











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Allergic reactions can occur without being triggered by an allergen such as grass or birch pollen – it is enough for the patient to be back in the same place where she was previously exposed to the allergen, as researchers at the University of Tübingen have found.
However, this kind of conditioned reaction to an intrinsically neutral and harmless situation only happens after a sleep phase that follows the conditioning.
The new study on the influence of psychological factors on allergic reactions was led by Dr. Luciana Besedovsky and Professor Jan Born from the Institute of Medical Psychology and Behavioral Neurobiology.
The findings go some way toward explaining why allergic reactions are frequently observed as a form of placebo reaction when the original allergen is not present. The study has been published in the latest Proceedings of the National Academy of Sciences.
The research team recruited volunteers with allergic rhinitis, who were then given a nasal spray containing their respective allergens (grass or birch pollen) in a neutral test room.
The strength of the allergic reaction occurring in the subjects was measured by the amount of a specific enzyme in the nasal secretion. Half of the subjects went to sleep for eight hours after this experiment, while the other half had to stay awake until the following evening.
One week later, the experiment was repeated in the same test room. But this time, no allergens were given.
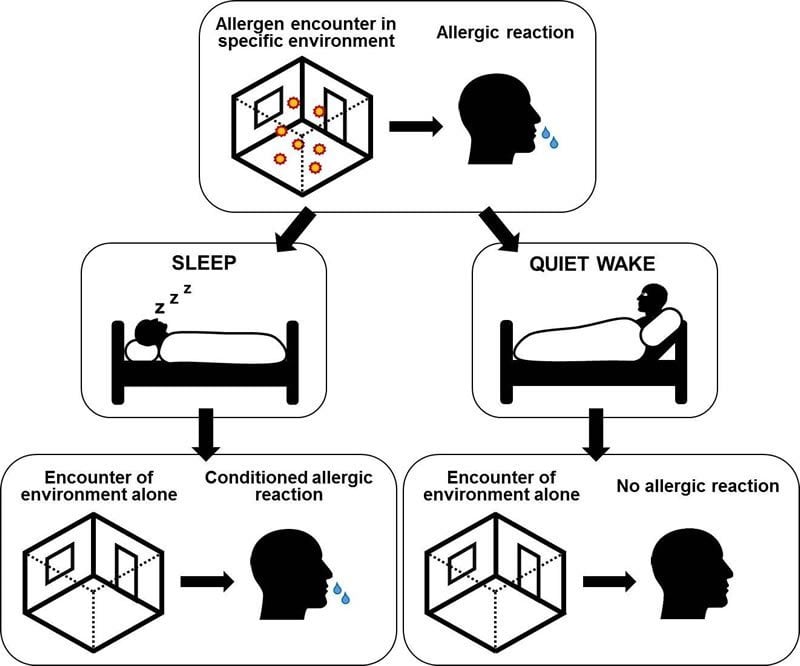
“The subjects reacted with allergic rhinitis shortly after entering the room—but only those from the group that slept,” says Besedovsky. subjects who remained awake did not have an allergic reaction upon returning to the room in which the experiment was conducted, Besedovsky adds. Nor did members of the group who slept suffer a reaction in another place they were taken to in the second week.
The brain is a fast learner
“Just as in a conventional learning process, the sleep phase played a decisive role in our study. Only through sleep did the brain firmly connect a certain environment with an allergic reaction,” says Jan Born, adding that this is the first experimental proof that a specific location alone can trigger an allergic reaction.
The researchers believe that – as in many memory-building processes – the hippocampus plays a role in conditioning the organism to respond to its environment. And the hippocampus functions in a sleep-dependent manner.
“It is astonishing how quickly the immune system learns the mismatched reaction. In the experiment, a single allergen dose was sufficient to link the allergic reaction with the environment,” Besedovsky says. Deciphering this learning mechanism is a boost to research into both allergies and sleep.
However, it is hard to draw simple conclusions on how to improve the situation of allergy sufferers. After all, they have to sleep – especially as sleep has a positive effect on other, helpful immune reactions.
In 1886, John N. MacKenzie published a famous case report on a woman who developed an asthmatic attack after seeing an artificial rose (1). More recently, Bennett G. Braun described several patients with multiple personalities in whom an allergic disorder was present with one but not the other personality (2).
“Placebo responses” in patients suffering from allergies are among the strongest observed in clinical studies (3, 4), and their great magnitude often results in insufficient statistical power for detecting verum effects (5).
