









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A joint research team led by Sunney Xie, Director of Beijing Advanced Innovation Center for Genomics (ICG) at Peking University (PKU) has successfully identified multiple highly potent neutralizing antibodies against the novel coronavirus SARS-CoV-2, the causative virus of the respiratory disease COVID-19, from convalescent plasma by high-throughput single-cell sequencing.
Generated by human immune system, neutralizing antibodies can effectively prevent viruses from infecting cells.
New results from animal studies showed that their neutralizing antibody provides a potential cure for COVID-19 as well as means for short-term prevention.
Sunney Xie, director of the university’s Beijing Advanced Innovation Center for Genomics, told AFP that the drug has been successful at the animal testing stage.
“When we injected neutralising antibodies into infected mice, after five days the viral load was reduced by a factor of 2,500,” said Xie.
“That means this potential drug has (a) therapeutic effect.”
The drug uses neutralising antibodies – produced by the human immune system to prevent the virus infecting cells – which Xie’s team isolated from the blood of 60 recovered patients.
A study on the team’s research, published Sunday in the scientific journal Cell, suggests that using the antibodies provides a potential “cure” for the disease and shortens recovery time.
Xie said his team had been working “day and night” searching for the antibody.
“Our expertise is single-cell genomics rather than immunology or virology. When we realised that the single-cell genomic approach can effectively find the neutralising antibody we were thrilled.”
He said he hopes that the drug will be ready for use later this year and in time for any potential winter outbreak of the virus, which has infected 4.8 million people around the world and killed more than 315,000.
“Planning for the clinical trial is underway,” said Xie, adding it will be carried out in Australia and other countries since cases have dwindled in China, offering fewer human guinea pigs for testing.
“The hope is these neutralising antibodies can become a specialised drug that would stop the pandemic,” he said.
China already has five potential coronavirus vaccines at the human trial stage, a health official said last week.
But the World Health Organization has warned that developing a vaccine could take 12 to 18 months.
This study has been published online in Cell, titled “Potent neutralizing antibodies against SARS-CoV-2 identified by high-throughput single-cell sequencing of convalescent patients’ B cells”.
This work was jointly conducted by Beijing Advanced Innovation Center for Genomics and Biomedical Pioneering Innovation Center of Peking University, Beijing YouAn Hospital of Capital Medical University, Institute of Laboratory Animal Science(ILAS) of Chinese Academy of Medical Sciences and Comparative Medicine Center, Peking Union Medical College, Beijing Institute of Microbiology and Epidemiology of Academy of Military Medical Sciences, Sino Biological, Inc., WuXi Biologics and Singlomics.

The co-authors of this article are Yunlong Cao, Bin Su, Xianghua Guo, Wenjie Sun, Yongqiang Deng, Linlin Bao, Qinyu Zhu. The corresponding authors are Chuan Qin, Chengfeng Qin, Ronghua Jin, and Sunney Xie.
The work which began on Jan. 27, 2020 was supported by The People’s Government of Beijing Municipality, the Ministry of Science and Technology and the Ministry of Education of the People’s Republic of China.
This marks a major milestone in the fight against the pandemic.
Coronavirus Disease 2019 (COVID-19) caused by a novel coronavirus named Severe Acute Respiratory Syndrome coronavirus 2 (SARS-CoV-2) has spread globally as a severe pandemic (Callaway et al., 2020). Both SARS-CoV-2 and SARS-CoV belong to lineage B of the betacoronavirus genus (Zhou et al., 2020; Wu et al., 2020), and their RNA genomes share around 82% identity (Chan et al., 2020).
The mechanisms by which SARS-CoV-2 infects target cells have been well studied and recently reported (Hoffmann et al., 2020; Walls et al., 2020). Similar to SARS-CoV, the spike (S) glycoprotein on the surface of SARS-CoV-2 mediates membrane fusion and receptor recognition of the virus (Wrapp et al., 2020).
The S1 subunit at the N-terminal region is responsible for virus attachment and contains the receptor-binding domain (RBD), which directly binds to the ACE2 receptor on the host cell. Currently, no validated therapeutics against virus-target interactions are available for COVID-19.
