








")


 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Gli scienziati dell’USC e dell’IFOM Cancer Institute di Milano hanno scoperto che una dieta che imita il digiuno potrebbe essere più efficace nel trattamento di alcuni tipi di tumore se combinata con la vitamina C.
Negli studi sui topi, i ricercatori hanno scoperto che la combinazione ritardava la progressione del tumore in più modelli murini di carcinoma del colon-retto; in alcuni topi, ha causato la regressione della malattia. I risultati sono stati pubblicati sulla rivista Nature Communications .
“Per la prima volta, abbiamo dimostrato come un intervento completamente atossico possa efficacemente curare un cancro aggressivo”, ha dichiarato Valter Longo, autore senior dello studio e direttore dell’USC Longevity Institute presso la USC Leonard Davis School of Gerontology e professore di scienze biologiche all’USC Dornsife College of Letters, Arts and Sciences.
“Abbiamo preso due trattamenti che sono stati ampiamente studiati come interventi per ritardare l’invecchiamento – una dieta che imita il digiuno e la vitamina C – e li abbiamo combinati come un potente trattamento per il cancro.”
I ricercatori hanno affermato che mentre il digiuno rimane un’opzione stimolante per i malati di cancro, un’opzione più sicura e più fattibile è una dieta a basso contenuto calorico a base vegetale che induce le cellule a rispondere come se il corpo stesse digiunando.
I loro risultati suggeriscono che un trattamento a bassa tossicità della dieta che imita il digiuno più la vitamina C ha il potenziale per sostituire trattamenti più tossici.
I risultati di ricerche precedenti sul potenziale di lotta contro il cancro della vitamina C sono stati mescolati. Studi recenti, tuttavia, stanno iniziando a mostrare una certa efficacia, soprattutto in combinazione con la chemioterapia.
In questo nuovo studio, il team di ricerca voleva scoprire se una dieta che imitasse il digiuno potesse migliorare l’ azione di lotta contro il tumore alla vitamina C ad alte dosi creando un ambiente che sarebbe insostenibile per le cellule tumorali ma ancora sicuro per le cellule normali.
“Il nostro primo esperimento in vitro ha mostrato effetti notevoli”, ha detto Longo.
“Se usato da solo, la dieta che imita il digiuno o la vitamina C da sola ha ridotto la crescita delle cellule tumorali e causato un lieve aumento della morte delle cellule tumorali. Ma se usati insieme, hanno avuto un effetto drammatico, uccidendo quasi tutte le cellule cancerose. “
Longo e i suoi colleghi hanno rilevato questo forte effetto solo nelle cellule tumorali che avevano una mutazione che è considerata uno degli obiettivi più impegnativi nella ricerca sul cancro.
Queste mutazioni del gene KRAS segnalano che il corpo sta resistendo alla maggior parte dei trattamenti antitumorali e riducono il tasso di sopravvivenza di un paziente. Le mutazioni del KRAS si verificano in circa un quarto di tutti i tumori umani e si stima che si verifichino fino alla metà di tutti i tumori del colon-retto.
Lo studio ha anche fornito indizi sul perché precedenti studi sulla vitamina C come potenziale terapia antitumorale avevano mostrato un’efficacia limitata. Di per sé, un trattamento con vitamina C sembra innescare le cellule mutate dal KRAS per proteggere le cellule tumorali aumentando i livelli di ferritina, una proteina che lega il ferro.
Ma riducendo i livelli di ferritina, gli scienziati sono riusciti ad aumentare la tossicità della vitamina C per le cellule tumorali. In base a questa scoperta, gli scienziati hanno anche scoperto che i pazienti con carcinoma del colon-retto con alti livelli di proteine leganti il ferro hanno minori possibilità di sopravvivenza.
“In questo studio, abbiamo osservato come i cicli dietetici che imitano il digiuno sono in grado di aumentare l’effetto delle dosi farmacologiche di vitamina C e di ottenere tumori mutati dal KRAS”, ha affermato Maira Di Tano, co-autore dello studio presso l’IFOM, FIRC Institute of Oncologia molecolare a Milano, Italia.
“Ciò avviene attraverso la regolazione dei livelli di ferro e dei meccanismi molecolari coinvolti nello stress ossidativo. I risultati hanno indicato in particolare un gene che regola i livelli di ferro: eme-ossigenasi-1 “.
I precedenti studi del team di ricerca hanno dimostrato che il digiuno e una dieta che imitano il digiuno rallentano la progressione del cancro e rendono la chemioterapia più efficace nelle cellule tumorali, proteggendo le cellule normali dagli effetti collaterali associati alla chemioterapia.
La combinazione migliora la risposta antitumorale del sistema immunitario nei modelli di topo con carcinoma mammario e melanoma.
