








")


 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Le reazioni allergiche possono verificarsi senza essere scatenate da un allergene come il polline di erba o betulla: è sufficiente che la paziente torni nello stesso posto in cui è stata precedentemente esposta all’allergene , come hanno scoperto i ricercatori dell’Università di Tubinga.
Tuttavia, questo tipo di reazione condizionata a una situazione intrinsecamente neutra e innocua si verifica solo dopo una fase del sonno che segue il condizionamento.
Il nuovo studio sull’influenza dei fattori psicologici sulle reazioni allergiche è stato condotto dalla dott.ssa Luciana Besedovsky e dal professor Jan Born dell’Istituto di psicologia medica e neurobiologia comportamentale.
I risultati vanno in qualche modo a spiegare perché le reazioni allergiche sono frequentemente osservate come una forma di reazione placebo quando l’allergene originale non è presente. Lo studio è stato pubblicato negli ultimi Atti della National Academy of Sciences.
Il team di ricerca ha reclutato volontari con rinite allergica, a cui è stato quindi somministrato uno spray nasale contenente i rispettivi allergeni (polline di erba o betulla) in una sala di prova neutrale.
La forza della reazione allergica che si verifica nei soggetti è stata misurata dalla quantità di un enzima specifico nella secrezione nasale. La metà dei soggetti è andata a dormire per otto ore dopo questo esperimento, mentre l’altra metà ha dovuto rimanere sveglia fino alla sera successiva.
Una settimana dopo, l’esperimento è stato ripetuto nella stessa sala prove. Ma questa volta non sono stati somministrati allergeni.
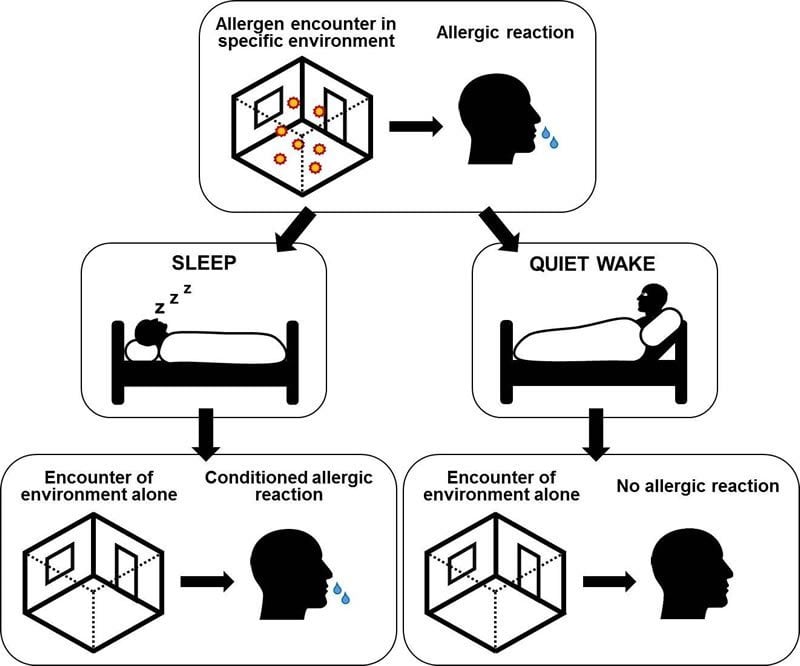
“I soggetti hanno reagito con la rinite allergica poco dopo essere entrati nella stanza, ma solo quelli del gruppo che hanno dormito”, afferma Besedovsky. i soggetti che sono rimasti svegli non hanno avuto una reazione allergica al ritorno nella stanza in cui è stato condotto l’esperimento, aggiunge Besedovsky. Né i membri del gruppo che hanno dormito hanno subito una reazione in un altro posto in cui sono stati portati nella seconda settimana.
Il cervello è uno studente veloce
“Proprio come in un processo di apprendimento convenzionale, la fase del sonno ha svolto un ruolo decisivo nel nostro studio. Solo attraverso il sonno il cervello ha collegato saldamente un determinato ambiente con una reazione allergica “, afferma Jan Born, aggiungendo che questa è la prima prova sperimentale che un luogo specifico da solo può scatenare una reazione allergica.
