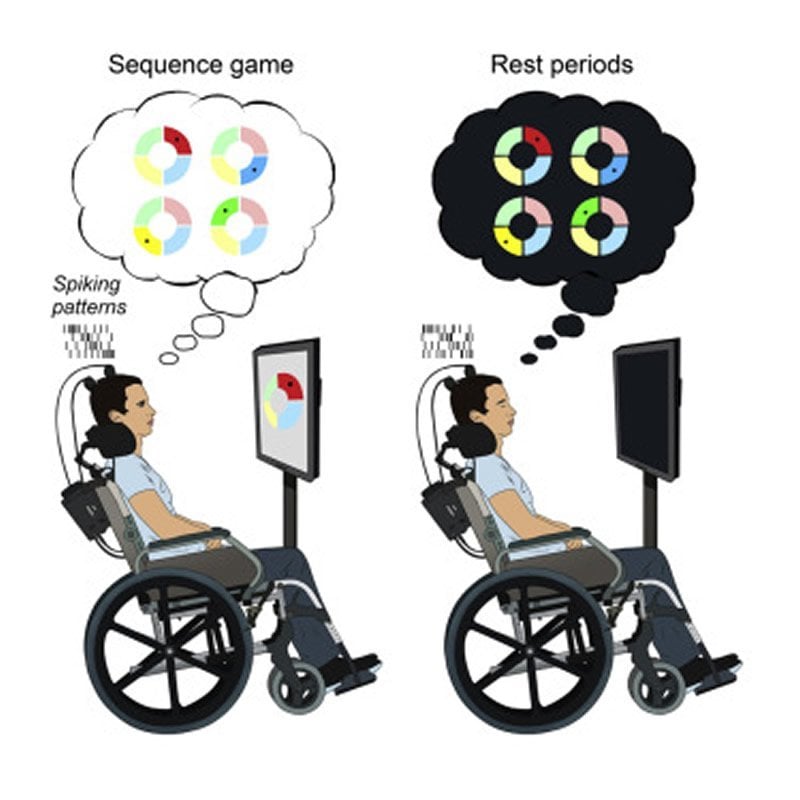
Researchers report in the journal Cell Reports on May 5 the first direct evidence that human brains replay waking experiences while asleep, seen in two participants with intracortical microelectrode arrays placed in their brains as part of a brain-computer interface pilot clinical trial.
During sleep, the brain replays neural firing patterns experienced while awake, also known as “offline replay.”
Replay is thought to underlie memory consolidation, the process by which recent memories acquire more permanence in their neural representation.
Scientists have previously observed replay in animals, but the study led by Jean-Baptiste Eichenlaub of Massachusetts General Hospital and Beata Jarosiewicz, formerly Research Assistant Professor at BrainGate, and now Senior Research Scientist at NeuroPace, tested whether the phenomenon happens in human brains as well.
The team asked the two participants to take a nap before and after playing a sequence-copying game, which is similar to the 80s hit game Simon. The video game had four color panels that lit up in different sequences for the players to repeat.
But instead of moving their arms, the participants played the game with their minds–imagining moving the cursor with their hands to different targets one by one, hitting the correct colors in the correct order as quickly as possible.
While the participants rested, played the game, and then rested again, the researchers recorded the spiking activity of large groups of individual neurons in their brains through an implanted multi-electrode array.
“There aren’t a lot of scenarios in which a person would have a multi-electrode array placed in their brain, where the electrodes are tiny enough to be able to detect the firing activity of individual neurons,” says co-first author Jarosiewicz.
Electrodes approved for medical indications, like those for treating Parkinson’s disease or epilepsy, are too big to track the spiking activity of single neurons. But the electrode arrays used in the BrainGate pilot clinical trials are the first to allow for such detailed neural recordings in the human brain. “That’s why this study is unprecedented,” she says.