Convalescent patients’ plasma, which contains neutralizing antibodies produced by the adaptive immune response, has led to a clear clinical improvement of both mild and severe COVID-19 patients when used as a therapeutic modality (Chen et al., 2020; Shen et al., 2020; Cao, 2020).
However, therapeutic use is limited since plasma can not be produced on a large-scale. On the other hand, neutralizing monoclonal antibodies (mAbs) isolated from convalescent patient’s memory B cells may serve as a promising intervention to SARS-CoV-2 due to their scalability and therapeutic effectiveness.
Human-sourced mAbs targeting viral surface proteins have increasingly shown their therapeutic and prophylactic efficacy against infectious diseases such as HIV, Ebola, and MERS (Corti et al., 2016; Wang et al., 2018; Scheid et al., 2009).
Their safety and potency in patients have been demonstrated in multiple clinical trials (Xu et al., 2019; Caskey et al., 2017). Despite their advantages, screening for potent neutralizing mAbs from human memory B cells is often a slow and laborious process, which is not ideal when responding to a global health emergency. A rapid and efficient method for screening SARS-CoV-2 neutralizing mAbs is urgently needed.
Due to VDJ recombination and somatic hypermutation, B cells exhibit diverse B-cell repertoires, necessitating the analysis of one B cell at a time (Bassing et al., 2002).
Techniques, such as single-cell clonal amplification of memory B cells, are usually utilized to obtain paired immunoglobulin heavy-light chain RNA sequences from the heterogeneous B cell population to produce mAbs (Debs et al., 2012; Niu et al., 2019).
Clonal amplification of (Epstein-Barr virus) EBV-immortalised memory B cells from convalescent patients has proved successful in isolating neutralizing mAbs against viral infections such as HIV, Dengue, and MERS (Scheid et al., 2009; Burton et al., 2009; Corti et al., 2015).
Yet, due to the time-consuming incubation and screening steps, the technique takes several months at least to complete a successful screen.
On the other hand, single-cell RT-PCR combined with fluorescence-activated cell sorting (FACS) or optofluidics platform, such as Beacon (Berkley Light), could obtain antibody sequences in several days by performing nested PCR on single antigen-binding memory B cells after single-cell sorting (Tiller et al., 2008; Wardemann et al., 2003; Wrammert et al., 2008; Liao et al., 2009).
The method has led to efficient isolation of neutralizing mAbs in various infectious diseases, including HIV and MERS (Scheid et al., 2009; Wang et al., 2018). Nevertheless, the recent development of high-throughput single-cell RNA and VDJ sequencing of B-cell receptor repertoires using 10X Chromium has outperformed single-cell RT-PCR in terms of B cell screening throughput (Goldstein et al., 2019; Horns et al., 2020).
The microfluidic-based technique could obtain auto-paired heavy- and light-chain sequences from tens of thousands single B cells in one run and has successfully been used for the isolation of human neutralizing mAbs against HIV (Setliff et al., 2019).
Here we report the rapid and efficient identification of SARS-CoV-2 neutralizing antibodies achieved by high-throughput single-cell RNA and VDJ sequencing of antigen-binding B cells from convalescent COVID-19 patients.