I ricercatori ritengono che – come in molti processi di costruzione della memoria – l’ippocampo abbia un ruolo nel condizionare l’organismo a rispondere al suo ambiente. E l’ippocampo funziona in modo dipendente dal sonno.
“È sorprendente quanto velocemente il sistema immunitario apprenda la reazione non corrispondente. Nell’esperimento, una singola dose di allergene è stata sufficiente per collegare la reazione allergica con l’ambiente “, afferma Besedovsky. Decifrare questo meccanismo di apprendimento è una spinta alla ricerca di allergie e sonno.
Tuttavia, è difficile trarre delle semplici conclusioni su come migliorare la situazione delle persone allergiche. Dopotutto, devono dormire, soprattutto perché il sonno ha un effetto positivo su altre, utili reazioni immunitarie.
Nel 1886, John N. MacKenzie pubblicò un famoso caso clinico su una donna che sviluppò un attacco asmatico dopo aver visto una rosa artificiale (1). Più recentemente, Bennett G. Braun ha descritto diversi pazienti con personalità multiple in cui era presente un disturbo allergico con l’una ma non con l’altra personalità (2).
Le “risposte del placebo” nei pazienti che soffrono di allergie sono tra le più forti osservate negli studi clinici (3, 4) e la loro grande ampiezza spesso si traduce in un potere statistico insufficiente per rilevare gli effetti verici (5).
Queste osservazioni sottolineano l’importanza dei fattori psicologici nei disturbi allergici, che sono diffusi con una prevalenza crescente in tutto il mondo, esigendo un elevato onere sociale (6).
Le risposte immunitarie, comprese le reazioni allergiche, sono note per essere soggette al condizionamento pavloviano; cioè, dopo aver appreso un’associazione tra un agente immuno-attivo (ad es. un allergene) e uno stimolo immunologicamente neutro (ad es. un odore distinto), lo stimolo neutro da solo può innescare la risposta immunitaria (7⇓-9).
Due studi sperimentalmente ben controllati sull’uomo hanno dimostrato lo sviluppo di reazioni allergiche condizionate dopo l’associazione di un allergene con un segnale specifico (10, 11).
Un altro studio sperimentale sull’uomo ha aggiunto a questi risultati dimostrando che anche le risposte antiallergiche possono essere condizionate dopo l’associazione di un farmaco antistaminergico con un nuovo gusto (12). Questi esperimenti sull’uomo completano i primi studi sugli animali che dimostrano risposte condizionate ai mastociti (13, 14).
I processi di condizionamento possono servire da meccanismi alla base delle forti risposte placebo nelle malattie allergiche sopra descritte (15). Sorprendentemente, mentre il condizionamento di diverse risposte immunitarie a segnali distinti (cioè, condizionamento di stecca) è stato dimostrato ripetutamente, il ruolo specifico del condizionamento del contesto (cioè l’associazione di una risposta al suo contesto ambientale) nell’apprendimento pavloviano delle risposte allergiche ha non è stato ancora esaminato sperimentalmente (15-17), anche se è noto che gli effetti del condizionamento del contesto contribuiscono in modo sostanziale alle risposte disadattive in altri settori (ad esempio, dei comportamenti di paura e dipendenza) (18).
Il sonno è generalmente considerato un processo adattivo e una delle sue principali funzioni è supportare la formazione della memoria (19, 20).
Pertanto, il sonno potrebbe anche promuovere risposte allergiche apprese, nonostante queste risposte siano disadattive. Il ruolo del sonno nel condizionamento delle risposte immunitarie non è stato ancora studiato. In questo contesto, qui abbiamo valutato gli effetti del sonno rispetto alla veglia dopo il condizionamento di una risposta allergica alla rinite nell’uomo.
Siamo stati particolarmente interessati a confrontare il condizionamento dell’indicazione e il condizionamento del contesto delle risposte allergiche, perché studi precedenti hanno indicato che il sonno migliora selettivamente le risposte condizionate dal contesto, ma non condizionate dall’indicazione, nel dominio comportamentale (21-24). Di conseguenza, qui ci aspettavamo che il sonno post-codifica migliorasse in modo specifico le risposte allergiche condizionate dal contesto, ma non condizionate dal segnale.