BrainGate is an academic research consortium spanning Brown University, Massachusetts General Hospital, Case Western Reserve University, and Stanford University. Researchers at BrainGate are working to develop chronically implanted brain-computer interfaces to help people with severe motor disabilities regain communication and control by using their brain signals to move computer cursors, robotic arms, and other assistive devices.
In this study, the team observed the same neuronal firing patterns during both the gaming period and the post-game rest period. In other words, it’s as though the participants kept playing the Simon game after they were asleep, replaying the same patterns in their brain at a neuronal level. The findings provided direct evidence of learning-related replay in the human brain.
“This is the first piece of direct evidence that in humans, we also see replay during rest following learning that might help to consolidate those memories,” said Jarosiewicz. “All the replay-related memory consolidation mechanisms that we’ve studied in animals for all these decades might actually generalize to humans as well.”
The findings also open up more questions and future topics of study who want to understand the underlying mechanism by which replay enables memory consolidation.
The next step is to find evidence that replay actually has a causal role in the memory consolidation process. One way to do that would be to test whether there’s a relationship between the strength of the replay and the strength of post-nap memory recall.
Although scientists don’t fully understand how learning and memory consolidation work, a cascade of animal and human studies has shown that sleep plays a vital role. Getting a good night’s sleep “before a test and before important interviews” is beneficial for good cognitive performance, said Jarosiewicz. “We have good scientific evidence that sleep is very important in these processes.”
Funding: This work was supported by the U.S. Office of Naval Research, NINDS, NIDCD, the Department of Veterans Affairs, the Fyssen Foundation, and the Center for Neurorestoration and Neurotechnology from the United States (U.S.) Department of Veterans Affairs, Rehabilitation Research and Development Service. The authors would like to thank participants T9 and T10 and their families and caregivers.
WHAT ARE DREAMING AND MIND WANDERING?
“Dreaming” is usually understood as subjective mental experi- ences during sleep. Although most famously (and strongly) asso- ciated with REM sleep (Aserinsky and Kleitman, 1953; Dement and Kleitman, 1957), dream-like thought is also reported during other sleep stages (see Methods).
For several reasons, by “dreaming” we will generally be refer- ring to subjective reports drawn from REM sleep: for one thing, the majority of “dream” reports have been elicited from REM sleep-stage laboratory awakenings; further, only REM sleep shows a particularly strong correlation with dream mentation ( 80% of awakenings from REM sleep result in dream reports:
Hobson et al., 2000). For the purposes of the present paper, then, “dream- ing” refers to mentation reports from REM sleep.
“Undirected” thought is a similarly complex construct, and can be divided into several different categories (Christoff, 2012). “Mind wandering” (MW) and “stimulus-independent thought” (SIT), for instance, are typically defined as thinking that devi- ates from a particular task a subject is meant to be completing (McGuire et al., 1996; Mason et al., 2007; Christoff et al., 2009).
“Spontaneous thought,” on the other hand, is characterized rather by its undirected, effortless nature—more akin to the everyday concept of “daydreaming” (Singer, 1966; Klinger, 1990; Christoff,2012); no particular task, or deviation from it, is required.
Subtle differences are apparent: MW, for example, might be initiated deliberately (as when a subject decides to “tune out” during a boring task) rather than being “spontaneous.”
Nonetheless, these terms are often used interchangeably or with only minimal def- inition. Fluidity of terminology seems inevitable, however, in a relatively young field of inquiry (Christoff, 2012); moreover, the subjective content and neural basis of these states appear highly similar (compare, e.g., Singer and McCraven, 1961; Christoff et al., 2004, 2009; Stawarczyk et al., 2011).
We therefore use these terms relatively interchangeably throughout this paper. MW, spontaneous thought, or daydreaming, then, all refer to subjec- tive reports of undirected thoughts during wakefulness (whether deviating from, or in the complete absence of, a task).