Gli scienziati ritengono che il cancro alla fine sarà trattato con farmaci a bassa tossicità in un modo simile a come gli antibiotici sono usati per trattare infezioni che uccidono determinati batteri, ma che possono essere sostituiti da altri farmaci se il primo non è efficace.
Per raggiungere questo obiettivo, affermano di dover prima verificare due ipotesi: che i loro interventi di combinazione non tossici funzionerebbero nei topi e che sembrerebbe promettente per gli studi clinici sull’uomo. In questo nuovo studio, hanno affermato di aver dimostrato entrambi.
Almeno cinque studi clinici, incluso uno presso l’USC su pazienti affetti da carcinoma mammario e prostatico, stanno attualmente studiando gli effetti delle diete che imitano il digiuno in combinazione con diversi farmaci antitumorali.
L’afta epizootica aumenta la tossicità della vitamina C nelle cellule tumorali mutanti del KRAS
Abbiamo studiato se il digiuno / FMD potenzia l’effetto anticancro della vitamina C contro diversi modelli di cancro mutante KRAS . Umano (HCT116, DLD-1) e murino (CT26) KRAS -mutant CRC linee cellulari, così come KRAS cancro ai polmoni -mutant (H23, H727) e le cellule adenocarcinoma del dotto pancreatico (PANC1), sono state coltivate in terreno di controllo (1 g / L di glucosio e siero al 10%; CTR) o in un mezzo simile all’afta epizootica (0,5 g / L di glucosio e 1% di siero), qui indicato come condizione di fame a breve termine (STS), che imita la riduzione del glucosio extracellulare e concentrazioni del fattore di crescita che si verificano durante il digiuno prolungato (> 48 h) o FMD in vivo, con o senza concentrazioni farmacologiche di vitamina C (≥0,3 mM).
Coerentemente con i recenti risultati 4 , le cellule tumorali mutanti del KRAS erano più sensibili alla vitamina C rispetto alle cellule tumorali di tipo KRAS di tipo selvaggio (Fig. 1a, b). Quando le cellule tumorali sono state coltivate in condizioni di STS prima e durante il trattamento, la tossicità mediata dalla vitamina C è stata fortemente potenziata (Fig. 1a).
Al contrario, le linee cellulari CRAS di tipo KRAS (SW48, HT29), carcinoma prostatico (PC-3), carcinoma ovarico (COV362) e una normale linea cellulare del colon (CCD841CoN) erano resistenti alla vitamina C se usate sia come agente singolo e in combinazione con STS (Fig. 1b).
In accordo con la tossicità selettiva della STS + vitamina C nelle cellule tumorali mutanti del KRAS , abbiamo scoperto che le cellule HT29 e SW48 geneticamente modificate per esprimere la forma attiva del KRAS erano più sensibili alla STS + vitamina C rispetto alla loro controparte isogena di tipo selvaggio (Fig. 1c e Supplementary Fig. 1).
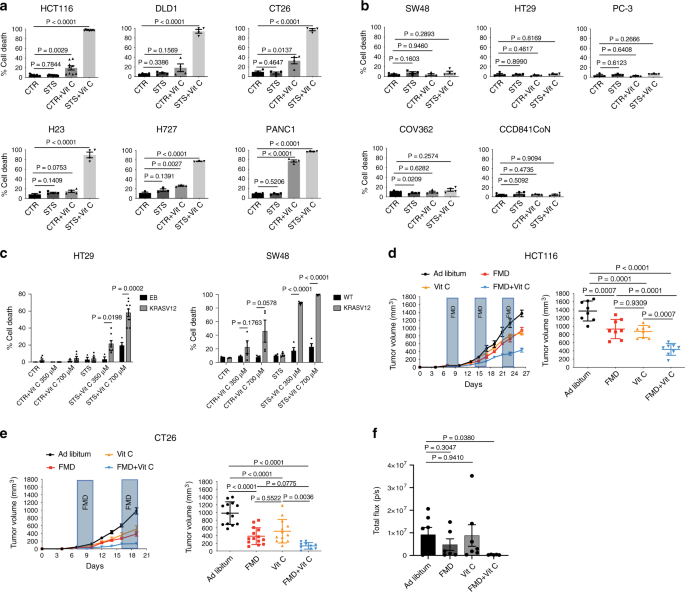
Coerentemente con i nostri risultati in vitro, abbiamo scoperto che i cicli dell’afta epizootica combinati con il trattamento giornaliero con vitamina C (4 g per kg due volte al giorno) erano efficaci nel ritardare la progressione dei tumori mutati KRAS in diversi modelli murini (Fig. 1d-f).
In particolare, i cicli settimanali di un FMD di tre giorni erano sufficienti per ridurre la crescita del tumore mutato KRAS nella stessa misura della vitamina C ad alto dosaggio (Fig. 1d, e). In particolare, l’afta epizootica settimanale e la vitamina C giornaliera hanno mostrato il miglior risultato terapeutico nel ridurre la progressione del CRC nei modelli di xenotrapianto e di topo sinergico, nonché in un modello ortotopico (Fig. 1d-f e Fig. 2a supplementare).