These observations underscore the importance of psychological factors in allergic disorders, which are widespread with an increasing prevalence worldwide, exacting a high societal burden (6).
Immune responses, including allergic reactions, are known to be subject to Pavlovian conditioning; that is, after learning an association between an immune-active agent (e.g., an allergen) and an immunologically neutral stimulus (e.g., a distinct odor), the neutral stimulus alone can trigger the immune response (7⇓–9).
Two experimentally well-controlled studies in humans have demonstrated the development of conditioned allergic reactions after pairing an allergen with a specific cue (10, 11).
Another experimental study in humans added to these findings by showing that antiallergic responses also can be conditioned after pairing an antihistaminergic drug with a novel taste (12). These human experiments complement early studies in animals demonstrating conditioned mast cell responses (13, 14).
Conditioning processes can serve as mechanisms underlying the strong placebo responses in allergic diseases described above (15). Surprisingly, whereas the conditioning of diverse immune responses to distinct cues (i.e., cue conditioning) has been shown repeatedly, the specific role of context conditioning (i.e., the association of a response to its environmental context) in the Pavlovian learning of allergic responses has not yet been scrutinized experimentally (15–17), although context conditioning effects are known to substantially contribute to maladaptive responses in other domains (e.g., of fear and addiction behaviors) (18).
Sleep is generally thought of as an adaptive process, and one of its major functions is to support memory formation (19, 20).
Thus, sleep also might promote learned allergic responses, despite these responses being maladaptive. The role of sleep in the conditioning of immune responses has not yet been investigated. Against this backdrop, here we assessed the effects of sleep versus wakefulness after conditioning of an allergic rhinitis response in humans.
We were especially interested in comparing cue conditioning and context conditioning of allergic responses, because previous studies indicated that sleep selectively enhances context-conditioned, but not cue-conditioned, responses in the behavioral domain (21–24). Accordingly, here we expected postencoding sleep to specifically enhance context-conditioned, but not cue-conditioned, allergic responses.
Results
We subjected our otherwise healthy participants with clinically verified seasonal allergic rhinitis to a single-trial combined context/cue-conditioning procedure consisting of a Learning session and a Test session.
Both sessions comprised a 45-min Context phase during which the participant remained in a standardized experimental room, allowing encoding of and acclimatization to the environmental context (Materials and Methods and Fig. 1).
Then the Cue phase started with a 6-s presentation of the cue (conditioned stimulus [CS])—a distinctive odor (isobutyraldehyde)—which was immediately followed by the administration of the unconditioned stimulus (UCS) consisting of a nasal spray containing pollen allergens.
A physiological measure (mucosal tryptase) and a clinical symptom measure (Lebel score) of allergic rhinitis reactions were assessed during the Context phase (preodor measurements) and after cue-conditioning (postodor measurements) (Fig. 1).
Following the Learning session, the participants had either a night of regular sleep for 8 h (Sleep group) or stayed awake in bed in a semisupine position for the same 8-h period (Wake group). The Test session took place 1 wk later; this session was identical to the Learning session, except that the nasal spray presented during the Cue phase contained only a saline solution.

Study design. The Learning session started with a 45-min Context phase, during which the participant remained in a standardized experimental room to allow for encoding of the context and acclimatization to the room. In the ensuing Cue phase, the conditioned stimulus (CS)—a distinct odor—was delivered by holding a bottle containing a solution of isobutyraldehyde for 6 s below the participant’s nose. Immediately thereafter, the unconditioned stimulus (UCS)—pollen allergens delivered via a nasal spray—was presented. Allergic reactions (assessed by mucosal tryptase levels and the Lebel score) were determined at the end of the Context phase (preodor measurements) and at 5 min after presentation of the CS and UCS (postodor measurements). On the night after the Learning session, one-half of the participants slept during an 8-h period in the sleep laboratory, while the other half stayed awake (until the next evening). At 1 wk later, the Test session took place in the same standardized experimental room following identical procedures, except that the nasal spray contained only saline solution (Sal) without the UCS. Context-conditioned responses were assessed by comparing values at the end of the Context phase (preodor measures) of the Test session with those of the Learning session (baseline). Cue-conditioned responses were assessed by comparing postodor measures with preodor measures of the Test session.
Polysomnography data showed that participants in the Sleep group slept on average for 465 min (Table 1 provides sleep parameters), and that sleep architecture was comparable to that of healthy individuals examined in other studies from our laboratory as well as other laboratories (25, 26).