THE DEFAULT MODE NETWORK (DMN) AND REM SLEEP
Though specific neural correlates of both daydreaming and dreaming remain somewhat elusive, these mental states, and their associated subjective content, are strongly correlated with the “resting state” and REM sleep, respectively (Aserinsky and Kleitman, 1953; Dement and Kleitman, 1957; Maquet et al., 1996; Mason et al., 2007; Christoff et al., 2009; Andrews-Hanna et al., 2010; Vanhaudenhuyse et al., 2010; Christoff, 2012; Hasenkampet al., 2012).
The default mode network (DMN) was discovered some- what serendipitously as a pattern of brain deactivations associated with the difference between brain activity during a quiet, resting state (the typical baseline condition for early fMRI studies) and a goal-oriented, directed task (Raichle et al., 2001).
Particular regions were consistently more active during “rest” than during goal-directed tasks of many kinds, suggesting a “default mode” network of regions active when a subject was “doing nothing” (Raichle et al., 2001; see Table 3 and Figure 2 for core regions of the DMN).
It quickly became clear, however, that physical “rest” by no means implied mental inactivity. With no explicit task, subjects almost immediately engaged in spontaneous thought, including daydreaming, planning for the future, recalling mem- ories, and so on (Gusnard et al., 2001).
Subsequent research has tied the subjective experience of MW to core DMN regions (Christoff et al., 2004, 2009; Mason et al., 2007; Andrews-Hanna et al., 2010; Vanhaudenhuyse et al., 2010; Hasenkamp et al., 2012).
Although regions beyond the DMN appear to also be recruited during MW (e.g., Christoff et al., 2009), the DMN still remains the most commonly used neural proxy for spontaneous thought (see also Methods).
REM sleep is initiated by a network of cells in the pons and nearby portions of the midbrain (Siegel, 2011), but involves a widespread recruitment of higher cortical brain regions (see our meta-analytic results, below, for regions of this theoretical REM network: Table 2 and Figure 1).
REM sleep recurs, in increas- ingly lengthy periods, approximately every 90 mins throughout the sleep cycle, overall constituting about 1.5–2 h of an aver- age night of sleep. Whereas non-REM (NREM) sleep stages are generally characterized by deactivation of many regions as com- pared to wakefulness (e.g., Kaufmann et al., 2006), REM is unique in that many brain regions are clearly more active than dur- ing wakefulness (Table 2, Figure 1). REM also appears to be the
most active state from the subjective point of view, with longer, more emotional, and more frequent dream mentation in REM than any other sleep stage (Hobson et al., 2000). REM therefore appears to be by far the best neural marker of dreaming, though it nonetheless remains problematic (see Methods).
SUBJECTIVE AND NEURAL SIMILARITIES BETWEEN DREAMING AND MIND WANDERING
A number of similarities in the subjective experience of dreaming and MW have previously been noted (see Section First-person Reports of Content from Mind Wandering and Dreaming for a detailed overview).
The possibility that the neural substrate of the DMN might be involved in, overlap with that of dream- ing/REM sleep has also been raised (Fosse and Domhoff, 2007; Pace-Schott, 2007, 2011; Ioannides et al., 2009; Nir and Tononi, 2010), but these comparisons too have remained qualitative: a quantitative meta-analysis has yet to be applied to the question of the similarity in neural substrates between DMN/MW and REM sleep/dreaming.
While major reviews and meta-analyses of the DMN have allowed for a tentative consensus regarding its neural basis (e.g., Buckner et al., 2008), a meta-analytic evaluation of brain activity during REM sleep has yet to be undertaken, making a direct comparison between brain activity in the two states difficult. The execution of such a meta-analysis of REM sleep was therefore a major goal of the present review.
FIRST-PERSON REPORTS OF CONTENT FROM MIND WANDERING AND DREAMING
Similarities in subjective content have been noted since the begin-ning of such research. For instance, the dreamlike nature of relaxed waking thought was documented in two early studies of what is now called MW, which were carried out in a sleep laboratory using EEG to monitor wakefulness. In both studies, participants were randomly asked to report anything that was going through their minds at the time of the probe. In the first
study, Foulkes and Scott (1973) found that 24% of thoughts could be categorized as visual, dramatic, and dreamlike. In a replication study, Foulkes and Fleisher (1975) discovered that 19% of reports were dreamlike.