Inoltre, la combinazione FMD-vitamina C è stata sicura e ben tollerata in entrambi i ceppi di topo, come indicato dalla perdita di peso corporeo del topo, che non ha superato il 20% e che è stata rapidamente recuperata dopo il refeeding (Figura 2b supplementare).
ROS media la sensibilizzazione alla vitamina C.
In precedenza abbiamo dimostrato che il digiuno / FMD sensibilizza alla chemioterapia diversi tipi di cellule cancerose attraverso un meccanismo che comporta un aumento della produzione di ROS 17,25 . I ROS, inclusi H 2 O 2 e superossido, generati come sottoprodotti del normale metabolismo, causano danni a DNA, lipidi e proteine 26 .
Recenti studi hanno dimostrato che le mutazioni del KRAS promuovono la riprogrammazione metabolica per sostenere tassi di proliferazione elevati, accompagnati da uno stato ossidativo più elevato rispetto alle cellule di tipo selvatico KRAS 11,12,27,28 . Pertanto, abbiamo ipotizzato che lo stato ossidativo più elevato dei tumori mutanti del KRAS possa essere alla base del meccanismo selettivo di tossicità da FMD + vitamina C.
In particolare, la combinazione di STS e vitamina C ha fortemente indotto un danno al DNA nelle cellule CT26 e HCT116, come indicato dalla fosforilazione dell’istone H2AX (Fig. 2a), suggerendo che lo stress ossidativo può partecipare alla mediazione di questo effetto citotossico. Infatti, la combinazione di STS e vitamina C ha esacerbato selettivamente la produzione di ROS nelle cellule tumorali mutanti del KRAS (Fig. 2b e Fig. 3a supplementare).
Coerentemente con la produzione di H 2 O 2 dipendente dalla vitamina C , abbiamo osservato lo stesso aumento selettivo di ROS nelle cellule tumorali mutanti del KRAS cresciute in condizioni di STS e trattate con H 2 O 2 (Figura 3b supplementare).
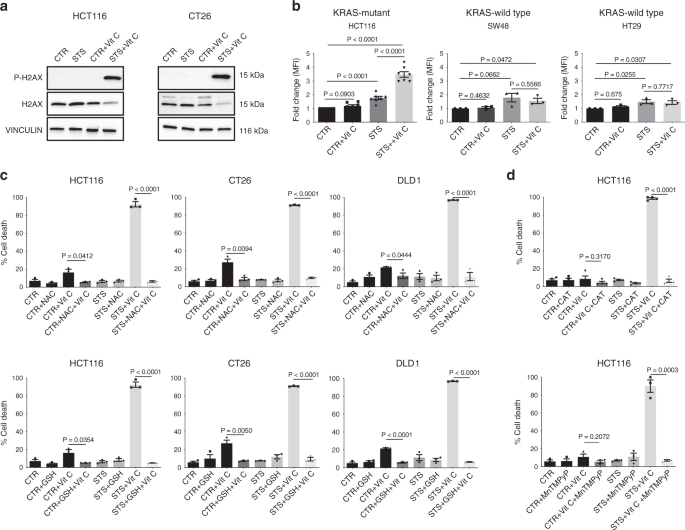
Per valutare direttamente se l’aumento della produzione di ROS è un evento causale o secondario nella sensibilizzazione indotta da STS alla vitamina C, abbiamo valutato l’effetto di diversi antiossidanti sulla tossicità di STS + vitamina C (Fig. 2c, d e Fig. 3c supplementare ).
Il glutatione (GSH) e la N-acetil cisteina (NAC), nonché la precedente esposizione alla catalasi impermeabile alla membrana (CAT) o alla superossido dismutasi permeabile alla membrana (SOD) / catalasi MnTMPyP sono riusciti a ripristinare la cellula indotta da STS + vitamina C morte (Fig. 2c, d ).
Collettivamente, questi risultati indicano che la produzione di ROS e le alterazioni redox rappresentano un meccanismo centrale attraverso il quale la combinazione STS e vitamina C uccide selettivamente le cellule tumorali mutate dal KRAS .
Il ferro è coinvolto nella tossicità mediata dall’afta epizootica
Numerose prove dimostrano che il meccanismo alla base degli effetti anticancro della vitamina C si basa sulla produzione di H 2 O 2 e che il LIP svolge un ruolo fondamentale in questo processo 3,6,7 .
In presenza di ferro libero, alti livelli di H 2 O 2 hanno effetti pro-ossidanti in parte attraverso la generazione di radicali idrossilici attraverso la reazione di Fenton e l’induzione del danno ossidativo 3,7 .
