











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



The lens of the eye is an unusual organ. Unlike most of the body’s organs, blood vessels don’t reach the lens. If they did, they’d obscure our vision and we wouldn’t be able to see.
The lack of vasculature led scientists to believe immune cells, which travel via the bloodstream, couldn’t get to this part of the body either.
But a few years ago, Jefferson researchers challenged this long held assumption by demonstrating that immune cells populate the lens in response to degeneration. Now the Jefferson team finds the eye also launches an immune response in the lens after injury.
The discovery adds to a growing body of evidence that is working to overturn the accepted dogma of the field.
“Why would we evolve a tissue that is so central to our being able to see without ways to ensure its protection, its ability to repair itself?” says, Sue Menko, Ph.D., Professor in the Department of Pathology, Anatomy and Cell Biology at Thomas Jefferson University, who led the research. “Immune cells are central to that protection and repair.”
The lens of the eye works like a camera lens. Its main purpose is to focus images coming in through the cornea – the transparent front layer of the eye – onto the retina at the back of the eye.
The images are detected by the retina and then translated in the brain as what we see. That lens must be crystal clear. As a result, scientists have always described the lens as a tissue without vasculature and therefore no source of immune cells either.
“At some point, you think about it and you wonder how that’s possible,” Dr. Menko says. “It doesn’t really make a lot of sense.”
The puzzle led Dr. Menko and her team to investigate whether immune cells are present in the eye. In a previous study, they discovered that when the lens is in a diseased state, immune cells are not only recruited there, but they also show up in the cornea, retina, and vitreous body – all parts of the eye that don’t normally have immune cells. Dr. Menko’s work suggested that the immune cells come from the ciliary body, a sort of muscle that helps squeeze and pull the lens, changing its shape, and helping it focus.
“The ciliary body is also a place that is vascular rich so it seemed like the most obvious place to look,” Dr. Menko says.
Now, in the latest work, Dr. Menko and colleagues show that after injury to the cornea, immune cells travel from the ciliary body to the lens along fibers known as ciliary zonules. The researchers used fluorescent markers and high-powered microscopes to observe structures of mouse eyes one day after receiving a scratch on the cornea.
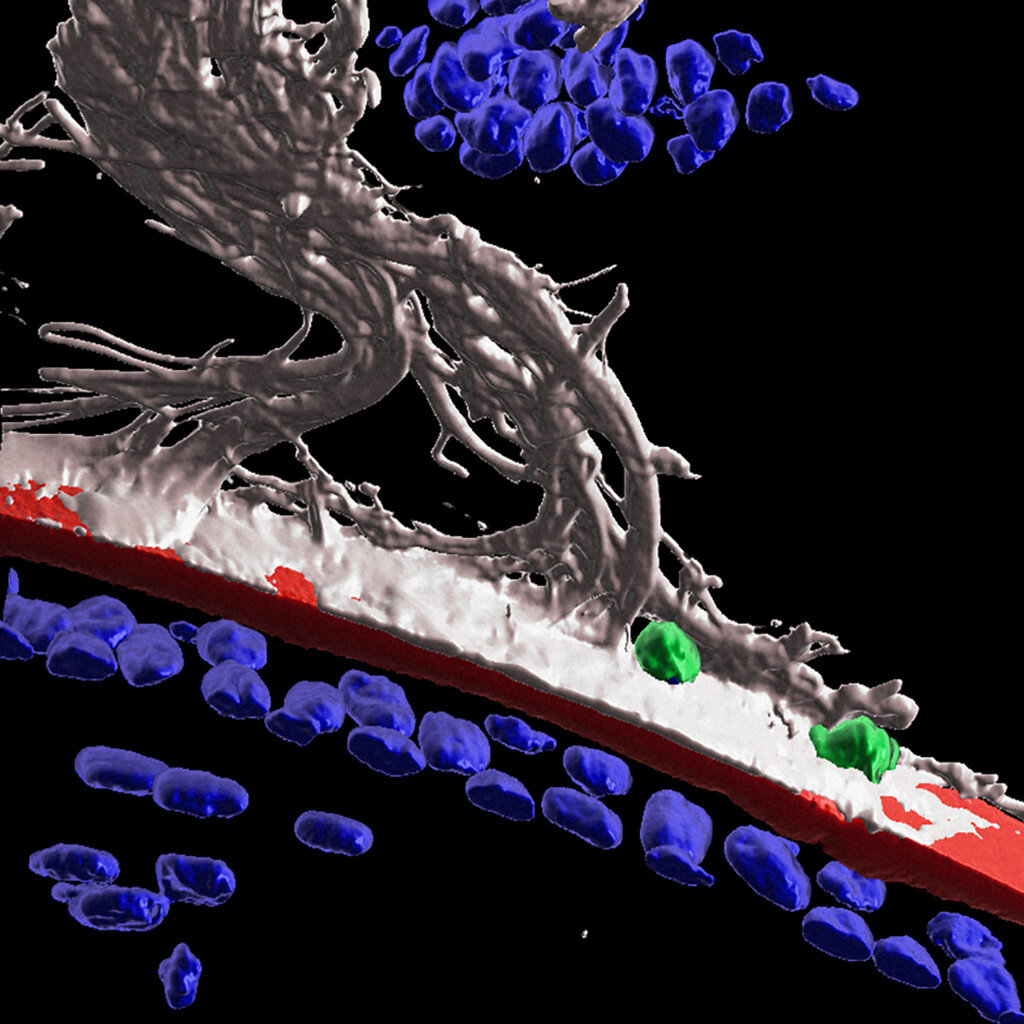
The high-tech imaging analysis Dr. Menko’s team used revealed that following injury to the cornea, the immune system launches a response to protect the lens. Immune cells are recruited to the lens via the ciliary zonules, and crawl along the surface of the lens to surveille and protect from adverse impacts of the corneal wound.