Risultati
Abbiamo sottoposto i nostri partecipanti altrimenti sani con rinite allergica stagionale verificata clinicamente a una procedura combinata a prova singola di contesto / indizio che consiste in una sessione di apprendimento e una sessione di test.
Entrambe le sessioni hanno comportato una fase di contesto di 45 minuti durante la quale il partecipante è rimasto in una stanza sperimentale standardizzata, consentendo la codifica e l’acclimatazione al contesto ambientale (materiali e metodi e figura 1).
Quindi la fase Cue è iniziata con una presentazione di 6 secondi dell’indicazione (stimolo condizionato [CS]) – un odore distintivo (isobutirraldeide) – che è stata immediatamente seguita dalla somministrazione dello stimolo incondizionato (UCS) costituito da uno spray nasale contenente polline allergeni.
Una misura fisiologica (triptasi della mucosa) e una misura dei sintomi clinici (punteggio di Lebel) delle reazioni di rinite allergica sono state valutate durante la fase di Contesto (misurazioni pre-odore) e dopo il condizionamento del segnale (misurazioni postodor) (Fig. 1).
Dopo la sessione di apprendimento, i partecipanti hanno avuto una notte di sonno regolare per 8 ore (gruppo del sonno) o sono rimasti svegli nel letto in una posizione semisupina per lo stesso periodo di 8 ore (gruppo del risveglio).
La sessione di test ha avuto luogo 1 settimana dopo; questa sessione era identica alla sessione di apprendimento, tranne per il fatto che lo spray nasale presentato durante la fase Cue conteneva solo una soluzione salina.

Progettazione dello studio. La sessione di apprendimento è iniziata con una fase contestuale di 45 minuti, durante la quale il partecipante è rimasto in una stanza sperimentale standardizzata per consentire la codifica del contesto e l’acclimatazione nella stanza. Nella successiva fase Cue, lo stimolo condizionato (CS) – un odore distinto – è stato erogato tenendo una bottiglia contenente una soluzione di isobutirraldeide per 6 secondi sotto il naso del partecipante. Immediatamente dopo, fu presentato lo stimolo incondizionato (UCS) – allergeni in polvere erogati tramite uno spray nasale. Le reazioni allergiche (valutate in base ai livelli di triptasi della mucosa e al punteggio di Lebel) sono state determinate al termine della fase di Contesto (misurazioni del preodor) e 5 minuti dopo la presentazione del CS e dell’UCS (misurazioni postodor). La notte dopo la sessione di apprendimento, metà dei partecipanti ha dormito durante un periodo di 8 ore nel laboratorio del sonno, mentre l’altra metà è rimasta sveglia (fino alla sera successiva). Dopo 1 settimana, la sessione del test si è svolta nella stessa stanza sperimentale standardizzata seguendo procedure identiche, tranne per il fatto che lo spray nasale conteneva solo soluzione salina (Sal) senza l’UCS. Le risposte condizionate dal contesto sono state valutate confrontando i valori alla fine della fase di contesto (misure preodor) della sessione di Test con quelle della sessione di Apprendimento (baseline). Le risposte condizionate sono state valutate confrontando le misure postodor con le misure preodor della sessione Test. tranne che lo spray nasale conteneva solo soluzione salina (Sal) senza l’UCS. Le risposte condizionate dal contesto sono state valutate confrontando i valori alla fine della fase di contesto (misure preodor) della sessione di Test con quelle della sessione di Apprendimento (baseline). Le risposte condizionate sono state valutate confrontando le misure postodor con le misure preodor della sessione Test. tranne che lo spray nasale conteneva solo soluzione salina (Sal) senza l’UCS. Le risposte condizionate dal contesto sono state valutate confrontando i valori alla fine della fase di contesto (misure preodor) della sessione di Test con quelle della sessione di Apprendimento (baseline). Le risposte condizionate sono state valutate confrontando le misure postodor con le misure preodor della sessione Test.