Poiché la combinazione di FMD / STS e vitamina C ha aumentato i livelli di ROS nelle cellule tumorali mutanti del KRAS , abbiamo studiato se ciò è correlato a un aumento del pool di ferro ferroso labile. Nelle cellule HCT116, i livelli di ioni ferrosi (Fe 2+ ) sono stati significativamente aumentati con STS e co-trattamento con vitamina C rispetto a tutte le altre condizioni (Fig. 3a), suggerendo un potenziale ruolo del ferro nella mediazione di questo effetto.
La ferritina, la principale proteina coinvolta nel legame e nella conservazione del ferro, regola il LIP intracellulare e la sua downregulation ha dimostrato di aumentare il LIP nelle cellule tumorali mutanti KRAS 10,11,12 .
Pertanto, abbiamo misurato i livelli della subunità pesante della ferritina (FTH), che è responsabile dello stoccaggio del ferro attraverso la sua attività ferroxidasi 10 . Coerentemente con l’aumento del ferro ferroso, abbiamo scoperto che STS, da solo o in combinazione con vitamina C, ha regolato in modo selettivo l’espressione della proteina FTH nelle cellule tumorali mutanti del KRAS . D’altra parte, la vitamina C ha invertito la downregulation della ferritina indotta da STS nelle cellule tumorali di tipo KRAS di tipo selvaggio (Fig. 3b, Fig. 4 supplementare).
Questi risultati in vitro sono stati confermati anche in vivo, in cui i cicli dell’afta epizootica combinati con il trattamento con vitamina C hanno ridotto l’espressione della proteina FTH nei tumori derivati da HCT116 (Fig. 3c).
Fig. 3: Il ferro è coinvolto selettivamente nella tossicità dell’afta epizootica + vitamina C nelle cellule tumorali mutate dal KRAS .
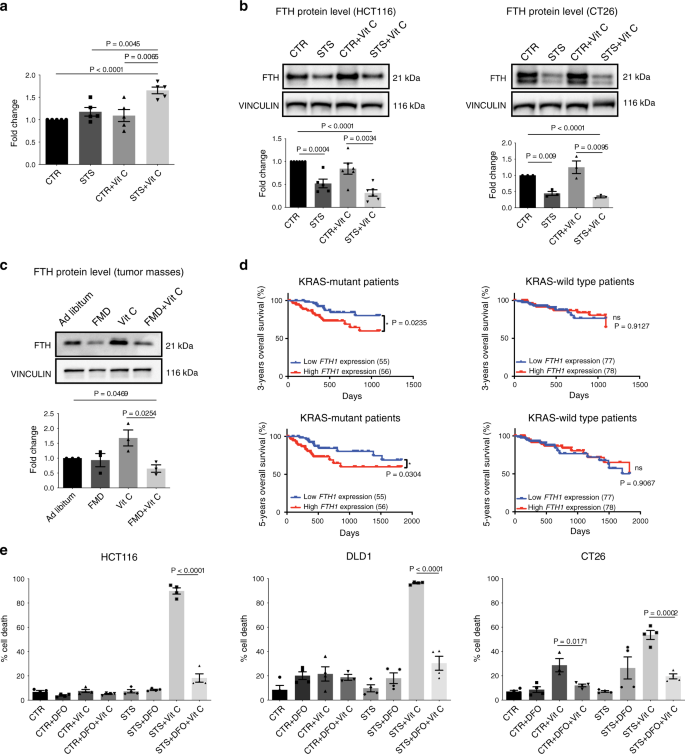
Coerentemente con un effetto sensibilizzante della downregulation dell’FTH nelle cellule tumorali mutanti KRAS , la nostra analisi dei dati di sopravvivenza del paziente CRC acquisiti dal Cancer Genome Atlas Database ( TCGA ), ha mostrato che i pazienti con tumori KRAS mutati e basso livello trascrizionale di ferritina intratumorale avevano un tempo più lungo 3 – e sopravvivenza globale a 5 anni se confrontata con pazienti i cui tumori esprimevano un alto livello di ferritina (Fig. 3d ). Questa associazione, che non è stata osservata per i tumori KRAS di tipo selvaggio, supporta il ruolo dell’afta epizootica + vitamina C come strategia per mantenere bassi livelli di ferritina e aumentare il ROS per il trattamento del KRAS tumori mutati. In particolare, questi dati rappresentano meno di 200 pazienti; pertanto, sono necessarie analisi di popolazioni di pazienti più grandi per comprendere meglio il ruolo dell’espressione o dell’attività della ferritina nella progressione dei tumori con mutazione KRAS .