“This is really the first demonstration that surveillance by immune cells of the lens in response to injury somewhere else in the eye,” Dr. Menko says.
The researchers also found that some immune cells were able to cross the lens capsule, a membranous structure that helps to keep the lens under tension. The results could point to a role for immune cells in cataract formation.
Together, the findings indicate that in response to damage or disease, the eye utilizes alternative mechanisms – rather than direct contact with the bloodstream like non-transparent tissues do – to ensure that immune cells get to sites to provide healing and protection.
“We’re excited to go from thinking this doesn’t make sense to proving that the body is amazing and can adapt to anything. You just have to go in and look for it,” Dr. Menko says.
“We should be willing to challenge dogma because that’s where discovery is,” she adds. “It can enlighten what we know if we always keep our mind open to what doesn’t make sense and what maybe should be challenged to understand things better.”
Dr. Menko and colleagues published the results on May 25th in The FASEB Journal.
The ocular surface, which comprises of the cornea, limbus conjunctiva, and tear film, plays a key role in the visual system. Its integrity is crucial for the health and normal function of the eye, and as a result it is equipped with a delicate mucosal immune system to help prevent or limit damage.
It is comprised of both the innate and adaptive immune system, and the prevention of ocular surface diseases depends on the appropriate activation and regulation of these inter-connected branches of the immune system, which in turn rely on various molecular sensors and receptors.
This wide spectrum of diseases can occur as a result of pure infection, stemming from “external’” factors, such as contact lens injury, or ‘internal’ factors, including autoimmune diseases such as rheumatoid arthritis and Sjögren’s syndrome [1,2].
Ocular surface diseases represent a significant unmet medical need and are a substantial burden to patients, families, and healthcare systems. In the United States, annual medical costs per patient with only dry eye are estimated to be $783 per year, with overall cost calculated to be approximately $3.84 billion per year.
In addition, societal costs encompassing the loss of productivity are estimated at $11,302 per year, with the overall cost calculated to reach $55.4 billion per year [3]. Since there are only very limited numbers of effective and generalized treatments for this group of diseases, more investigation into its etiology and underlying mechanisms is crucial [4,5,6].
The innate nature of the immune cells (i.e., dendritic cells, macrophages, or natural killer cells) includes recognition of the wide variety of danger signals during infection, inflammation, and tissue injury.
Their adaptive component involves antigen-experienced memory-like cell characteristics, which includes the ability to produce cytokines, cytolytic molecules, and growth factors after insult from infection.
Among the innate immune cell types, dendritic cells (DCs) are an important player, forming an essential interface between the innate sensing of pathogens and the activation of the adaptive immune response [7]. DCs, if not γδ and resident memory T cells, are quite an important population in bridging the activation between the adaptive and innate immune system pathway.
It is inspiringly shown that there is an 80-fold increase in the number of myeloid DCs in the airway mucosa layer and in bronchoalveolar lavage fluid in a mouse or rat model of asthma. There are two pathways to control the inhalational tolerance: The first silences antigen (Ag)-reactive T cells, along with an induction of expanding regulatory T cells in the lymph nodes.
They ingest inhaled Ags and migrate to the draining lymph nodes with an upregulation of the homing receptor CCR7, and DCs activate T cells in the draining lymph nodes to induce tolerance. On the other hand, plasmacytoid DCs (pDCs) directly suppress the capacity of myeloid DCs to generate effector T cells and they can also stimulate the generation of Treg cells, possibly in an inducible T-cell co-stimulator ligand (ICOS-L)- dependent manner. Following short-term allergen inhalation, there is fast recruitment of myeloid DC precursors from the circulation to the airway mediated by a chemoattractant receptor signal relay initiated by CCR2, followed by CCR7, to complete the journey of DCs to the mesenteric lymph node for Th2 priming [7].
On the other hand, Langerhans cells (LCs), discovered by Paul Langerhans in the mid-19th century, functioning as skin-resident antigen presenting cells (APCs) was only recently appreciated [8].
Recently, LCs have shown to be important in the functions of the mucosal epithelial layer lining the ocular, vaginal, cervical, and oral surfaces [9,10,11]. LCs have been important in initiating the adaptive immune responses by presenting the antigens to the naïve T cells [12].
Similar to the asthma model, the DCs have been shown to play a significant role in the progression or control of autoimmune diseases. The mere change in the number of DCs though cannot be directly related to autoimmunity, alternation in their functionality might indicate autoimmune infestation [13], pDC-mediated production of type I interferon has been suggested as a common pathway that leads to pathogenesis in psoriasis, type I diabetes, and systemic lupus erythematosus (SLE).
Similar to our previous observation with the homogeneous presence of immune cell types in barrier tissues with a resident memory cell function, our current meta-analysis also attempts to find the quantitative traits of dendritic cells, especially LCs, in the mucosa of the ocular surface, like other mucosal organs (skin, lung, intestine, cervix, gastric mucosa) [6].