Participants in the Wake group were under constant supervision by the experimenter to ensure that they did not fall asleep during the sleep deprivation period between 23:00 and 7:00 h. Actigraphy data confirmed that none of the participants slept during the day after the sleep deprivation night, except for one participant who might have fallen asleep for approximately 90 min while watching TV in the afternoon of this day.
Basic physiological parameters, including heart rate, blood pressure, and body temperature, were not significantly different between the groups in the evening before the sleep manipulation (SI Appendix, Table S1).
Table 1.
Sleep parameters
| Parameter | Duration, min, mean ± SEM | Percentage of total sleep time, mean ± SEM |
| Total sleep time | 464.8 ± 3.3 | 100 |
| S1 | 39.9 ± 3.2 | 8.6 ± 0.7 |
| S2 | 226.6 ± 11.2 | 48.6 ± 2.2 |
| SWS | 80.5 ± 4.5 | 17.3 ± 0.9 |
| REM | 83.1 ± 6.2 | 17.9 ± 1.3 |
| WASO | 28.9 ± 11.9 | 6.3 ± 2.7 |
- S1, sleep stage 1; S2, sleep stage 2; SWS, slow-wave sleep, REM, rapid eye movement sleep; WASO, wake after sleep onsettblBody.
- n = 12; for one participant, the EEG could not be completely scored due to quality problems, but the part that could be scored (>6 h) indicated regular sleep.
During the Learning session, all participants developed an allergic reaction to the UCS, confirming their allergy (P < 0.001 for increases in tryptase levels and the Lebel score after UCS presentation—i.e., postodor values—with reference to values before UCS presentation—i.e., preodor values—serving as baseline) (SI Appendix, Fig. S1).
There were no significant differences between the Sleep and Wake groups in terms of tryptase levels and Lebel score in the Learning session (main effect “Sleep/Wake” and “Sleep/Wake” × “Preodor/Postodor” interaction; P > 0.832).
During the Test session 1 wk later, participants showed already in the Context phase (i.e., before the odor presentation) distinctly increased tryptase levels compared with preodor levels of the Learning session, demonstrating a conditioned response to the context (main effect “Session”: F(1,21) = 6.957, P = 0.015).
Importantly, the context-induced increase in tryptase levels occurred only in the participants who had slept after the Learning session (t(1,11) = 2.693, P = 0.021, d = 0.74) and was entirely absent in the Wake group (P = 0.341 for the post hoc pairwise t test; F(1,21) = 6.237, P = 0.021 for “Sleep/Wake” × “Session” interaction) (Fig. 2A and SI Appendix, Fig. S2A).
There was no significant difference in preodor values between the Learning and Test sessions for the Lebel score (P > 0.336 for main effect “Session” and “Session” × “Sleep/Wake” interaction) (Fig. 2A and SI Appendix, Fig. S2A).

Context-conditioned and cue-conditioned allergic responses measured after a single pairing of an environmental context and an odor cue (CS) with the administration of pollen allergens (UCS). (A) Mean ± SEM values of mucosal tryptase levels (Left) and the Lebel score (Right) for preodor measurements at the Learning session (empty bars) and at the Test session (black bars) (i.e., context-conditioning effect). n = 12 and n = 13 for tryptase levels and Lebel scores, respectively, of the Sleep group; n = 11 and n = 12 for tryptase levels and Lebel scores, respectively, of the Wake group. *P < 0.05 for ANOVA “Session” (Learning vs. Test session) × “Sleep/Wake” interaction and post hoc two-sided paired t test. n.s., not significant. (B) Mean ± SEM values of mucosal tryptase levels (Left) and the Lebel score (Right) before (empty bars) and at 5 min after (black bars) presentation of the odor cue during the Test session (i.e., cue-conditioning effect). n = 23 and n = 25 for tryptase levels and Lebel scores, respectively. ***P < 0.001, *P < 0.05 for ANOVA main effect of the factor “Preodor/Postodor” (preodor vs. postodor presentation). Because the Sleep and Wake groups did not differ in terms of the context-conditioning effect of the Lebel score (P > 0.928 for ANOVA “Session” × “Sleep/Wake” interaction and ANOVA main effect “Sleep/Wake”) and in terms of the cue-conditioning effects of both parameters (P > 0.526 for ANOVA “Preodor/Postodor” × “Sleep/Wake” interactions and ANOVA main effect “Sleep/Wake”) data for these effects are shown collapsed across both groups.