Per valutare se l’alterazione del contenuto di ferro cellulare contribuisce alla sensibilizzazione STS-mediata alla vitamina C, le cellule CRC mutanti KRAS coltivate in condizioni di STS sono state trattate con il ferro chelatore desferrioxamina (DFO) prima dell’esposizione alla vitamina C. Coerentemente con la nostra ipotesi, il trattamento con DFO che precede l’esposizione alla vitamina C ha salvato la citotossicità cellulare indotta dalla vitamina C (Fig. 3e ), confermando così che l’aumento del ferro libero intracellulare mediato da FMD / STS e la vitamina C è, almeno in parte, responsabile di il loro effetto citotossico.
L’afta epizootica inverte l’effetto della vitamina C su HO-1
Numerosi studi hanno suggerito un potenziale ruolo della ferritina nella protezione delle cellule dal danno ossidativo attraverso il sequestro del ferro libero intracellulare 10,11,12 . Tra gli enzimi che promuovono l’espressione della ferritina, HO-1 inducibile dallo stress è stato implicato nel promuovere la sopravvivenza cellulare durante l’esposizione cellulare agli insulti ossidativi 29,30 .
Poiché l’FMD / STS sottoregola il livello di espressione proteica dell’FTH, abbiamo studiato se HO-1 è implicato nella regolazione dell’FTH in risposta al trattamento con STS e vitamina C. In uno studio recente del nostro gruppo, l’afta epizootica ha sensibilizzato le cellule tumorali al seno alla chemioterapia in parte ridimensionando HO-1, supportando ulteriormente un possibile ruolo di questa proteina inducibile dallo stress nel mediare gli effetti benefici dell’afta epizootica 31 .
Per verificare la nostra ipotesi, abbiamo valutato l’associazione tra espressione HO-1 e induzione FTH. A tal fine, abbiamo trattato le cellule HCT116 con l’emino attivatore HO-1 e verificato che l’emina aumenta i livelli di proteina HO-1 e FTH in condizioni di crescita CTR e STS (Figura 5 aggiuntiva).
Inoltre, abbiamo scoperto che il trattamento con vitamina C ha significativamente sovraregolato HO-1, mentre l’FMD / STS ha invertito questo effetto sia in vitro che in vivo nelle cellule tumorali mutanti del KRAS (Fig. 4a e Fig. 6a supplementare).
Nelle cellule tumorali di tipo KRAS di tipo selvatico i livelli di HO-1 non sono stati alterati in seguito alla somministrazione di vitamina C in condizioni di CTR e la combinazione di vitamina C e STS non ha downregolato ma ha indotto invece il livello di espressione proteica di HO-1 (Fig. 4b, Figura supplementare 6b).
Fig. 4: La modulazione HO-1 e la transferrina legata al ferro sono i protagonisti della sensibilizzazione FMD-dipendente alla vitamina C.
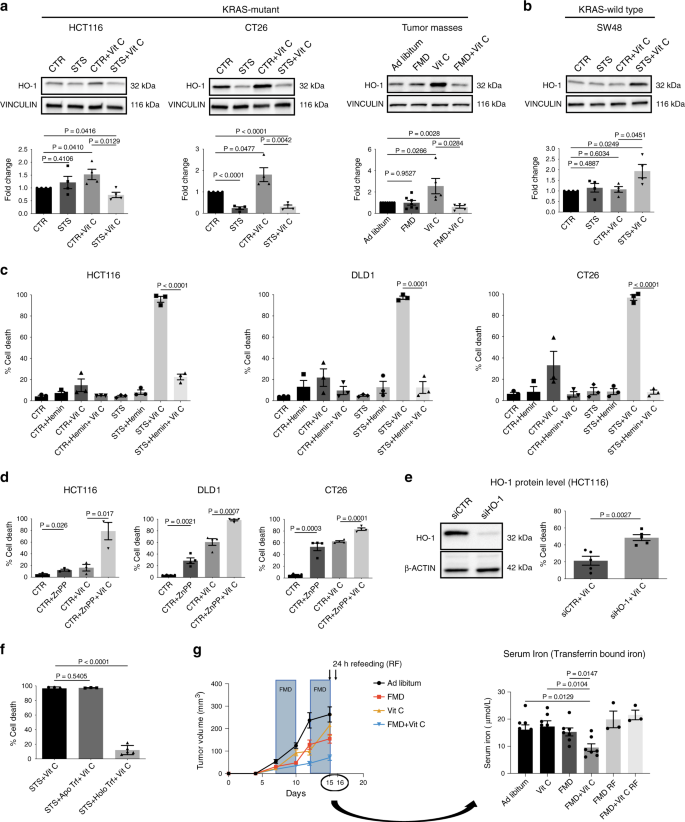
Collettivamente, questi risultati indicano che la regolazione differenziale di HO-1 e l’effetto risultante sulla via ferritina / ferro mediano selettivamente la sensibilizzazione FMD-dipendente alla vitamina C in uno sfondo mutante di KRAS .