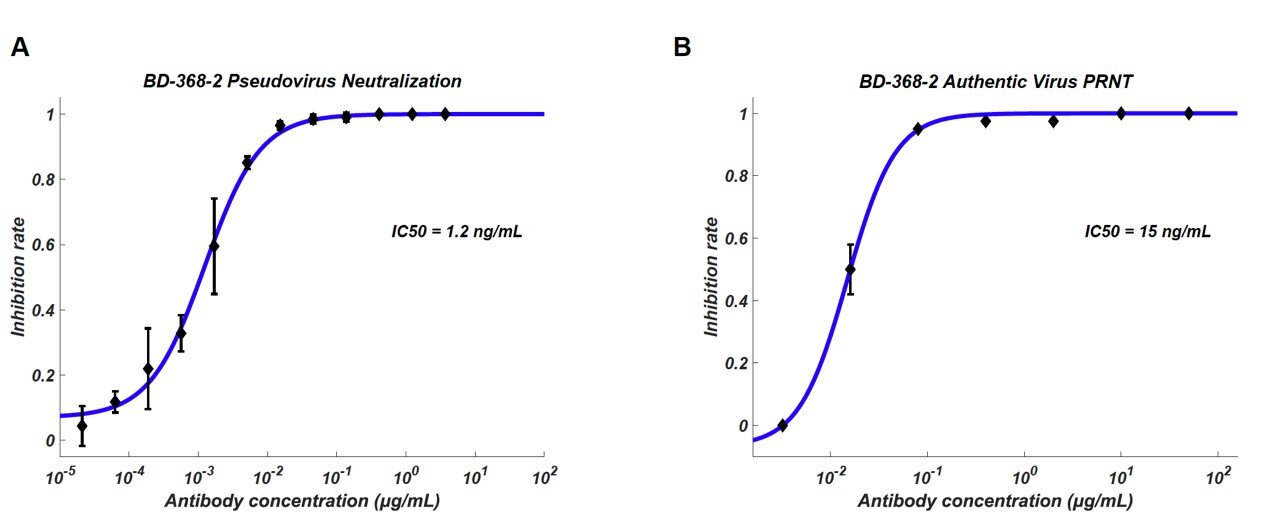
Over 8,500 antigen-binding B cell clonotypes expressing IgG1 antibodies were identified from 60 convalescent patients. In total, we identified 14 potent neutralizing mAbs, among which the most potent mAb, BD-368-2, exhibited an IC50 of 1.2 ng/mL and 15 ng/mL against pseudotyped and authentic SARS-CoV-2.
Additionally, in vivo experiments confirmed that BD-368-2 could provide strong therapeutic efficacy and prophylactic protection against SARS-CoV-2, using the hACE2 transgenic mice model (Bao et al., 2020; Yang et al., 2007; McCray et al., 2007).
Further, we solved the Cryo-EM structure of one neutralizing mAb, BD-23, in complex with the SARS-CoV-2 spike ectodomain trimer and showed that its epitope overlaps with the RBD/ACE2 binding motif.
Moreover, based on the high conservation between SARS-CoV-2 and SARS-CoV, we demonstrated that potent neutralizing mAbs against SARS-CoV-2 could be directly chosen from the large antigen-binding clonotype library, utilizing the CDR3H structure’s similarity to that of SARS-CoV neutralizing antibody m396 (Prabakaran et al., 2006; Zhu et al., 2007).
Overall, we showed that high-throughput single-cell sequencing could lead to the identification of highly potent neutralizing mAbs that have strong therapeutic and prophylactic efficacy, which could greatly assist in the intervention of prevailing and emerging infectious diseases, such as COVID-19.
Results
High-throughput sequencing of single B cells from convalescent patients
Unlike traditional methodologies, large-scale data obtained from high-throughput scVDJ-seq enabled us to examine B cell clonotype enrichment prior to in vitro antibody expression (Goldstein et al., 2019; Croote et al., 2018).
B cells that share an identical CDR3 region for both heavy- and light chains were grouped into the same clonotypes, and their enrichment was calculated based on the number of cells observed for the clonotype.
Since antigen-activated B cells would go through clonal selection and expansion from pre-existing naive and memory B cells (Murugan et al., 2018; Seifert and Kuppers, 2016), we hypothesized that enriched B cell clonotypes would more likely yield high-affinity SARS-CoV-2 binding and neutralizing antibodies.
To exploit this hypothesis, we first collected peripheral blood mononuclear cells (PBMCs) and isolated the B cells from 12 COVID-19 convalescent patients from Beijing Youan Hospital (Table S1). We performed scRNA and scVDJ sequencing on the freshly isolated B cells or CD27+ memory B cell subsets using 10X Chromium 5’ mRNA and VDJ sequencing
(Figure 1A and S1A). The scVDJ data indeed revealed enriched IgG1 clonotypes (Figure S1F and S1H), and the CD27+ memory B cell selection largely improved the number of memory B cells sequenced (Figure S1D and S1E) as well as the IgG1 clonotypes discovered (Figure S1G and S1I).
However, from the 130 in vitro expressed mAbs selected from memory B cells containing enriched IgG1+ clonotypes (Table S2), only one out of the two RBD-binding mAbs, named BD-23, showed a weak virus neutralization ability against pseudotyped and authentic SARS-CoV-2 (Figure S1B and S1C).