The qualitative characteristics of dreaming have been inten- sively studied over the past century, yielding a considerable body of research from which some firm conclusions can be drawn regarding subjective content.
Though qualitative data on the content of MW is not nearly as comprehensive, a tentative overview is nonetheless possible. Although a comprehensive review of the lit- erature is beyond the scope of this article, we highlight consistent findings regarding the subjective content of dreaming and MW.
We focus on similarities in subject matter across several key areas, including sensory, emotional, fanciful, mnemonic, motivational, and social aspects, as well as addressing the presence or absence of cognitive control and metacognition. Various disparities and inconsistencies are addressed here, as well as in the Discussion.
SENSORY ASPECTS
The broadest similarity between dreaming and MW is perhaps also the most basic: the sensory building blocks of spontaneous thought in both waking and dreaming are overwhelmingly visual and auditory (though experiences in other sensory modalities are by no means precluded).
Dreaming
The largely audiovisual nature of dreaming was noted over two millennia ago by Artemidorus in his Oneirocritica (Harris- McCoy, 2012) and has been often replicated in contemporary research. For instance, a recent review of dream content (Schredl, 2010), based on more than 4000 dream reports from both laboratory awakenings and home dream diaries, found that visual content was present in 100%, and auditory content in 57%, of all reports (Table 1).
Other sense modalities (tactile, olfactory, gustatory, and nociceptive experiences), by contrast, were present in 1% or less of all reports. Indeed, the next most prominent modality after vision and audition was the vestibular sense: 8% of reports contained experiences of flying, floating, acceleration, etc. (Schredl, 2010).
Intriguingly, a comparison with studies of dream reports from more than a century ago shows a very similar trend: in the late nineteenth century, dream reports also almost always featured visual elements, followed by auditory imagery as the next most dominant aspect, and with the remaining senses accounting for very small percentages ( 1–7%) (Schwartz, 2000). This suggests that the sensory aspects of dreaming may be consis- tent cross-culturally (or at least, cross-temporally).

The apparent predominance of audio-visual content in dreams may underestimate other sensory modalities, however. A num- ber of studies sampling other sensory data revealed that, when prompted specifically for sensations such as pain (Nielsen et al., 1993; Raymond et al., 2002; Solomonova et al., 2008) or bodily orienting movements (Solomonova et al., 2008), participants often reported more information. To our knowledge, similar targeted sensory-content probes have not yet been undertaken during MW, precluding a more detailed comparison.
Mind wandering
Content findings from mind wandering are not usually directly comparable, since MW researchers have tended to focus on the intensity (rather than the prevalence) of audiovisual imagery, but available evidence suggests similar trends.
For example, factor analysis of nearly 1500 experience reports found that visual and auditory intensity are two of eight dimensions significantly characterizing spontaneous thoughts (Klinger and Cox, 1987).
A more recent study similarly found a very high prevalence of self-reported visual and auditory imagery during spontaneous thoughts (mean ratings of 4.22 and 4.02, respectively, on a 7- point Likert scale) (Stawarczyk et al., 2011).
Along these lines, a recent review concluded that the average spontaneous thought is moderately visual, contains at least some sound, and is very likely (74% of reports) to contain some form of interior monolog or “self-talk” (Klinger, 2008).
POSITIVE AND NEGATIVE EMOTIONALITY
Dreaming
It appears that most dreams ( 70–75% or more in adults) con- tain some emotion, though affect in dreams may not always be particularly strong, or appropriate to the context (see Domhoff, 2011, for a discussion). A number of studies have found a relative predominance of negative emotions in dreams, particularly when dreams are scored by judges rather than by dreamers (see Schredl, 2010, for a review).