I dati di polisonnografia hanno mostrato che i partecipanti al gruppo del sonno hanno dormito in media per 465 minuti (la tabella 1 fornisce i parametri del sonno) e che l’architettura del sonno era paragonabile a quella di soggetti sani esaminati in altri studi dal nostro laboratorio e in altri laboratori (25, 26 ).
I partecipanti al gruppo Wake erano sotto la costante supervisione dello sperimentatore per assicurarsi che non si addormentassero durante il periodo di privazione del sonno tra le 23:00 e le 7:00 h.
I dati di Actigraphy hanno confermato che nessuno dei partecipanti ha dormito durante il giorno dopo la notte di privazione del sonno, ad eccezione di un partecipante che potrebbe essersi addormentato per circa 90 minuti mentre guardava la TV nel pomeriggio di questo giorno.
I parametri fisiologici di base, tra cui frequenza cardiaca, pressione sanguigna e temperatura corporea, non erano significativamente differenti tra i gruppi la sera prima della manipolazione del sonno (Appendice SI, Tabella S1).
Tabella 1. Parametri di sonno
| Parametro | Durata, min, media ± SEM | Percentuale del tempo di sonno totale, media ± SEM |
| Tempo di sonno totale | 464.8 ± 3.3 | 100 |
| S1 | 39.9 ± 3.2 | 8.6 ± 0.7 |
| S2 | 226.6 ± 11.2 | 48.6 ± 2.2 |
| SWS | 80.5 ± 4.5 | 17.3 ± 0.9 |
| Rem | 83.1 ± 6.2 | 17.9 ± 1.3 |
| WASO | 28.9 ± 11.9 | 6.3 ± 2.7 |
- S1, fase 1 del sonno; S2, fase 2 del sonno; SWS, sonno ad onde lente, REM, sonno a movimento rapido degli occhi; WASO, sveglia dopo il sonno onsettblBody.
- n = 12; per un partecipante, l’EEG non è stato completamente valutato a causa di problemi di qualità, ma la parte che poteva essere segnata (> 6 h) indicava un sonno regolare.
Durante la sessione di apprendimento, tutti i partecipanti hanno sviluppato una reazione allergica all’UCS, confermando la loro allergia (P <0,001 per gli aumenti dei livelli di triptasi e il punteggio Lebel dopo la presentazione dell’UCS, ovvero i valori postodor, con riferimento ai valori prima della presentazione dell’UCS, ovvero valori di pre-odore — che servono come base) (Appendice SI, Fig. S1).
Non ci sono state differenze significative tra i gruppi Sleep e Wake in termini di livelli di triptasi e punteggio Lebel nella sessione di apprendimento (effetto principale “Sleep / Wake” e “Sleep / Wake” × interazione “Preodor / Postodor”; P> 0,832).
Durante la sessione di test 1 settimana dopo, i partecipanti hanno mostrato già nella fase di contesto (cioè prima della presentazione degli odori) livelli nettamente superiori di triptasi rispetto ai livelli di preodor della sessione di apprendimento, dimostrando una risposta condizionata al contesto (effetto principale “Sessione”: F (1,21) = 6,957, P = 0,015).
È importante sottolineare che l’aumento indotto dal contesto nei livelli di triptasi si è verificato solo nei partecipanti che avevano dormito dopo la sessione di apprendimento (t (1,11) = 2,697, P = 0,021, d = 0,74) ed era completamente assente nel gruppo di veglia (P = 0,341 per il test t a coppie post hoc; F (1,21) = 6,237, P = 0,021 per l’interazione “Sleep / Wake” × “Sessione”) (Fig. 2A e Appendice SI, Fig. S2A).
Non vi era alcuna differenza significativa nei valori del preodor tra le sessioni di apprendimento e test per il punteggio Lebel (P> 0,336 per l’effetto principale “Sessione” e “Sessione” × Interazione “Sonno / veglia”) (Fig. 2A e Appendice SI, Fig. S2A).