The low efficiency in terms of RBD-binding mAbs and neutralizing mAbs identification necessitates an antigen-affinity based selection, which could greatly enrich for spike/RBD binding B cells.
Efficient identification of neutralizing mAbs by high-throughput sequencing of antigen-binding single B cells
To enrich for RBD-binding B cells, we utilized both biotinylated RBD and S protein to select for antigen-binding B cells through magnetic beads separation (Figure 1A). Since the selection introduces a large reduction in the number of B cells, we combined the PBMCs from different patients for sufficient 10X loading.
In total, we analyzed 60 convalescent patients’ antigen-binding B cells in 6 different batches (Table S1).
Since the goal was to produce neutralizing mAbs, we filtered out all B cells that failed to have productive V-J spanning heavy-light chain pairs.
A total of 8,558 distinct IgG1-presenting antigen-binding clonotypes were detected (Table 1). Furthermore, the scRNA-seq data enabled us to perform cell-typing based on mRNA expression for memory B cell identifications (Figure 1B and S2).
We observed sufficient separation between naive B cells and memory B cells, as well as non-B cells versus memory B cells. However, a clear separation between switched/non-switched memory B cells and exhausted memory B cells may not be observed in some batches, mostly due to the relatively low sequencing depth and the low number of genes detected per cell of the scRNA sequencing (Table S3).
To increase the efficiency of neutralizing mAbs identification, we developed a set of criteria to filter out the clonotypes that have low chances to produce neutralizing mAbs (Figure 1D).
First, only enriched clonotypes containing IgG1-expressing B cells were selected (Figure 1C and S3A), since IgG1-expressing B cells respond strongly to viral stimuli (Vidarsson et al., 2014).
Secondly, IgG2-expressing B cells normally respond weakly to viral pathogens, and thus, IgG2-presenting clonotypes were not included as ideal candidates (Irani et al., 2015).
Next, clonotypes that did not contain any B cells with a somatic hypermutation rate (SHM) higher than 2% indicate insufficient affinity maturation and were excluded (Croote et al., 2018; Methor et al., 2017).
Lastly, as exhausted B cells and naive B cells respond lesseffective to antigen stimuli (Moir and Fauci, 2014), clonotypes that contained only exhausted memory B cells or naive B cells were not considered as ideal candidates (Figure 1B and S2).
Together, a total of 169 ideal candidates were selected from enriched clonotypes and were expressed in HEK293 cells through transfection (Table S2). Meanwhile, 47 non-ideal candidates were also expressed in vitro as a reference to validate the effectiveness of our selection criteria (Table S2).
The purified mAbs were tested for SARS-CoV-2 RBD/spike reactivity by ELISA and surface plasmon resonance (SPR), and 149 S-binding mAbs were identified, among which over half of the mAbs bind to the RBD.
We further screened all ELISA-positive mAbs for neutralizing ability using a SARS-CoV-2 pseudovirus system. We found that only RBD-binding mAbs showed pseudovirus neutralization effects (Figure 1E), and only mAbs bound to the RBD with a Kd smaller or close to the dissociation constant of ACE2/RBD (15.9 nM) (Figure S5B), would have significant neutralization effects (IC50<3 µg/mL) on SARS-CoV-2 pseudovirus (Figure S4A and S4B).
Both full-length S protein and the RBD protein could enrich for RBD-binding mAbs, but with distinct efficiency (Figure 1F).
Using the RBD protein as the enrichment antigen resulted in an RBD ELISA-positive rate three-fold higher than using S protein. Most importantly, mAbs selected from ideal candidates showed an efficiency of 46% and 25% in identifying strong RBD-binding mAbs (Kd<20 nM) and SARS-CoV-2 neutralizing mAbs, respectively (Figure S3D), which is significantly higher compared to the non-ideal mAbs candidates (Figure S3B and S3C).
High neutralizing potency against SARS-CoV-2 among mAbs from enriched clonotypes
Among all the neutralizing mAbs isolated from enriched clonotypes, seven of them showed potent neutralization ability with an IC50 lower than 0.05 µg/mL against SARS-CoV-2 pseudovirus (Figure 2A and 2C).