La capacità dell’afta epizootica di prevenire l’ upregolazione di HO-1 indotta dalla vitamina C nelle cellule tumorali mutanti del KRAS ci ha spinto a studiare se i livelli di HO-1 influenzano la sensibilità delle cellule tumorali al trattamento combinato. Coerentemente con questa ipotesi, l’attivatore HO-1 ha protetto le cellule CRC mutanti KRAS umane e murine dalla morte cellulare indotta dalla vitamina C, da sole o in combinazione con STS (Fig. 4c).
D’altra parte, l’inibitore di HO-1 protoporfirina di zinco (ZnPP) ha reso le cellule CRC mutanti del KRAS più sensibili alla vitamina C in condizioni ricche di nutrienti (CTR) (Fig. 4d). Coerentemente con questi risultati, il knockdown HO-1 nelle cellule HCT116 ha anche aumentato la morte delle cellule tumorali in seguito all’esposizione alla vitamina C in condizioni CTR (Fig. 4e). Pertanto, nel complesso questi risultati supportano il ruolo di HO-1 nella regolazione della sensibilità delle cellule tumorali mutanti del KRAS alla vitamina C.
Per caratterizzare come FMD / STS influenza i livelli di HO-1 e modulare la sensibilità delle cellule tumorali alla vitamina C, abbiamo analizzato l’effetto della glicemia o della deprivazione sierica. È interessante notare che abbiamo scoperto che il glucosio, i fattori di crescita sierica e gli amminoacidi non erano responsabili del miglioramento della tossicità da vitamina C dipendente dalla STS (Figura 7a supplementare).
L’aggiunta di holo-transferrina (forma legata al ferro) ma non apo-transferrina (forma priva di ferro) ha invertito la tossicità mediata da STS + vitamina C e la downregulation dell’asse HO-1 / FTH (Fig. 4f; Fig. 7b supplementare). Questi risultati sono coerenti con il concetto che i livelli di ferro nel siero sono importanti nel mediare l’effetto STS.
In effetti, in accordo con i nostri risultati in vitro, FMD + vitamina C in vivo ha ridotto i livelli ematici di ferro legato alla transferrina (Fig. 4g). I nostri dati supportano ulteriormente il ruolo del ferro come fattore chiave nel siero la cui riduzione è stata responsabile del sinergismo di afta epizootica e vitamina C.
L’afta epizootica e la vitamina C potenziano gli effetti citotossici dell’oxaliplatino
Numerosi studi hanno descritto la tollerabilità e la potenziale efficacia della vitamina C ad alte dosi come trattamento adiuvante durante la chemioterapia 2,3,32,33 . Inoltre, il nostro gruppo ha recentemente dimostrato l’efficacia dei cicli di digiuno o FMD in combinazione con la chemioterapia per ridurre la crescita tumorale in una vasta gamma di tipi di cancro rispetto alla sola chemioterapia standard 17,31 .
Attingendo ai nostri dati precedenti, abbiamo studiato se FMD + vitamina C avrebbe sensibilizzato le cellule tumorali mutate dal KRAS all’azione pro-ossidante della chemioterapia in vivo, possibilmente aumentando lo stress ossidativo cellulare.
Abbiamo scelto l’oxaliplatino perché è uno dei composti citotossici più efficaci utilizzati nell’adiuvante e nell’impostazione avanzata del trattamento CRC 34 . Da notare che la combinazione di FMD + vitamina C era efficace quanto oxaliplatino + FMD o oxaliplatino + vitamina C, supportando la potente azione di queste combinazioni non tossiche nell’arrestare la crescita del tumore (Fig. 5a).
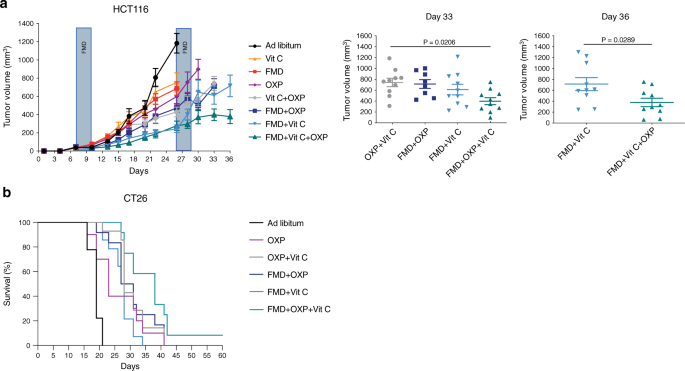
Topi NSG e BALB / c sono stati iniettati per via sottocutanea con cellule HCT116 e cellule CT26, rispettivamente. I topi sono stati nutriti ad libitum o sottoposti a cicli di afta epizootica e trattati con o senza vitamina C o oxaliplatino (10 mg / kg). una progressione del tumore HCT116 (a sinistra) e volume al giorno 33 e 36 (a destra), rispettivamente ( n = 10 in Ad libitum, FMD, FMD + Vit C, Vit C + OXP, FMD + Vit C + OXP, n = 8 in FMD + OXP, n = 9 in OXP, n = 11 in Vit C). I valori di P sono stati determinati da ANOVA a una via con analisi post di Tukey (giorno 33) e t-test (giorno 36). I dati sono rappresentati come media ± SEM. b BALB / c con curve di sopravvivenza CT26 ( n = 9 in Ad libitum, n = 10 in OXP, n = 12 in FMD + OXP e FMD + OXP + Vit C, n = 14 in OXP + Vit C e FMD + Vit C ). I valori di P sono stati determinati dal test Log-rank (Mantel-Cox) (Ad libitum vs OXP, p = 0,0040; OXP vs FMD + Vit C, p = 0.7177; OXP vs FMD + OXP + VitC, p = 0,0114; FMD + OXP vs FMD + OXP + Vit C, p = 0,0488; OXP + Vit C vs FMD + OXP + Vit C, p = 0,0345; FMD + OXP + Vit C vs FMD + Vit C, p = 0.0003).