Risposte allergiche condizionate dal contesto e condizionate misurate dopo una singola associazione di un contesto ambientale e un segnale di odore (CS) con la somministrazione di allergeni di polline (UCS). (A) Valori medi ± SEM dei livelli di triptasi della mucosa (sinistra) e punteggio Lebel (destra) per le misurazioni del preodor nella sessione di apprendimento (barre vuote) e nella sessione di test (barre nere) (cioè, effetto di condizionamento del contesto). n = 12 e n = 13 per i livelli di triptasi e i punteggi di Lebel, rispettivamente, del gruppo Sonno; n = 11 e n = 12 per i livelli di triptasi e i punteggi di Lebel, rispettivamente, del gruppo Wake. * P <0,05 per ANOVA “Sessione” (Apprendimento vs. sessione di test) × Interazione “Sleep / Wake” e test t bilaterale post hoc accoppiato. ns, non significativo. (B) Media ± valori di SEM dei livelli di triptasi della mucosa (a sinistra) e del punteggio di Lebel (a destra) prima (barre vuote) e 5 minuti dopo (barre nere) presentazione dell’odore cue durante la sessione di test (ad es. Condizionamento dell’indicazione effetto). n = 23 e n = 25 per i livelli di triptasi e i punteggi di Lebel, rispettivamente. *** P <0,001, * P <0,05 per l’effetto principale ANOVA del fattore “Preodor / Postodor” (presentazione preodor vs. postodor). Perché i gruppi Sleep and Wake non differivano in termini di effetto condizionamento del contesto del punteggio Lebel (P> 0.928 per interazione ANOVA “Session” × “Sleep / Wake” e effetto principale ANOVA “Sleep / Wake”) e in termini degli effetti di condizionamento di entrambi i parametri (P> 0,526 per le interazioni ANOVA “Preodor / Postodor” × “Sleep / Wake” e ANOVA effetto principale “Sleep / Wake”) per questi effetti sono mostrati collassati in entrambi i gruppi.
I dati della sessione di test hanno anche rivelato una risposta distinta condizionata dall’indicazione dell’odore, come evidenziato da significativi aumenti dei livelli di triptasi postodor e del punteggio di Lebel rispetto ai valori di preodor (effetto principale “Preodor / Postodor”: F (1,21 ) = 4.567, P = 0.045, d = 0.64 per livelli di triptasi; F (1,23) = 18,232, P <0,001, d = 1,08 per il punteggio di Lebel) (Fig. 2B e Appendice SI, Fig. S2B). In particolare, a differenza della risposta condizionata dal contesto, questa risposta allergica basata sul segnale era indipendente dal fatto che i partecipanti avessero dormito o meno nella notte post-condizionamento (interazione “Sleep / Wake” × “Preodor / Postodor”: P> 0,526 per entrambi i parametri) .
Per convalidare che l’effetto di condizionamento del contesto osservato nel gruppo del sonno era specifico del contesto ambientale in cui l’apprendimento aveva avuto luogo, abbiamo aggiunto un gruppo di “controllo del contesto”, che è stato sottoposto alla stessa procedura sperimentale del gruppo del sonno del esperimento principale (incluso il periodo di sonno dopo il condizionamento), tranne per il fatto che la sessione di test si è svolta in un contesto ambientale diverso rispetto alla sessione di apprendimento.
Come previsto, i partecipanti a questo gruppo non hanno mostrato un aumento dei livelli di pre-triptasi durante la sessione di test rispetto ai livelli di pre-odore della sessione di apprendimento.
Pertanto, nessuna risposta condizionata al contesto era evidente quando il contesto della sessione Test differiva da quello della sessione Learning, anche se i partecipanti hanno dormito dopo la sessione Learning (P = 0,713 per il test t a coppie; P = 0,018 per il contrasto tra il gruppo Sonno, il controllo Contesto e i gruppi Wake) (Fig. 3 e Appendice SI, Fig. S3). I dati per il gruppo di controllo del contesto hanno confermato una risposta basata sull’indicazione dell’odore (dettagli nell’appendice SI, Fig. S4).