The ocular surface immune system has been investigated to have different immune cell components, both in the innate and adaptive branches and the phenotype, though the functions of those components are still under-investigated. We have selected a spectrum of ocular surface diseases in our study with underlying etiologies, both in the branches of infection and autoimmune diseases.
The selected diseases in our study are dry eye, glaucoma, Graft-vs-host disease (GvHD), keratitis, scleritis, Sjögren’s syndrome, and contact lens wear. We have mostly found the involvement of dendritic cells among our chosen studies, followed by cytokine IL6, IL17, a prominent helper (CD4+, Th), and cytotoxic (CD8+, Tc) cells.
Along with the predominant immune population contributing to the manifestation of the disease spectrum, we have conglomerated the innovative diagnostic methods. This paper has the vision of opening several new opportunities relating to the anti-inflammatory paradigm of treatments in ocular surface inflammatory disorders (OSIDs).
Results
Dendritic Cells are the Major Immune Cells Responsible for Ocular Surface Diseases
We divided the immune cell components into two major sections depending on their predominant functions. The major cells associated with the adaptive immune system are T- (Th, TC, TRM) and the sub-division of Th cells, such as Th17, Tregs, and the B-lymphocytes.
The major immune cells associated with the innate immune system are dendritic cells, platelets, neutrophils, monocytes, and macrophages. The raw data for each of the diseases to perform individual cell type analysis for adaptive and innate immune branches is provided in the Supplementary Figures S1–S8.
A representative example of the overlay of the innate and adaptive immune system is shown in Figure 2. Along with the immune cells, we also retrieved the importance of cytokines associated with both of the sub-divisions of the immune system.
Among the several cells and inflammatory mediators that are important for the ocular surface diseases, we found the number of research papers following this decreasing order: dendritic cells (19) > IL6 (15) > IL17 (8) > CD4+ and CD8+ T cells together (7). The eight diseases that matched the inclusion criteria of the study mentioned in the method section are: contact lens wear [15,16,17,18,19,20], corneal graft rejection [21,22,23], dry eye [6,24,25,26,27,28,29,30,31,32,33,34], glaucoma [35,36,37,38,39,40,41,42], Graft-vs-host (GvHD) [43,44], keratitis [45,46,47], scleritis [48,49,50], and Sjögren’s syndrome [28,29,32,33,34,51,52,53].
This is a combination of different ocular surface disorders, the range of whose etiologies lie from the infectious encounter to autoimmune factors. Among these diseases, we could not find any involvement of adaptive immune system factors in case of contact lens wear and GvHD.
Regarding the contact lens wear and GvHD (Supplementary Figures S1 and S5), we have found the significance of dendritic cells only (langerhans cells for the contact lens wear).
For the corneal graft rejection, combinations of innate (monocytes, macrophages) and adaptive (CD3, CD25) factors are responsible (Supplementary Figure S2). For the dry eye disease, we found the maximum number of innate and adaptive factors responsible for initiating the dendritic cells, the platelets to a series of cytokines (IL17, IL23, IL33, IL4, IL5, IL6, IL10, IFNγ, transforming growth factor (TGF)-β), and the chemokines (CCL2, CXCL3, CXCL9, CXCR4, CXCL10, CXCL11, CXCL12) in the adaptive component (Supplementary Figure S3).
Dry eye disease is a multi-factorial disease, in which the clinical characteristics can be either systematic (SLE, rheumatoid arthritis (RA), hypothyroidism), or idiopathic, and sometimes requiring ocular surgery.
This was properly displayed through the huge number of immune factors responsible for dry eye disease in comparison to other diseases. For the studies of glaucoma, we also found many factors common to the dry eye disease, along with the plasma cells in the adaptive part (Supplementary Figure S4).
For the infection-related keratitis, we considered the papers related to the fungal, bacterial, and stromal keratitis. Among the different components, the intriguing papers were those involved with CD19, CD20, anti-inflammatory component IL10, and pro-inflammatory component IL1β (Supplementary Figure S6). For scleritis, the one component which stands out, both in the innate and adaptive component, is the myeloid cell markers (Supplementary Figure S7). For Sjögren’s syndrome, the adaptive components play major roles in comparison to the innate factors. The one adaptive component most mentioned is IL17 (Supplementary Figure S8).

Analysis of different immune cell components in the eight chosen ocular surface diseases. The global standard mean difference for each disease obtained by our proposed meta-analysis is shown as a part of adaptive immune system (A) and innate immune system (B). The diamonds represent the standard mean difference and the 95% confidence interval for each disease. It can be seen that the presence of immune cells is higher for every analyzed disease. A detailed description of each of the diseases and the different cells involved can be found in the Supplementary Material (Supplementary Figures S1–S8).
Discussion
Ocular surface disorder is a complicated disorder in the sense that there might be an overlap between different diseases, and the etiologies of the diseases can also be extremely variable. We can take an example of dry eye and Sjögren’s syndrome to explain the complexity.
In most of the references studied, DED patients were divided into two groups—the Sjögren’s syndrome group and the non-Sjögren’s syndrome group. The clinical features checked for both the diseases are dry eye symptoms (>3 months), high OSDI scores (>13 points), abnormal TBUT (<10 s), low Schirmer I test (<10 mm), and positive corneal fluorescein staining.