Inoltre, il triplo trattamento (FMD + vitamina C + chemioterapia) è stato l’intervento terapeutico più attivo nel ritardare la progressione del tumore in uno xenotrapianto di topo e nell’estendere la sopravvivenza in un modello sinergico (Fig. 5a, be Fig. 8 Supplementare).
Questi risultati indicano che la chemioterapia può potenziare ulteriormente gli effetti dell’afta epizootica + vitamina C contro i tumori mutati KRAS .
More information: Maira Di Tano et al, Synergistic effect of fasting-mimicking diet and vitamin C against KRAS mutated cancers, Nature Communications (2020). DOI: 10.1038/s41467-020-16243-3
References
1. Hoffer, L. J. et al. Phase I clinical trial of i.v. ascorbic acid in advanced malignancy. Ann. Oncol. 19, 1969–1974 (2008).
2.Ma, Y. et al. High-dose parenteral ascorbate enhanced chemosensitivity of ovarian cancer and reduced toxicity of chemotherapy. Sci. Transl. Med. 6, 222ra18 (2014).
3. Schoenfeld, J. D. et al. O2⋅- and H2O2-mediated disruption of Fe metabolism causes the differential susceptibility of NSCLC and GBM cancer cells to pharmacological ascorbate. Cancer Cell 32, 268 (2017).
4. Yun, J. et al. Vitamin C selectively kills KRAS and BRAF mutant colorectal cancer cells by targeting GAPDH. Science 350, 1391–1396 (2015).
5.Aguilera, O. et al. Vitamin C uncouples the Warburg metabolic switch in KRAS mutant colon cancer. Oncotarget 7, 47954–47965 (2016).
6. Chen, Q. et al. Ascorbate in pharmacologic concentrations selectively generates ascorbate radical and hydrogen peroxide in extracellular fluid in vivo. Proc. Natl Acad. Sci. USA 104, 8749–8754 (2007).
7.Chen, Q. et al. Pharmacologic doses of ascorbate act as a prooxidant and decrease growth of aggressive tumor xenografts in mice. Proc. Natl Acad. Sci. USA 105, 11105–11109 (2008).
8.Du, J. et al. Mechanisms of ascorbate-induced cytotoxicity in pancreatic cancer. Clin. Cancer Res. 16, 509–520 (2010).
9.Moser, J. C. et al. Pharmacological ascorbate and ionizing radiation (IR) increase labile iron in pancreatic cancer. Redox Biol. 2, 22–27 (2013).
10.Torti, S. V. & Torti, F. M. Iron and cancer: more ore to be mined. Nat. Rev. Cancer 13, 342–355 (2013).
11.Kakhlon, O., Gruenbaum, Y. & Cabantchik, Z. I. Ferritin expression modulates cell cycle dynamics and cell responsiveness to H-ras-induced growth via expansion of the labile iron pool. Biochem. J. 363, 431–436 (2002).
12.Yang, W. S. & Stockwell, B. R. Synthetic lethal screening identifies compounds activating iron-dependent, nonapoptotic cell death in oncogenic-RAS-harboring cancer cells. Chem. Biol. 15, 234–245 (2008).
13.Ibrahim, W. H., Habib, H. M., Kamal, H., Clair, D. K. S. & Chow, C. K. Mitochondrial superoxide mediates labile iron level: evidence from Mn-SOD-transgenic mice and heterozygous knockout mice and isolated rat liver mitochondria. Free Radic. Biol. Med. 65, 143–149 (2013).
14.Caltagirone, A., Weiss, G. & Pantopoulos, K. Modulation of cellular iron metabolism by hydrogen peroxide. Effects of H2O2 on the expression and function of iron-responsive element-containing mRNAs in B6 fibroblasts. J. Biol. Chem. 276, 19738–19745 (2001).