Other studies, however, have found a balance of emotions in REM sleep dream reports, and one study (Fosse et al., 2001) found that joy/elation was in fact the most fre- quently reported emotion. An interesting study directly compared self-reports of dreaming vs. waking events, finding that negative emotion (particularly fear) was more prevalent during dreaming, and positive emotions more common in waking (Nielsen et al., 1991).
It may be, however, that more intense and negatively toned dreams are better remembered, and thus over-reported. Additionally, sampling techniques (e.g., laboratory awakenings vs. home dream journals) may contribute to differences in findings. Irrespective of these differences and methodological limitations, however, it is evident that both positive and negative emotions are ubiquitous during dreaming.
Mind-wandering
Though not yet extensively studied, emotion appears to be similarly ubiquitous during MW. One recent study, for instance,involving thousands of reports, found that the majority (69%) of spontaneous thought reports involved emotion (positive emo- tion in 42.5% of reports, negative emotion in 26.5%), whereas only 31% of reports were reported to be emotionally neutral (Killingsworth and Gilbert, 2010).
Though data are generally lacking, it is interesting to note that, in contrast to dream- ing, positive emotion appears to predominate during waking MW, and that many more waking spontaneous thoughts appear to be characterized by relatively flat (neutral) affect.
Also of interest is that the temporal focus of MW content appears to be more directed toward the past when negative mood has been experimentally induced (Smallwood and O’Connor, 2011).
IMPLAUSIBILITY AND BIZARRENESS
Though the typical spontaneous thought or dream is a rel- atively plausible simulation or elucidation of past memories, current events, or future plans, generally in line with the cur- rent concerns of the subject (see “Motivational Aspects,” below), nonetheless implausible and bizarre elements are common to both states—though their precise frequency remains disputed (Snyder, 1970; Dorus et al., 1971; Zadra and Domhoff, 2011). Examples are physically impossible or socially unlikely situa- tions, fanciful locales and characters, large discontinuities of time and/or space, and so on.
Dreaming
Depending on scoring criteria, it has been estimated that between 32% (Schredl, 2010) and 71% (Stenstrom, 2006) of dream reports feature bizarre or impossible elements. Despite widely varying estimates, however, there is general agreement that bizarre, incon- gruous or impossible elements are features of at least a substantial proportion of dreams. Differences in precise estimates are likely due to differing scoring procedures, as well as differences between dreamer- or judge-rated scores.
Mind-wandering
Though many MW episodes contain relatively realistic simula- tions of plausible events in the external world, nonetheless a substantial number ( 20% of reports) contain elements that are bizarre, implausible, or fanciful (defined as “departing sub- stantially from physical or social reality”) (Klinger and Cox, 1987; Kroll-Mensing, 1992; Klinger, 2008).
A more recent study has provided a general replication of earlier results: analyzing thousands of thoughts reported by 124 subjects, Kane et al. (2007) found that the average thought during MW contained a moderate level of fantasy (a mean of 3.77 on a 7-point scale).
In a rare study examining both waking fantasy and dream reports in the same 12 subjects, Williams et al. (1992) found that bizarre elements were about twice as prevalent in dreams vs. waking spontaneous thought.
In a similar vein, dream and daydream bizarreness have been studied in relation to “thick” vs. “thin” boundaries (Kunzendorf et al., 1997): though thin bound- ary personality was associated with more bizarre dreams and daydreams than thick boundary, dreams were scored more bizarre than daydreams across both personality types.
MNEMONIC FEATURES: CONTRIBUTIONS OF EPISODIC AND SEMANTIC MEMORY
Both dreaming and MW draw on episodic and semantic mem- ory sources as building blocks for novel subjective experiences. In this section we discuss the prevalence of past-oriented thoughts during both wakefulness and dreaming, and the potential contri- butions of both episodic and semantic memory to these states.
Dreaming
There is an intriguing literature suggesting that sleep, especially NREM sleep, may have a role in memory consolidation (Walker and Stickgold, 2006; Born and Wilhelm, 2012), including specific roles for REM sleep in consolidation of procedural (Smith et al., 2004) and emotional episodic (Nishida et al., 2009; Groch et al., 2013) memories.