However, the distinction between these two diseases is not quite clear, but the researchers are continuously making different innovative efforts to diagnose the different forms. For example, the American–European consensus group in the year of 2002 revised this classification and included oral symptoms, salivary gland involvement, serum autoantibodies, rheumatoid factors, and anti-double-stranded DNA (dsDNA).
Along with the difficulty of diagnosis, multiple etiologies can be involved in dry eye diseases or Sjögren’s syndrome-like diseases, including autoimmune diseases, ageing, medications, refractive surgery, habits, diet, vitamin D, and environmental factors [24,26,28].
What can be reflected from this example is that it is not very easy to classify ocular surface diseases, but they can be represented as a spectrum in which they reflect a range of phenotypic features from mild to severe.
DCs are central to the immune memory activation and tolerance induction. DC development takes place in the bone marrow and has tissue-residency and migratory characteristics, predominantly due to its requirement to replenish the mature DCs in peripheral tissues [55,56,57].
DCs can be classified into four broad subsets, according to the phenotype, function, and developmental origin, and they each play distinct roles in immune responses. DC classification includes plasmacytoid DCs (pDCs), tissue-resident and migrators conventional DCs (cDCs), Langehans cells (LCs), and monocyte-derived DCs (mDCs). This classification of DCs is consistent between mice and humans.
There is a functional and structural difference between the tissue-resident and migratory DCs. Tissue-resident DCs express different receptors for microbe-associated molecular patterns (MAMPs) and damage-associated molecular patterns (DAMPs), and these are activated by the infections and promote the production of pro- or anti-inflammatory cytokines. After capturing different antigens, DCs migrate through the lymphatics to reach the secondary lymphoid organs, where DCs present processed antigenic peptides to stimulate the naïve T cells.
The tissue-resident DCs exist in several forms with several markers in different organs, according to previous literature. Various organs where DCs display resident immune cell functions are the skin, blood, lungs, liver, gastrointestinal (GI)-tract, kidney, spleen, and lymph node. Tissue-resident DCs contribute to the prognosis of a range of diseases, such as allergies, auto-immunities, inflammation, and cancer progression [58,59,60,61,62].
It is not only the tissue-resident population but the migratory DCs as well that contribute hugely to the overall functionality of any organ [18]. For example, in mice kidneys, the DCs are replaced within 2–4 weeks after lethal irradiation. In contrast, the replacement of migratory DCs is much faster (within 7–13 days) in the vagina, airway epithelia, and gut.
In the steady-state, DC progenitors migrate form the bone marrow through the blood to the peripheral tissues, where they are designated to raise distinct subsets of myeloid DCs after a final differentiation with the exception of Langerhans cells, which resided in the epidermis of skin independent of its circulating precursors.
The human and mice DCs have been investigated intensively in previous literatures, but our article is mainly focused on the human experiments, except for a few examples of the animal models [58,63,64,65].
In our current study, we made an effort to characterize the ocular surface diseases with a common mechanism. Dendritic cells are the cornerstone of those diseases and can activate the differentiated T cells in different ways, mediating through many chemokines and cytokines.
An easier explanation of this would be to activate the Th1 differentiated pathway with the mediator of IL12, which in turn secretes IFNγ, which keeps the production of Th1 cells in a negative feedback loop.
Same mechanisms can be seen in the case of Th2 cells, but feedback, with the help of IL6 and IL4, is formed. In the case of Th17 and Treg pathways, these are controlled by IL17, IL23, and IL10 pathways. We also have seen the difference in the mechanism in different ocular diseases, except the contact lens wear, which is an entirely innate immune system phenomenon, involving only the Langerhans cells.
In the case of glaucoma, GvHD, and scleritis, we have seen the factors regulating the Th1 pathway, whereas Sjögren’s syndrome is regulated by the Th2 pathway. On the other hand, there is involvement of Th17 pathways in the case of glaucoma, DED, scleritis, and Sjögren’s syndrome and the Treg pathway in the case of scleritis [7,66,67,68,69,70].
The reliability of these common mechanisms in different diseases can be compared with the mouse studies, which are beyond the capacity of this manuscript. Overall, this manuscript represents a combination of different factors through which ocular surface disorders can be prevented with the help of several cells and mediators, the key of which is in the activity of dendritic cells, predominantly the Langerhans type.
The research in the field of Langerhans cells is overlooked in the case of skin immunity, the reason being that the deletion of Langerhans cells does not result in fatal susceptibility to a skin infection nor due to an overt autoimmunity due to lack of immune regulation.
The eye, being the mucosal organ similar to the skin, also represents the importance of Langerhans cells in maintaining and modulating the tissue environment in shaping the effector immunity [71]. More understanding in the field of Langerhans cells will develop new possibilities for therapeutic approaches in the field of ocular surface diseases.
Pitfalls and Future Perspectives
This study is a conglomeration of the observations from the publications of the last 20 years, but this is already a hint to the research direction in the field of ocular surface disorders. The number of references we have worked with did not represent a huge number—this is one of the limitations of the study.
In addition, we reflected the phenotypic features of ocular surface diseases with the help of eight selected diseases. In spite of these two limitations, this study gives an indication as to which immune cells or which immune mediator can able us to alleviate the ocular surface diseases. Future studies with the knockout models with the deletion of genes for the important cytokine and chemokine factors, such as IL2 and IL12, will give new insight into the field of research.