The most potent mAb BD-368-2, which was selected from the same clonotype as BD-368 (Figure 1C), exhibited an IC50 of 1.2 ng/mL. All seven mAbs bind strongly to the RBD with nM or sub-nM Kd, revealed by surface plasmon resonance (SPR) (Figure S5A), and could competitively inhibit the ACE2 binding with the RBD (Figure 5C and 5D).
To evaluate their neutralization potential against the authentic virus, we performed the plaque reduction neutralization test (PRNT) using authentic SARS-CoV-2 isolated from COVID-19 patients.
Among all mAbs tested, BD-368-2 displayed the highest potency against the authentic virus with an IC50 of 15 ng/mL (Figure 2B and 2C). To further validate the authentic virus neutralization shown by PRNT, we performed the cytopathic effect (CPE) inhibition assay using BD-218 on authentic SARS-CoV-2. BD-218 showed complete viral inhibition across all three replicates, at the concentration of 1.2 µg/mL (Figure
S6A and S6B), consistent with the PRNT results. Together, our data have shown that highly potent neutralizing mAbs could be identified from convalescent patients by high-throughput single-cell sequencing of antigen-binding B cells.
BD-368-2 showed high therapeutic and prophylactic efficacy in hACE2 mice
To evaluate whether the identified neutralizing mAbs could serve as therapeutic interventions and prophylactic protection against SARS-COV-2 in vivo, we tested the neutralization efficacy of BD-368-2 on hACE2 transgenic mice infected with SARS-CoV-2 (Bao et al., 2020).
BD-368-2 was chosen since it exhibits the highest potency against both pseudotyped and authentic SARS-CoV-2. A total of 9 hACE2 transgenic mice were used in this study, and were equally split into three groups (N=3).
The prophylactic efficacy was examined by intraperitoneally injecting 20 mg/kg (of mice body weight) BD-368-2 into hACE2 transgenic mice 24 h before viral infection (Figure 3A). For the therapeutic group, 20 mg/kg BD-368-2 was injected 2 h after infection. As a negative control, 20 mg/kg HG1K, an IgG1 antibody against the H7N9 virus, was injected 2 h after infection.
We recorded the body weight for each mice daily after infection for 5 days (Figure 3A), and found that the therapeutic and prophylactic treatment groups maintained their body weight, whereas the negative control group significantly lost weight (Figure 3B).
This indicates that BD-368-2 applied both before and after the infection could greatly improve the physiological condition of the SARS-CoV-2-infected mice. Moreover, we analyzed the viral load by qRT-PCR of the lungs at 5 dpi (Yang et al.,2007), and found that injections of BD-368-2 before the viral challenge could completely inhibit SARS-CoV-2 (Figure 3C).
Furthermore, applying BD-368-2 2 h after infection could result in a 3-4 log decrease in virus copies in mice lung, indicating an effective reduction of SARS-CoV-2 replication (Figure 3C). Together, BD-368-2 exhibits high therapeutic and prophylactic efficacy in vivo.
Cryo-EM structure of the neutralizing mAb bound to the trimeric spike protein
Neutralizing mAbs against SARS-CoV-2 were identified efficiently after analyzing enriched clonotypes. However, due to the shallow sampling of B cells for each patient, the majority of the clonotypes did not show any enrichment.
Nevertheless, the non-enriched antigen-binding clonotypes still remain as a valuable library for future identification of neutralizing mAbs. A new antibody selection criterion for non-enriched clonotypes is needed, since arbitrary selection may lead to a high percentage of weak antigen-binding mAbs, and is also prone to non-specific binding during the antigen pull-down.
Previously, people have used antibody structure prediction to annotate Ig-seq data for better mAbs selection (DeKosky et al., 2016). By selecting the mAbs that share similar CDR3H structures with reference neutralizing mAbs,
the structural annotation approach has proven to be successful in improving the performance of Ig-seq as well as in vitro antibody maturation (Kovaltsuk et al., 2017; Krawczyk et al., 2014; Sela-Culang et al., 2014).
Inspired by these results, we hypothesized that CDR3H-based structure prediction could also help to annotate the scVDJ-seq data from convalescent patients, which may serve to improve the efficiency in identifying neutralizing mAbs against SARS-CoV-2.
The main difficulty, however, lies in the selection of the reference antibody structure. To date, no SARS-CoV-2 neutralizing mAb structures are recorded in the Protein Database (PDB), which could serve as a reference.