Le risposte allergiche condizionate dal contesto dopo il sonno regolare vengono abolite se la sessione del test si svolge in un contesto diverso da quello della sessione di apprendimento. Valori medi ± SEM dei livelli di triptasi della mucosa (sinistra) e del punteggio Lebel (destra) per le misurazioni del preodor nella sessione di apprendimento (barre vuote) e nella sessione di test (barre nere), che si sono svolte in un contesto ambientale diverso da quello durante apprendimento. Si noti che non si è verificata alcuna manipolazione del sonno / veglia in questo gruppo di controllo del contesto; tutti i partecipanti di questo gruppo hanno dormito regolarmente dopo la sessione di apprendimento. I livelli di P si riferiscono a test t accoppiati su due lati. n = 9 e n = 10 rispettivamente per i livelli di triptasi e i punteggi di Lebel.
Riferimenti
- JN Mackenzie, La produzione del cosiddetto “freddo rosa” per mezzo di una rosa artificiale. Am. J. Med. Sci. 181, 45–56 (1886). Google Scholar
- BG Braun, fenomeni psicofisiologici nella personalità multipla e nell’ipnosi. Am. J. Clin. Hypn. 26, 124–137 (1983). PubMed Google Scholar
- S. Gilles et al ., Il ruolo dei fattori ambientali nell’allergia: una rivalutazione critica. Exp. Dermatol. 27, 1193-1200 (2018). Google Scholar
- A. Narkus et al ., L’effetto placebo negli studi di immunoterapia specifici per allergeni. Clin. Trad. Allergy 3, 42 (2013). CrossRef PubMed Google Scholar
- A. del Cuvillo et al ., Effetto placebo in studi clinici su pazienti con rinite allergica. J. Investig. Allergol. Clin. Immunol. 21 (suppl. 3), 40–45 (2011). Google Scholar
- D. Simon, recenti progressi nell’allergia clinica e nell’immunologia. Int. Arco. Allergy Immunol. 177, 324–333 (2018). Google Scholar
- R. Ader, N. Cohen, immunosoppressione comportamentale condizionata. Psychosom. Med. 37, 333–340 (1975). Estratto / LIBERO Testo completo Google Scholar
- F. Hucklebridge, Condizionamento comportamentale del sistema immunitario. Int. Rev. Neurobiol. 52, 325–351 (2002). CrossRef PubMed Google Scholar
- M. Hadamitzky, L. Lückemann, G. Pacheco-López, M. Schedlowski, condizionamento pavloviano delle funzioni immunologiche e neuroendocrine. Physiol. Rev. 100, 357–405 (2020). Google Scholar
- M. Gauci, AJ Marito, H. Saxarra, MG King, condizionamento pavloviano del rilascio di triptasi nasale in soggetti umani con rinite allergica. Physiol. Behav. 55, 823–825 (1994). CrossRef PubMed Google Scholar
- JE Barrett, MG King, G. Pang, Rinite condizionante nell’uomo allergico. Ann. NY Acad. Sci. 917, 853–859 (2000). PubMed Google Scholar
- MU Goebel et al ., Il condizionamento comportamentale dell’immunosoppressione è possibile nell’uomo. FASEB J. 16, 1869-1873 (2002). CrossRef PubMed Google Scholar
- M. Russell et al ., Imparato rilascio di istamina. Science 225, 733–734 (1984). Abstract / FREE Testo completo Google Schola
- G. MacQueen, J. Marshall, M. Perdue, S. Siegel, J. Bienenstock, condizionamento pavloviano dei mastociti della mucosa di ratto per secernere la proteasi dei mastociti di ratto II. Science 243, 83–85 (1989). Estratto / LIBERO Testo completo Google Scholar
- S. Vits et al ., Condizionamento comportamentale come mediatore delle risposte placebo nel sistema immunitario. Philos. Trans. R. Soc. Lond. B Biol. Sci. 366, 1799–1807 (2011). CrossRef PubMed Google Scholar
- E. Dekker, HE Pelser, J. Groen, Condizionamento come causa di attacchi asmatici; Uno studio di laboratorio. J. Psychosom. Res. 2, 97–108 (1957). CrossRef PubMed Google Scholar