References
1. Garcia D.M., Reis de Oliveira Modulo C.M., Faustino JBarbosa A.P., Alves M., Rocha E.M. Is Sjögren’s syndrome dry eye similar to dry eye caused by other etiologies? Discriminating different diseases by dry eye tests. PLoS ONE. 2018;13:e0208420. doi: 10.1371/journal.pone.0208420. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
2. Milner M.S., Beckman K.A., Luchs J.I., Allen Q.B., Awdeh R.M., Berdahl J., Boland T.S., Buznego C., Gira J.P., Goldber D.F., et al. Dysfunctional tear syndrome: Dry eye diseases and associated tear film disorders—New strategies for diagnosis and treatments. Curr. Opin. Opthalmol. 2017:3–47. doi: 10.1097/01.icu.0000512373.81749.b7. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
3. Yu J., Asche C.V., Fairchild C.J. The economic burden of dry eye disease in the United States: A decision tree analysis. Cornea. 2011;30:379–387. doi: 10.1097/ICO.0b013e3181f7f363. [PubMed] [CrossRef] [Google Scholar]
4. Tong L., Lan W., Lim R.R., Chaurasia S.S. S100A proteins as molecular targets in the ocular surface inflammatory diseases. Ocul. Surf. 2014;12:23–31. doi: 10.1016/j.jtos.2013.10.001. [PubMed] [CrossRef] [Google Scholar]
5. Shih K.C., Lam K.S., Tong L. A systematic review on the impact of diabetes mellitus on the ocular surface. Nutr Diabetes. 2017;7:e251. doi: 10.1038/nutd.2017.4. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
6. Bose T., Lee R., Hou A., Tong L., Chandy K.G. Tissue resident memory T cells in the human conjunctiva and immune signatures in human dry eye disease. Sci. Rep. 2017;7:45312. doi: 10.1038/srep45312. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
7. Walsh K.P., Mills K.H. Dendritic cells and other innate determinants of T helper cell polarisation. Trends Immunol. 2013;34:521–530. doi: 10.1016/j.it.2013.07.006. [PubMed] [CrossRef] [Google Scholar]
8. Katz S.I., Tamaki K., Sachs D.H. Epidermal Langerhans cells are derived from cells originating in bone marrow. Nature. 1979;282:324–326. doi: 10.1038/282324a0. [PubMed] [CrossRef] [Google Scholar]
9. Hamrah P., Huq S.O., Liu Y., Zhang Q., Dana M.R. Corneal immunity is mediated by heterogeneous population of antigen-presenting cells. J. Leukoc. Biol. 2003;74:172–178. doi: 10.1189/jlb.1102544. [PubMed] [CrossRef] [Google Scholar]
10. Miller C.J., Shattock R.J. Target cells in vaginal HIV transmission. Microbes Infect. 2003;5:59–67. doi: 10.1016/S1286-4579(02)00056-4. [PubMed] [CrossRef] [Google Scholar]
11. Valladeau J., Saeland S. Cutaneous dendritic cells. Semin. Immunol. 2005;17:273–283. doi: 10.1016/j.smim.2005.05.009. [PubMed] [CrossRef] [Google Scholar]
12. Stoitzner P., Tripp C.H., Eberhart A., Price K.M., Jung J.Y., Bursch L., Ronchese F., Romani N. Langerhans cells cross-present antigen derived from skin. Proc. Natl. Acad. Sci. USA. 2006;103:7783–7788. doi: 10.1073/pnas.0509307103. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
13. Ganguly D., Haak S., Sisirak V., Reizis B. The role of dendritic cells in autoimmunity. Nat. Rev. Immunol. 2013;13:566–577. doi: 10.1038/nri3477. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
14. Stroup D.F., Berlin J.A., Morton S.C., Olkin I., Williamson G.D., Rennie D., Moher D., Becker B.J., Sipe T.A., Thacker S.B. Meta-analysis of observational studies in epidemiology: A proposal for reporting. Meta-analysis Of Observational Studies in Epidemiology (MOOSE) group. JAMA. 2000;283:2008–2012. doi: 10.1001/jama.283.15.2008. [PubMed] [CrossRef] [Google Scholar]
15. Alzahrani Y., Pritchard N., Efron N. Changes in corneal Langerhans cell density during the first few hours of contact lens wear. Cont. Lens Anterior Eye. 2016;39:307–310. doi: 10.1016/j.clae.2016.02.008. [PubMed] [CrossRef] [Google Scholar]
16. Alzahrani Y., Colorado L.H., Pritchard N., Efron N. Longitudinal changes in Langerhans cell density of the cornea and conjunctiva in contact lens-induced dry eye. Clin. Exp. Optom. 2017;100:33–40. doi: 10.1111/cxo.12399. [PubMed] [CrossRef] [Google Scholar]
17. Efron N., Al-Dossari M., Pritchard N. Confocal microscopy of the bulbar conjunctiva in contact lens wear. Cornea. 2010;29:43–52. doi: 10.1097/ICO.0b013e3181acf82a. [PubMed] [CrossRef] [Google Scholar]
18. Lopez-De La Rosa A., Arroyo-Del Arroyo C., Canadas P., Lopez-Miguel A., Calonge M., Enriquez-De-Salamanca A., Gonzalez-Garcia M.J. Are Contact Lens Discomfort or Soft Contact Lens Material Properties Associated with Alterations in the Corneal Sub-Basal Nerve Plexus? Curr. Eye Res. 2018;43:487–492. doi: 10.1080/02713683.2017.1420804. [PubMed] [CrossRef] [Google Scholar]