The data from the Test session also revealed a distinct cue-conditioned response to the odor cue, as evidenced by significant increases in postodor tryptase levels and the Lebel score compared with preodor values (main effect “Preodor/Postodor”: F(1,21) = 4.567, P = 0.045, d = 0.64 for tryptase levels; F(1,23) = 18.232, P < 0.001, d = 1.08 for the Lebel score) (Fig. 2B and SI Appendix, Fig. S2B). Notably, unlike the context-conditioned response, this cue-conditioned allergic response was independent of whether or not the participants had slept on the postconditioning night (“Sleep/Wake” × “Preodor/Postodor” interaction: P > 0.526 for both parameters).
To validate that the context-conditioning effect observed in the Sleep group was specific to the environmental context in which the learning had taken place, we added a “Context control” group, which was subjected to the same experimental procedure as the Sleep group of the main experiment (including the sleep period after conditioning), except that the Test session took place in a different environmental context than the Learning session.
As predicted, participants in this group did not show an increase in preodor tryptase levels during the Test session compared with preodor levels of the Learning session. Thus, no context-conditioned response was evident when the context of the Test session differed from that of the Learning session, even though the participants slept after the Learning session (P = 0.713 for the pairwise t test; P = 0.018 for the contrast between the Sleep group and the Context control and Wake groups) (Fig. 3 and SI Appendix, Fig. S3). The data for the Context control group confirmed a cue-conditioned response to the odor (details in SI Appendix, Fig. S4).

Context-conditioned allergic responses after regular sleep are abolished if the Test session takes place in a context different from that of the Learning session. Mean ± SEM values of mucosal tryptase levels (Left) and the Lebel score (Right) for preodor measurements at the Learning session (empty bars) and at the Test session (black bars), which took place in an environmental context different from that during learning. Note that no sleep/wake manipulation occurred in this Context control group; all participants of this group had regular sleep after the Learning session. P levels refer to two-sided paired t tests. n = 9 and n = 10 for tryptase levels and Lebel scores, respectively.
References
- J. N. Mackenzie, The production of the so-called “rose cold” by means of an artificial rose. Am. J. Med. Sci. 181, 45–56 (1886).Google Scholar
- B. G. Braun, Psychophysiologic phenomena in multiple personality and hypnosis. Am. J. Clin. Hypn. 26, 124–137 (1983).PubMedGoogle Scholar
- S. Gilles et al., The role of environmental factors in allergy: A critical reappraisal. Exp. Dermatol. 27, 1193–1200 (2018).Google Scholar
- A. Narkus et al., The placebo effect in allergen-specific immunotherapy trials. Clin. Transl. Allergy 3, 42 (2013).CrossRefPubMedGoogle Scholar
- A. del Cuvillo et al., Placebo effect in clinical trials involving patients with allergic rhinitis. J. Investig. Allergol. Clin. Immunol. 21 (suppl. 3), 40–45 (2011).Google Scholar
- D. Simon, Recent advances in clinical allergy and immunology. Int. Arch. Allergy Immunol. 177, 324–333 (2018).Google Scholar
- R. Ader,N. Cohen, Behaviorally conditioned immunosuppression. Psychosom. Med. 37, 333–340 (1975).Abstract/FREE Full TextGoogle Scholar
- F. Hucklebridge, Behavioral conditioning of the immune system. Int. Rev. Neurobiol. 52, 325–351 (2002).CrossRefPubMedGoogle Scholar
- M. Hadamitzky,L. Lückemann, G. Pacheco-López, M. Schedlowski, Pavlovian conditioning of immunological and neuroendocrine functions. Physiol. Rev. 100, 357–405 (2020).Google Scholar