- DR Justesen, EW Braun, RG Garrison, RB Pendleton, differenziazione farmacologica dell’asma allergico e classicamente condizionato nella cavia. Science 170, 864–866 (1970). Estratto / LIBERO Testo completo Google Scholar
- S. Maren, KL Phan, I. Liberzon, Il cervello contestuale: implicazioni per il condizionamento della paura, l’estinzione e la psicopatologia. Nat. Rev. Neurosci. 14, 417–428 (2013). CrossRef PubMed Google Scholar
- B. Rasch, J. Born, A proposito del ruolo del sonno nella memoria. Physiol. Rev. 93, 681–766 (2013). CrossRef PubMed Google Scholar
- L. Besedovsky, T. Lange, M. Haack, La diafonia immuno-sonno in salute e malattia. Physiol. Rev. 99, 1325-1380 (2019). CrossRef PubMed Google Scholar
- LA Graves, EA Heller, AI Pack, T. Abel, La privazione del sonno compromette selettivamente il consolidamento della memoria per il condizionamento contestuale della paura. Imparare. Mem. 10, 168-176 (2003). Estratto / LIBERO Testo completo Google Scholar
- DJ Cai, T. Shuman, MR Gorman, JR Sage, SG Anagnostaras, Sleep migliorano selettivamente la memoria dipendente dall’ippocampo nei topi. Behav. Neurosci. 123, 713–719 (2009). CrossRef PubMed Google Scholar
- CG Vecsey et al ., La privazione del sonno compromette la segnalazione del cAMP nell’ippocampo. Nature 461, 1122–1125 (2009). CrossRef PubMed Google Scholar
- CV Latchoumane, HV Ngo, J. Born, HS Shin, mandrini Thalamic promuovono la formazione della memoria durante il sonno attraverso il triplo blocco della fase dei ritmi corticali, talamici e ippocampali. Neurone 95, 424–435.e6 (2017). Google Scholar
- MI Boulos et al ., Parametri normali di polisonnografia in adulti sani: una revisione sistematica e una meta-analisi. Lancet Respir. Med. 7, 533-543 (2019). Google Scholar
- M. Hirshkowitz, Sonno umano normale: una panoramica. Med. Clin. Nord. 88, 551-565, vii (2004). CrossRef PubMed Google Scholar
- P. Ottenberg, M. Stein, J. Lewis, C. Hamilton, hanno imparato l’asma nella cavia. Psychosom. Med. 20, 395–400 (1958). Estratto / LIBERO Testo completo Google Scholar
- SK Wise et al ., Dichiarazione di consenso internazionale su allergia e rinologia: rinite allergica. Int. Forum Allergy Rhinol. 8, 108–352 (2018). Google Scholar
- CR Roxbury, M. Qiu, J. Shargorodsky, SY Lin, Associazione tra rinite allergica e parametri di sonno insufficiente negli adulti statunitensi. Int. Forum Allergy Rhinol. 8, 1098-1106 (2018). Google Scholar
- JE Carroll, MR Irwin, S. Stein Merkin, TE Seeman, Sonno e rischio biologico multisistemico: uno studio basato sulla popolazione. PLoS One 10, e0118467 (2015). Google Scholar
- JJ Kim, MS Fanselow, amnesia retrograda specifica della modalità. Science 256, 675–677 (1992). Estratto / LIBERO Testo completo Google Scholar
- RG Phillips, JE LeDoux, Contributo differenziale di amigdala e ippocampo al condizionamento della paura innescato e contestuale. Behav. Neurosci. 106, 274–285 (1992). CrossRef PubMed Google Scholar
- NR Selden, BJ Everitt, LE Jarrard, TW Robbins, ruoli complementari per l’amigdala e l’ippocampo nel condizionamento avverso a segnali espliciti e contestuali. Neuroscience 42, 335–350 (1991). CrossRef PubMed Google Scholar
- A. Sawangjit et al ., L’ippocampo è fondamentale per formare una memoria a lungo termine non ippocampale durante il sonno. Nature 564, 109-113 (2018). CrossRef PubMed Google Scholar
- E. Sahin et al ., Meccanismo d’azione dell’immunoterapia con allergeni. Am. J. Rhinol. Allergy 30, 1–3 (2016). CrossRef Google Scholar