19. Sindt C.W., Grout T.K., Critser D.B., Kern J.R., Meadows D.L. Dendritic immune cell densities in the central cornea associated with soft contact lens types and lens care solution types: A pilot study. Clin. Ophthalmol. 2012;6:511–519. doi: 10.2147/OPTH.S28083. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
20. Zhivov A., Stave J., Vollmar B., Guthoff R. In vivo confocal microscopic evaluation of langerhans cell density and distribution in the corneal epithelium of healthy volunteers and contact lens wearers. Cornea. 2007;26:47–54. doi: 10.1097/ICO.0b013e31802e3b55. [PubMed] [CrossRef] [Google Scholar]
21. Funding M., Vorum H., Nexo E., Moestrup S.K., Ehlers N., Moller M.J. Soluble CD163 and interleukin-6 are increased in aqueous humour from patients with endothelial rejection of corneal grafts. Acta Ophthalmol. Scand. 2005;83:234–239. doi: 10.1111/j.1600-0420.2005.00397.x. [PubMed] [CrossRef] [Google Scholar]
22. Kuffova L., Holan V., Lumsden L., Forrester J.V., Filipec M. Cell subpopulations in failed human corneal grafts. Br. J. ophthalmol. 1999;83:1364–1369. doi: 10.1136/bjo.83.12.1364. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
23. Wang D., Song P., Wang S., Sun D., Wang Y., Zhang Y., Gao H. Laser Scanning In Vivo Confocal Microscopy of Clear Grafts after Penetrating Keratoplasty. BioMed Res. Int. 2016;2016:5159746. doi: 10.1155/2016/5159746. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
24. Celik T. Assessment of Neutrophil-to-Lymphocyte Ratio and Platelet-to—Lymphocyte Ratio in Patients with Dry Eye Disease. Ocul. Immunol. Inflamm. 2018;26:1219–1222. doi: 10.1080/09273948.2017.1340486. [PubMed] [CrossRef] [Google Scholar]
25. Ji Y.W., Mittal S.K., Hwang H.S., Chang E.J., Lee J.H., Seo Y., Yeo A., Noh H., Lee H.S., Chauhan S.K. Lacrimal gland-derived IL-22 regulates IL-17-mediated ocular mucosal inflammation. Mucosal Immunol. 2017;10:1202–1210. doi: 10.1038/mi.2016.119. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
26. Kheirkhah A., Rahimi Darabad R., Cruzat A., Hajrasouliha A.R., Witkin D., Wong N., Dana R., Hamrah P. Corneal Epithelial Immune Dendritic Cell Alterations in Subtypes of Dry Eye Disease: A Pilot In Vivo Confocal Microscopic Study. Investig. Ophthalmol. Vis. Sci. 2015;56:7179–7185. doi: 10.1167/iovs.15-17433. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
27. Kheirkhah A., Saboo U.S., Abud T.B., Dohlman T.H., Arnoldner M.A., Hamrah P., Dana R. Reduced Corneal Endothelial Cell Density in Patients with Dry Eye Disease. Am. J. Ophthalmol. 2015;159:1022–1026.e2. doi: 10.1016/j.ajo.2015.03.011. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
28. Liu R., Gao C., Chen H., Li Y., Jin Y., Qi H. Analysis of Th17-associated cytokines and clinical correlations in patients with dry eye disease. PLoS ONE. 2017;12:e0173301. doi: 10.1371/journal.pone.0173301. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
29. Luo G., Xin Y., Qin D., Yan A., Zhou Z., Liu Z. Correlation of interleukin-33 with Th cytokines and clinical severity of dry eye disease. Indian J. Ophthalmol. 2018;66:39–43. doi: 10.4103/ijo.IJO_405_17. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
30. Nicolle P., Liang H., Reboussin E., Rabut G., Warcoin E., Brignole-Baudouin F., Melik-Parsadaniantz S., Baudouin C., Labbe A., Reaux-Le Goazigo A. Proinflammatory Markers, Chemokines, and Enkephalin in Patients Suffering from Dry Eye Disease. Int. J. Mol. Sci. 2018;19:1221. doi: 10.3390/ijms19041221. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
31. Shetty R., Sethu S., Deshmukh R., Deshpande K., Ghosh A., Agarwal A., Shroff R. Corneal Dendritic Cell Density Is Associated with Subbasal Nerve Plexus Features, Ocular Surface Disease Index, and Serum Vitamin D in Evaporative Dry Eye Disease. BioMed Res. Int. 2016;2016:4369750. doi: 10.1155/2016/4369750. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
32. Tan X., Sun S., Liu Y., Zhu T., Wang K., Ren T., Wu Z., Xu H., Zhu L. Analysis of Th17-associated cytokines in tears of patients with dry eye syndrome. Eye (Lond.) 2014;28:608–613. doi: 10.1038/eye.2014.38. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
33. Tepelus T.C., Chiu G.B., Huang J., Huang P., Sadda S.R., Irvine J., Lee O.L. Correlation between corneal innervation and inflammation evaluated with confocal microscopy and symptomatology in patients with dry eye syndromes: A preliminary study. Graefes Arch. Clin. Exp. Ophthalmol. 2017;255:1771–1778. doi: 10.1007/s00417-017-3680-3. [PubMed] [CrossRef] [Google Scholar]