- M. Gauci, A. J. Husband, H. Saxarra, M. G. King, Pavlovian conditioning of nasal tryptase release in human subjects with allergic rhinitis. Physiol. Behav. 55, 823–825 (1994).CrossRefPubMedGoogle Scholar
- J. E. Barrett, M. G. King, G. Pang, Conditioning rhinitis in allergic humans. Ann. N. Y. Acad. Sci. 917, 853–859 (2000).PubMedGoogle Scholar
- M. U. Goebel et al., Behavioral conditioning of immunosuppression is possible in humans. FASEB J. 16, 1869–1873 (2002).CrossRefPubMedGoogle Scholar
- M. Russell et al., Learned histamine release. Science 225, 733–734 (1984).Abstract/FREE Full TextGoogle Schola
- G. MacQueen, J. Marshall, M. Perdue, S. Siegel, J. Bienenstock, Pavlovian conditioning of rat mucosal mast cells to secrete rat mast cell protease II. Science 243, 83–85 (1989).Abstract/FREE Full TextGoogle Scholar
- S. Vits et al., Behavioural conditioning as the mediator of placebo responses in the immune system. Philos. Trans. R. Soc. Lond. B Biol. Sci. 366, 1799–1807 (2011).CrossRefPubMedGoogle Scholar
- E. Dekker, H. E. Pelser, J. Groen, Conditioning as a cause of asthmatic attacks; A laboratory study. J. Psychosom. Res. 2, 97–108 (1957).CrossRefPubMedGoogle Scholar
- D. R. Justesen, E. W. Braun, R. G. Garrison, R. B. Pendleton, Pharmacological differentiation of allergic and classically conditioned asthma in the guinea pig. Science 170, 864–866 (1970).Abstract/FREE Full TextGoogle Scholar
- S. Maren, K. L. Phan, I. Liberzon, The contextual brain: Implications for fear conditioning, extinction and psychopathology. Nat. Rev. Neurosci. 14, 417–428 (2013).CrossRefPubMedGoogle Scholar
- B. Rasch, J. Born, About sleep’s role in memory. Physiol. Rev. 93, 681–766 (2013).CrossRefPubMedGoogle Scholar
- L. Besedovsky, T. Lange, M. Haack, The sleep-immune crosstalk in health and disease. Physiol. Rev. 99, 1325–1380 (2019).CrossRefPubMedGoogle Scholar
- L. A. Graves, E. A. Heller, A. I. Pack, T. Abel, Sleep deprivation selectively impairs memory consolidation for contextual fear conditioning. Learn. Mem. 10, 168–176 (2003).Abstract/FREE Full TextGoogle Scholar
- D. J. Cai, T. Shuman, M. R. Gorman, J. R. Sage, S. G. Anagnostaras, Sleep selectively enhances hippocampus-dependent memory in mice. Behav. Neurosci. 123, 713–719 (2009).CrossRefPubMedGoogle Scholar
- C. G. Vecsey et al., Sleep deprivation impairs cAMP signalling in the hippocampus. Nature 461, 1122–1125 (2009).CrossRefPubMedGoogle Scholar
- C. V. Latchoumane, H. V. Ngo, J. Born, H. S. Shin, Thalamic spindles promote memory formation during sleep through triple phase-locking of cortical, thalamic, and hippocampal rhythms. Neuron 95, 424–435.e6 (2017).Google Scholar
- M. I. Boulos et al., Normal polysomnography parameters in healthy adults: A systematic review and meta-analysis. Lancet Respir. Med. 7, 533–543 (2019).Google Scholar
- M. Hirshkowitz, Normal human sleep: An overview. Med. Clin. North Am. 88, 551–565, vii (2004).CrossRefPubMedGoogle Scholar
- P. Ottenberg, M. Stein, J. Lewis, C. Hamilton, Learned asthma in the guinea pig. Psychosom. Med. 20, 395–400 (1958).Abstract/FREE Full TextGoogle Scholar
- S. K. Wise et al., International consensus statement on allergy and rhinology: Allergic rhinitis. Int. Forum Allergy Rhinol. 8, 108–352 (2018).Google Scholar
- C. R. Roxbury, M. Qiu, J. Shargorodsky, S. Y. Lin, Association between allergic rhinitis and poor sleep parameters in US adults. Int. Forum Allergy Rhinol. 8, 1098–1106 (2018).Google Scholar
- J. E. Carroll, M. R. Irwin, S. Stein Merkin, T. E. Seeman, Sleep and multisystem biological risk: A population-based study. PLoS One 10, e0118467 (2015).Google Scholar
- J. J. Kim, M. S. Fanselow, Modality-specific retrograde amnesia of fear. Science 256, 675–677 (1992).Abstract/FREE Full TextGoogle Scholar
- R. G. Phillips, J. E. LeDoux, Differential contribution of amygdala and hippocampus to cued and contextual fear conditioning. Behav. Neurosci. 106, 274–285 (1992).CrossRefPubMedGoogle Scholar