- EF Pace-Schott, PW Verga, TS Bennett, RM Spencer, Sleep promuove il consolidamento e la generalizzazione dell’apprendimento dell’estinzione nella terapia dell’esposizione simulata per la paura del ragno. J. Psychiatr. Res. 46, 1036-1044 (2012). CrossRef PubMed Google Scholar
- S. Bandarian-Balooch, DL Neumann, MJ Boschen, Il trattamento dell’esposizione in più contesti attenua il ritorno della paura attraverso il rinnovamento in soggetti spaventosi di alto ragno. J. Behav. Ther. Exp. Psychiatry 47, 138-144 (2015). Google Scholar
- J. Kirchhof et al ., Imparate le risposte immunosoppressive al placebo nei pazienti con trapianto renale. Proc. Natl. Acad. Sci. USA 115, 4223–4227 (2018). Estratto / LIBERO Testo completo Google Scholar
- RM Smits, DS Veldhuijzen, NM Wulffraat, AWM Evers, Il ruolo degli effetti placebo in condizioni immuno-correlate: meccanismi e considerazioni cliniche. Esperto Rev. Clin. Immunol. 14, 761–770 (2018). Google Scholar
- TL Ben-Shaanan et al ., L’attivazione del sistema di ricompensa aumenta l’immunità innata e adattiva. Nat. Med. 22, 940-944 (2016). Google Scholar
- F. Benedetti et al ., Aspettativa cosciente e condizionamento inconscio nelle risposte placebo / nocebo analgesiche, motorie e ormonali. J. Neurosci. 23, 4315–4323 (2003). Estratto / LIBERO Testo completo Google Scholar
- E. Jensen-Jarolim, E. Untersmayr, Aspetti di medicina di genere in allergologia. Allergy 63, 610–615 (2008). CrossRef PubMed Google Scholar
- M. Osman, implicazioni terapeutiche delle differenze sessuali nell’asma e nell’atopia. Arco. Dis. Bambino. 88, 587-590 (2003). Estratto / LIBERO Testo completo Google Scholar
- MR Milad et al ., Condizionamento della paura ed estinzione: influenza del sesso e del ciclo mestruale in esseri umani sani. Behav. Neurosci. 120, 1196–1203 (2006). CrossRef PubMed Google Scholar
- K. Löwgren et al ., Le prestazioni nel condizionamento degli occhi sono dipendenti dall’età e dal sesso. PLoS One 12, e0177849 (2017). Google Scholar
- V. Hajali, ML Andersen, SS Negah, V. Sheibani, Differenze sessuali nel sonno e deficit cognitivi indotti dalla perdita di sonno: l’influenza degli ormoni gonadici. Horm. Behav. 108, 50–61 (2019). Google Scholar
- S. Redline et al ., Gli effetti dell’età, del sesso, dell’etnia e della respirazione disturbata dal sonno sull’architettura del sonno. Arco. Stagista. Med. 164, 406–418 (2004). CrossRef PubMed Google Scholar
- M. Castells, LB Schwartz, livelli di triptasi nel liquido di lavaggio nasale come indicatore della risposta allergica immediata. J. Allergy Clin. Immunol. 82, 348–355 (1988). CrossRef PubMed Google Scholar
- GW Scadding et al ., Ottimizzazione della sfida degli allergeni nasali del polline di erba per la valutazione degli esiti clinici e immunologici. J. Immunol. Metodi 384, 25–32 (2012). CrossRef PubMed Google Scholar
- PH Howarth et al ., Monitoraggio oggettivo dell’infiammazione delle vie aeree nasali nella rinite. J. Allergy Clin. Immunol. 115 (3, suppl.1), S414 – S441 (2005). CrossRef PubMed Google Scholar
- B. Lebel et al ., Correlazione tra sintomi e soglia di rilascio di mediatori nelle secrezioni nasali durante la sfida nasale con granuli di polline d’erba. J. Allergy Clin. Immunol. 82, 869–877 (1988). CrossRef PubMed Google Scholar
- A. Rechtschaffen, A. Kales, un manuale di terminologia standardizzata, tecniche e sistema di punteggio per il sonno di soggetti umani (pubblicazione 204 del National Institutes of Health) (US Goverment Printing Office, Washington, DC, 1968). Google Scholar
Fonte:
Università di Tubinga



{kind=link}