34. Yoon K.C., Park C.S., You I.C., Choi H.J., Lee K.H., Im S.K., Park H.Y., Pflugfelder J.C. Expression of CXCL9, -10, -11, and CXCR3 in the tear film and ocular surface of patients with dry eye syndrome. Investig. Ophthalmol. Vis. Sci. 2010;51:643–650. doi: 10.1167/iovs.09-3425. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
35. Baghdasaryan E., Tepelus T.C., Vickers L.A., Huang P., Chopra V., Sadda S.R., Lee O.L. Assessment of Corneal Changes Associated with Topical Antiglaucoma Therapy Using in vivo Confocal Microscopy. Ophthalmic Res. 2019;61:51–59. doi: 10.1159/000484632. [PubMed] [CrossRef] [Google Scholar]
36. Gramlich O.W., Beck S., von Thus Und Hohenstein-Blaul N., Boehm N., Ziegler A., Vetter J.M., Pfeiffer N., Grus F.H. Enhanced insight into the autoimmune component of glaucoma: IgG autoantibody accumulation and pro-inflammatory conditions in human glaucomatous retina. PLoS ONE. 2013;8:e57557. doi: 10.1371/journal.pone.0057557. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
37. Liang H., Baudouin C., Hamard P., Creuzot-Garcher C., Warnet J.M., Brignole-Baudouin F. Activation of TH1/TH2 pathways detected through the expression of CCR4 and CCR5 on the ocular surface of glaucomatous patients treated over the long term. J. Fr. Ophtalmol. 2006;29:121–126. doi: 10.1016/S0181-5512(06)73759-7. [PubMed] [CrossRef] [Google Scholar]
38. Malvitte L., Montange T., Vejux A., Baudouin C., Bron A.M., Creuzot-Garcher C., Lizard G. Measurement of inflammatory cytokines by multicytokine assay in tears of patients with glaucoma topically treated with chronic drugs. Br. J. Ophthalmol. 2007;91:29–32. doi: 10.1136/bjo.2006.101485. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
39. Margeta M.A., Lad E.M., Proia A.D. CD163+ macrophages infiltrate axon bundles of postmortem optic nerves with glaucoma. Graefes Arch. Clin. Exp. Ophthalmol. 2018;256:2449–2456. doi: 10.1007/s00417-018-4081-y. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
40. Mastropasqua R., Agnifili L., Fasanella V., Lappa A., Brescia L., Lanzini M., Oddone F., Perri P., Mastropasqua L. In Vivo Distribution of Corneal Epithelial Dendritic Cells in Patients with Glaucoma. Investig. Ophthalmol. Vis. Sci. 2016;57:5996–6002. doi: 10.1167/iovs.16-20333. [PubMed] [CrossRef] [Google Scholar]
41. Taurone S., Ripandelli G., Pacella F., Bianchi E., Plateroti A.M., De Vito S., Plateroti P., Grippaudo F.R., Cavallotti C., Artico M. Potential regulatory molecules in the human trabecular meshwork of patients with glaucoma: Immunohistochemical profile of a number of inflammatory cytokines. Mol. Med. Rep. 2015;11:1384–1390. doi: 10.3892/mmr.2014.2772. [PubMed] [CrossRef] [Google Scholar]
42. Wong M., Huang P., Li W., Li Y., Zhang S.S., Zhang C. T-helper1/T-helper2 cytokine imbalance in the iris of patients with glaucoma. PLoS ONE. 2015;10:e0122184. doi: 10.1371/journal.pone.0122184. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
43. He J., Ogawa Y., Mukai S., Saijo-Ban Y., Kamoi M., Uchino M., Yamane M., Ozawa N., Fukui M., Mori T. In Vivo Confocal Microscopy Evaluation of Ocular Surface with Graft-Versus-Host Disease-Related Dry Eye Disease. Sci. Rep. 2017;7:10720. doi: 10.1038/s41598-017-10237-w. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
44. Kheirkhah A., Qazi Y., Arnoldner M.A., Suri K., Dana R. In Vivo Confocal Microscopy in Dry Eye Disease Associated with Chronic Graft-Versus-Host Disease. Investig. Ophthalmol. Vis. Sci. 2016;57:4686–4691. doi: 10.1167/iovs.16-20013. [PubMed] [CrossRef] [Google Scholar]
45. Cruzat A., Witkin D., Baniasadi N., Zheng L., Ciolino J.B., Jurkunas U.V., Chodosh J., Pavan-Langston D., Dana R., Hamrah P. Inflammation and the nervous system: The connection in the cornea in patients with infectious keratitis. Investig. Ophthalmol. Vis. Sci. 2011;52:5136–5143. doi: 10.1167/iovs.10-7048. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
46. Iskeleli G., Camcioglu Y., Akova N., Kiran B., Bahar H., Deniz G. Lymphocyte subgroups and natural killer cell activity in recurrent herpetic stromal keratitis. Eye Contact Lens. 2008;34:169–173. doi: 10.1097/ICL.0b013e318157a5c3. [PubMed] [CrossRef] [Google Scholar]
47. Yamaguchi T., Calvacanti B.M., Cruzat A., Qazi Y., Ishikawa S., Osuka A., Lederer J., Hamrah P. Correlation between human tear cytokine levels and cellular corneal changes in patients with bacterial keratitis by in vivo confocal microscopy. Investig. Ophthalmol. Vis. Sci. 2014;55:7457–7466. doi: 10.1167/iovs.14-15411. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