- N. R. Selden, B. J. Everitt, L. E. Jarrard, T. W. Robbins, Complementary roles for the amygdala and hippocampus in aversive conditioning to explicit and contextual cues. Neuroscience 42, 335–350 (1991).CrossRefPubMedGoogle Scholar
- A. Sawangjit et al., The hippocampus is crucial for forming non-hippocampal long-term memory during sleep. Nature 564, 109–113 (2018).CrossRefPubMedGoogle Scholar
- E. Sahin et al., Mechanism of action of allergen immunotherapy. Am. J. Rhinol. Allergy 30, 1–3 (2016).CrossRefGoogle Scholar
- E. F. Pace-Schott, P. W. Verga, T. S. Bennett, R. M. Spencer, Sleep promotes consolidation and generalization of extinction learning in simulated exposure therapy for spider fear. J. Psychiatr. Res. 46, 1036–1044 (2012).CrossRefPubMedGoogle Scholar
- S. Bandarian-Balooch, D. L. Neumann, M. J. Boschen, Exposure treatment in multiple contexts attenuates return of fear via renewal in high spider fearful individuals. J. Behav. Ther. Exp. Psychiatry 47, 138–144 (2015).Google Scholar
- J. Kirchhof et al., Learned immunosuppressive placebo responses in renal transplant patients. Proc. Natl. Acad. Sci. U.S.A. 115, 4223–4227 (2018).Abstract/FREE Full TextGoogle Scholar
- R. M. Smits, D. S. Veldhuijzen, N. M. Wulffraat, A. W. M. Evers, The role of placebo effects in immune-related conditions: Mechanisms and clinical considerations. Expert Rev. Clin. Immunol. 14, 761–770 (2018).Google Scholar
- T. L. Ben-Shaanan et al., Activation of the reward system boosts innate and adaptive immunity. Nat. Med. 22, 940–944 (2016).Google Scholar
- F. Benedetti et al., Conscious expectation and unconscious conditioning in analgesic, motor, and hormonal placebo/nocebo responses. J. Neurosci. 23, 4315–4323 (2003).Abstract/FREE Full TextGoogle Scholar
- E. Jensen-Jarolim, E. Untersmayr, Gender-medicine aspects in allergology. Allergy 63, 610–615 (2008).CrossRefPubMedGoogle Scholar
- M. Osman, Therapeutic implications of sex differences in asthma and atopy. Arch. Dis. Child. 88, 587–590 (2003).Abstract/FREE Full TextGoogle Scholar
- M. R. Milad et al., Fear conditioning and extinction: Influence of sex and menstrual cycle in healthy humans. Behav. Neurosci. 120, 1196–1203 (2006).CrossRefPubMedGoogle Scholar
- K. Löwgren et al., Performance in eyeblink conditioning is age and sex dependent. PLoS One 12, e0177849 (2017).Google Scholar
- V. Hajali, M. L. Andersen, S. S. Negah, V. Sheibani, Sex differences in sleep and sleep loss-induced cognitive deficits: The influence of gonadal hormones. Horm. Behav. 108, 50–61 (2019).Google Scholar
- S. Redline et al., The effects of age, sex, ethnicity, and sleep-disordered breathing on sleep architecture. Arch. Intern. Med. 164, 406–418 (2004).CrossRefPubMedGoogle Scholar
- M. Castells, L. B. Schwartz, Tryptase levels in nasal-lavage fluid as an indicator of the immediate allergic response. J. Allergy Clin. Immunol. 82, 348–355 (1988).CrossRefPubMedGoogle Scholar
- G. W. Scadding et al., Optimisation of grass pollen nasal allergen challenge for assessment of clinical and immunological outcomes. J. Immunol. Methods 384, 25–32 (2012).CrossRefPubMedGoogle Scholar
- P. H. Howarth et al., Objective monitoring of nasal airway inflammation in rhinitis. J. Allergy Clin. Immunol. 115 (3, suppl. 1), S414–S441 (2005).CrossRefPubMedGoogle Scholar
- B. Lebel et al., Correlation between symptoms and the threshold for release of mediators in nasal secretions during nasal challenge with grass-pollen grains. J. Allergy Clin. Immunol. 82, 869–877 (1988).CrossRefPubMedGoogle Scholar
- A. Rechtschaffen, A. Kales, A Manual of Standardized Terminology, Techniques and Scoring System for Sleep of Human Subjects (National Institutes of Health publication 204) (US Goverment Printing Office, Washington, DC, 1968).Google Scholar
Source:
University of Tubingen



{kind=link}