A dynamic model of sleep-dependent memory consolidation and reconsolidation has recently been proposed, suggesting a complex relationship between sleep stages, memory types and their contribution to cognitive stability, flexibility and brain plasticity (Walker and Stickgold, 2006, 2010).
It is now well documented that dream content borrows from both temporally proximal and distal memories (Nielsen and Stenstrom, 2005). The most proximal memories (those from the previous day) are generally known as “day residue” (Freud, 1908), whereas the recurrence of elements 5–7 days following an experience is referred to as the “dream-lag” effect (Nielsen and Powell, 1989).
Personally relevant and emotionally salient events appear to manifest themselves in dream content as day residue and dream lag effects, but can also surface many years after initial encoding (Grenier et al., 2005).
The presence of emo- tional and personally relevant content in dreams may be related to the fact that emotional and impactful events are preferentially consolidated in memory (McGaugh et al., 2002; Nishida et al., 2009). While dreaming contains clear episodic autobiographical elements, memories only rarely get “replayed” in dream content
(∼1–2% of reports: Fosse et al., 2003).
Mind-wandering
MW appears to involve roughly equal percentages of thoughts about the past and future (Fransson, 2006), though some stud- ies suggest a “prospective bias” toward future-oriented thoughts (Smallwood et al., 2009; Andrews-Hanna et al., 2010; Stawarczyk et al., 2011), and also a past-bias inducible by negative mood (Smallwood and O’Connor, 2011).
Overall, however, it is clear that memories, particularly episodic ones, play a large role in spontaneous thought. Many studies have reported a high preva- lence ( 20% or more of reports) of past-focused MW (Fransson, 2006; Smallwood et al., 2009; Andrews-Hanna et al., 2010; Smallwood et al., 2011).
Indeed, one of the first studies to explore “resting state” activity using PET noted the similarities between such activity and episodic memory recall, as well as the fact that subjective reports of “rest” actually involved a large amount of past recollection and future planning (Andreasen et al., 1995).
Similar to dreaming, memories incorporated in waking MW tend to be of emotional and personally relevant material, and are often related to people’s current concerns (see section below on “Motivational Aspects”).
In summary, dreaming and MW both contain specific trace- able episodic and semantic memory sources, but very rarely
reproduce memories in their entirety. Rather, memories tend to reappear in novel, re-contextualized thoughts and scenarios (Nielsen and Stenstrom, 2005).
MOTIVATIONAL ASPECTS: CURRENT CONCERNS
Reports from both dreaming and MW show a strong proclivity to reflect the ongoing concerns of subjects, as well as elements of anticipating and planning for the future.
Dreaming
A wealth of data supports the notion that dreaming reflects ongo- ing waking concerns, desires, and experiences, in line with the “continuity hypothesis” of dreaming and waking mental activity (see, e.g., Domhoff, 1996, Ch. 8). For example, transient stress- ful situations, such as divorce (Cartwright et al., 1984) and grief (Kuiken et al., 2008) are also often present in dream reports in a general form.
Although dream content is often found to be thematically and emotionally consistent with the waking state of the dreamer, cer- tain activities prevalent in waking are only rarely found in dreams. These include cognitive activities such as reading, writing, and using a phone or a computer (Schredl, 2000).
Mind-wandering
Similar to dreaming, the content of waking MW also centers heav- ily on subjects’ current concerns (Klinger and Cox, 1987; Klinger, 2008; Andrews-Hanna, 2012).
Further, when the temporal focus of MW is examined, a large percentage ( 40% in one recent study: Andrews-Hanna et al., 2010) of spontaneous thoughts center around the present time 1 day, supporting the notion that MW strongly involves current concerns and experiences. Future-oriented thought is also incred- ibly common during MW (Smallwood et al., 2009; Andrews- Hanna et al., 2010; Stawarczyk et al., 2011), further supporting a role for MW in future-planning and potentially problem-solving.