To address this, we solved the cryo-EM structure of BD23-Fab in complex with the S ectodomain trimer at an overall resolution of 3.8 Å (Figure 4A and S7; Table S4). The S ectodomain here adopts an asymmetric conformation as previously reported (Wrapp et al., 2020), with the RBD in one protomer (mol A) adopting an “up” position, whereas the other two RBDs (mol B and C) adopt “down” positions.
In this 3D reconstruction, a single BD23-Fab is observed per S trimer, and it binds the “down” RBD in protomer B. Only the heavy chain variable domain of BD-23 is involved in binding to the RBD (Figure 4B). Interestingly, the binding is also facilitated by an N-linked glycan on Asn165 of protomer C.
Although the current resolution does not support further analysis of the molecular interactions between BD23-Fab and RBD, it is evident that BD23’s epitope overlaps with the motif recognized by ACE2. In fact, a comparison with the RBD-ACE2 complex structure demonstrates that BD23-Fab competes with ACE2 for binding to RBD (Figure 4C).
This observation suggests that the SARS-CoV-2 neutralization ability of BD-23 comes from the disruption of the ACE2-RBD binding, which is highly similar to previously identified neutralizing mAbs against SARS-CoV (Prabakaran et al., 2006; Yuan et al., 2020).
Indeed, several studies have suggested that SARS-CoV neutralizing mAbs could cross-neutralize SARS-CoV-2 (Wanga et al., 2020; Tian et al., 2020; Lv et al., 2020), which further highlights the resemblance between the RBDs of SARS-CoV and SARS-CoV-2.
Altogether, the structural observations and the high conservation between SARS-CoV and SARS-COV-2 suggest that it may also be feasible to use SARS-CoV neutralizing mAbs’ crystal structures as a reference, to screen for SARS-CoV-2 antigen-binding clonotypes that share similar CDR3H structures to identify neutralizing mAbs.
Selecting potent neutralizing mAbs based on the structural similarity of CDR3H
We compared every IgG1-presenting clonotype’s predicted structure based on CDR3H using FREAD to the available SARS-CoV neutralizing mAbs’ crystal structure in the PDB database as well as the structure of BD-23. Unfortunately, no IgG1-presenting clonotypes showed high CDR3H structural similarity with BD-23. However, a total of 12 IgG1 clonotypes showed
high structure similarity with PDB ID 2dd8 and 2ghw (Table S2; Figure 5C), which corresponds to previously isolated SARS-CoV neutralizing mAbs m396 and 80R, respectively (Prabakaran et al., 2006; Hwang et al., 2006).
The CDR3H sequence of the identified mAbs shared high homology with m396 (2dd8) (Figure 5A), which neutralizes SARS-CoV by disrupting ACE2/RBD binding (Figure 5D).
Surprisingly, we found that 7 out of the 12 mAbs exhibited a strong RBD-binding affinity and a potent neutralizing ability against pseudotyped SARS-CoV-2 (Figure 5B and 5C). The neutralization abilities of three representative mAbs, BD-503, BD-508, and BD-515, were also verified in the PRNT against authentic SARS-CoV-2 and showed high potencies (Figure 5E and 5F).
Moreover, these mAbs were mostly encoded by VH3-66/JH6 or VH3-53/JH6 gene segments for the heavy chain, and VLK1-9 or VLK1D-33/39 for the light chain (Figure 5C). Due to the similarity of amino acid sequences and VDJ combinations, we questioned whether these mAbs came from the same lineage.
After RNA mutation analysis, we found that none of the mAbs was from the same lineage nor the same patients. Together, these observations suggest that stereotypic B-cell receptors that bind to a particular SARS-CoV-2 epitope may exist in different individuals, similar to what has been observed in HIV, Influenza, and hepatitis C (Gorny et al., 2009; Ekiert et al., 2009; Marasca et al., 2001).
Indeed, epitope binning experiments using a double-antibodies sandwich ELISA has shown that BD-503, BD-508, and BD-515 likely share an overlapping epitope (Figure 6). However, more structural analysis is needed to obtain the exact epitopes to confirm whether they are largely conserved.









{kind=link}