15.Otterbein, L. E., Soares, M. P., Yamashita, K. & Bach, F. H. Heme oxygenase-1: unleashing the protective properties of heme. Trends Immunol. 24, 449–455 (2003).
16.Stephen, A. G., Esposito, D., Bagni, R. K. & McCormick, F. Dragging ras back in the ring. Cancer Cell 25, 272–281 (2014).
17.Lee, C. et al. Fasting cycles retard growth of tumors and sensitize a range of cancer cell types to chemotherapy. Sci. Transl. Med 4, 124ra27 (2012).
18.Raffaghello, L. et al. Starvation-dependent differential stress resistance protects normal but not cancer cells against high-dose chemotherapy. Proc. Natl Acad. Sci. USA 105, 8215–8220 (2008).
19.Lee, C. et al. Reduced levels of IGF-I mediate differential protection of normal and cancer cells in response to fasting and improve chemotherapeutic index. Cancer Res. 70, 1564–1572 (2010).
20.Lee, C. & Longo, V. D. Fasting vs dietary restriction in cellular protection and cancer treatment: from model organisms to patients. Oncogene 30, 3305–3316 (2011).
21.Longo, V. D. & Mattson, M. P. Fasting: molecular mechanisms and clinical applications. Cell Metab. 19, 181–192 (2014).
22.Di Biase, S. et al. Fasting regulates EGR1 and protects from glucose- and dexamethasone-dependent sensitization to chemotherapy. PLoS Biol. 15, e1002603 (2017).
23.Brandhorst, S. et al. A periodic diet that mimics fasting promotes multi-system regeneration, enhanced cognitive performance, and healthspan. Cell Metab. 22, 86–99 (2015).
24.Wei, M. et al. Fasting-mimicking diet and markers/risk factors for aging, diabetes. Cancer, Cardiovascular Dis. Sci. Transl. Med. 15, 9 (2017).
25.Bianchi, G. et al. Fasting induces anti-Warburg effect that increases respiration but reduces ATP-synthesis to promote apoptosis in colon cancer models. Oncotarget 6, 11806–11819 (2015).
26.Reczek, C. R. & Chandel, N. S. The two faces of reactive oxygen species in cancer. Annu. Rev. Cancer Biol. 1, 79–98 (2017).
27.Weinberg, F. et al. Mitochondrial metabolism and ROS generation are essential for Kras-mediated tumorigenicity. Proc. Natl Acad. Sci. USA 107, 8788–8793 (2010).
28.Ogrunc, M. et al. Oncogene-induced reactive oxygen species fuel hyperproliferation and DNA damage response activation. Cell Death Differ. 21, 998–112 (2014).
29.Ferris, C. D. et al. Haem oxygenase-1 prevents cell death by regulating cellular iron. Nat. Cell Biol. 1, 152–157 (1999).
30.Gonzales, S., Erario, M. A. & Tomaro, M. L. Heme oxygenase-1 induction and dependent increase in ferritin. Dev. Neurosci. 24, 161–168 (2002).
31.Di Biase, S. et al. Fasting-mimicking diet reduces HO-1 to promote T cell-mediated tumor cytotoxicity. Cancer Cell 30, 136–146 (2016).
32.Monti, D. A. et al. Phase I evaluation of intravenous ascorbic acid in combination with gemcitabine and erlotinib in patients with metastatic pancreatic cancer. PLoS ONE 7, e29794 (2012).
33.Welsh, J. L. et al. Pharmacological ascorbate with gemcitabine for the control of metastatic and node-positive pancreatic cancer (PACMAN): results from a phase I clinical trial. Cancer Chemother. Pharmacol. 71, 765–775 (2013).
34.Alcindor, T. & Beauger, N. Oxaliplatin: a review in the era of molecularly targeted therapy. Curr. Oncol. 18, 18–25 (2011).
35.Muliaditan, T. et al. Repurposing tin mesoporphyrin as an immune checkpoint inhibitor shows therapeutic efficacy in preclinical models of cancer. Clin. Cancer Res. 24, 1617–1628 (2018).
36.Was, H., Dulak, J. & Jozkowicz, A. Heme oxygenase-1 in tumor biology and therapy. Curr. Drug Targets 11, 1551–1570 (2010).
37.Matsuo, T. et al. Pathological significance and prognostic implications of heme oxygenase 1 expression in non-muscle-invasive bladder cancer: Correlation with cell proliferation, angiogenesis, lymphangiogenesis and expression of VEGFs and COX-2. Oncol. Lett. 13, 275–280 (2017).
38.Deng, Y. et al. The Nrf2/HO-1 axis can be a prognostic factor in clear cell renal cell carcinoma. Cancer Manag. Res. 7, 1221–1230 (2019).
39.Sun, W. et al. TSVdb: a web-tool for TCGA splicing variants analysis. BMC Genomics. 19, 405 (2018).



{kind=link}