Intriguingly, in one of the few neuroimaging studies to directly examine periods of MW, MW was associated with activations not only in the DMN but also in key executive prefrontal areas, including the dorsal anterior cingulate cortex and dorsolateral prefrontal cortex (Christoff et al., 2009). Such results are con- sistent with the prevalence of current concerns and unresolved issues in first-person content reports, and may reflect an ongoing (if unconscious) effort to address them (Christoff et al., 2009; see
also Discussion).
IMAGINED SOCIAL INTERACTION
Dreaming
Similar to waking life, dreaming is nearly always organized around interactions with others. Most dreams include other characters in some kind of relationship with the dreamer, or a generalized social situation (Hall and Van de Castle, 1966; Nielsen et al., 2003; Schredl et al., 2004; Zadra and Domhoff, 2011).
Social interactions in dreams follow a multitude of patterns, including threatening (Valli et al., 2005) and other- wise emotionally-charged situations (Cartwright et al., 1984). Occasionally, recognizable dream characters may change appear- ance or appear as a generalized entity, fused with features of other individuals.
Also of interest is the prevalence of “mental- izing” or use of “theory of mind” in dreaming—i.e., thinking about others’ thoughts, emotions and motivations (even though the “others” are of course merely imagined) (McNamara et al., 2007). In general, meaningful interactions with others may be one of the key factors guiding the progression of the dream narrative.
Mind wandering
First-person reports of MW often involve imagined social inter- actions with others, as well as thoughts about the intentions and beliefs of other people (Klinger, 2008). This has led to the general notion that “mentalizing” (i.e., thinking about the thoughts and minds of others) and the consideration of hypothetical social situ- ations may be key components of spontaneous thought (Buckner et al., 2008; Andrews-Hanna, 2012). Supporting this idea, numer- ous studies have found that brain activity underlying “theory of mind” and mentalizing overlaps significantly with DMN regions (see Buckner et al., 2008, for a review).
COGNITIVE CONTROL AND METACOGNITION
Dreaming
A singular aspect of dreams is the seemingly total lack of metacog- nitive awareness in the dream state. One experiences a com- plex simulation of oft-bizarre experiences, but without the overt
capacity to reflect on the bizarre state of affairs the mind and body are actually in see, e.g., Rechtschaffen (1978). Intriguingly, it appears that well-trained, or talented, individuals can develop metacognitive awareness of the dream state, becoming “lucid” in the dream and sometimes even directing its course and content (Dresler et al., 2012).
The exceptional nature of “lucid” dream- ing, however, serves to prove the rule of the general lack of control and metacognitive awareness in ordinary dreaming, a character- istic likely attributable to the deactivation of numerous prefrontal cortical regions during REM sleep (see our results in Table 2 and Figure 1; also Hobson et al., 2000; Muzur et al., 2002).
Mind wandering
A lack of explicit goals, and an unawareness that one is even daydreaming or has deviated from the task at hand, are typi- cal of MW (Schooler et al., 2011). But although MW tends to be less characterized by intentional thought and self-reflective awareness, this is not always the case.
A recent study from our group, for instance, found that subjects who were probed at ran- dom intervals reported being unaware that they had been mind wandering about half (45%) of the time.
One’s impression of the “controllability” of a segment of MW also varies widely, from a sense of being able to end it at any time, to being com- pletely absorbed in and swept along by a daydream (Klinger, 1978, 2008; Klinger and Cox, 1987; Kroll-Mensing, 1992; Klinger and Kroll-Mensing, 1995).
Collectively, these results suggest that cog- nitive control and metacognitive awareness in MW lie somewhere between the relative lucidity and self-reflectiveness of normal waking thought and behavior, and the near-total lack of con- trol and metacognitive nescience characteristic of regular (i.e., non-lucid) dreams. See the Discussion for an elaboration of this theme.


Source:
Cell Press



{kind=link}