48. Bernauer W., Watson P.G., Daicker B., Lightman S. Cells perpetuating the inflammatory response in scleritis. Br. J. Ophthalmol. 1994;78:381–385. doi: 10.1136/bjo.78.5.381. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
49. Fong L.P., Sainz de la Maza M., Rice B.A., Kupferman A.E., Foster C.S. Immunopathology of scleritis. Ophthalmology. 1991;98:472–479. doi: 10.1016/S0161-6420(91)32280-2. [PubMed] [CrossRef] [Google Scholar]
50. Sainz de la Maza M., Foster C.S. Necrotizing scleritis after ocular surgery. A clinicopathologic study. Ophthalmology. 1991;98:1720–1726. doi: 10.1016/S0161-6420(91)32062-1. [PubMed] [CrossRef] [Google Scholar]
51. Fujihara T., Fujita H., Tsubota K., Saito K., Tsuzaka K., Abe T., Takeuchi T. Preferential localization of CD8+ alpha E beta 7+ T cells around acinar epithelial cells with apoptosis in patients with Sjogren’s syndrome. J. Immunol. 1999;163:2226–2235. [PubMed] [Google Scholar]
52. Pflugfelder S.C., Bian F., Gumus K., Farley W., Stern M.E., De Paiva C.S. Severity of Sjogren’s Syndrome Keratoconjunctivitis Sicca Increases with Increased Percentage of Conjunctival Antigen—Presenting Cells. Int. J. Mol. Sci. 2018;19:2760. doi: 10.3390/ijms19092760. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
53. Williams G.P., Nighingale P., Southworth S., Denniston A.K., Tomlinus P.J., Turner S., Hamburger J., Bowman S.J., Curnow S.J., Rauz S. Conjunctival Neutrophils Predict Progressive Scarring in Ocular Mucous Membrane Pemphigoid. Investig. Ophthalmol. Vis. Sci. 2016;57:5457–5469. doi: 10.1167/iovs.16-19247. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
54. Harden J.L., Egilmez N.K. Indolamine 2,3-dioxygenase and dendritic cell tolerogenicity. Immunol. Investig. 2012;41:738–764. doi: 10.3109/08820139.2012.676122. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
55. Collin M., McGovern N., Haniffa M. Human dendritic cell subsets. Immunology. 2013;140:22–30. doi: 10.1111/imm.12117. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
56. Palucka K., Banchereau J. Human dendritic cell subsets in vaccination. Curr. Opin. Immunol. 2013;25:396–402. doi: 10.1016/j.coi.2013.05.001. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
57. Segura E., Amigorena S. Cross-presentation by human dendritic cell subsets. Immunol. Lett. 2014;158:73–78. doi: 10.1016/j.imlet.2013.12.001. [PubMed] [CrossRef] [Google Scholar]
58. Segura E., Valladeau-Guilemond J., Donnadieu M.-H., Sastre-Garau X., Soumelis V., Amigorena S. Characterization of resident and migratory dendritic cells in human lymph nodes. J. Exp. Med. 2012;209:653–660. doi: 10.1084/jem.20111457. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
59. Durand M., Segura E. The known unknowns of the human dendritic cell network. Front. Immunol. 2015;6:129. doi: 10.3389/fimmu.2015.00129. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
60. Granot T., Senda T., Carpenter D.J., Matsuoka N., Weiner J., Gordon C.L., Miron M., Kumar B.V., Griesemer A., Ho S.H. Dendritic cells display subset and tissue-specific maturation dynamics over human life. Immunity. 2017;46:504–515. doi: 10.1016/j.immuni.2017.02.019. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
61. Rhodes J.W., Tong O., Harman A.N., Turville S.G. Human dendritic cell subsets, ontogeny, and impact on HIV infection. Front. Immunol. 2019;10:1088. doi: 10.3389/fimmu.2019.01088. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
62. Chen K., Wang J.M., Yuan R., Yi X., Li L., Gong W., Yang T., Li L., Su S. Tissue-resident dendritic cells and diseases involving dendritic cell malfunction. Int. Immunopharmacol. 2016;34:1–15. doi: 10.1016/j.intimp.2016.02.007. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
63. Enamorado M., Khouili S.C., Iborra S., Sancho D. Genealogy, Dendritic Cell Priming, and Differentiation of Tissue-Resident Memory CD8(+) T Cells. Front. Immunol. 2018;9:1751. doi: 10.3389/fimmu.2018.01751. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
64. Mueller S.N. Spreading the load: Antigen transfer between migratory and lymph node-resident dendritic cells promotes T-cell priming. Eur. J. Immunol. 2017;47:1798–1801. doi: 10.1002/eji.201747248. [PubMed] [CrossRef] [Google Scholar]
65. Vitali C., Mingozzi F., Broggi A., Barresi S., Zolezzi F., Bayry J., Raimondi G., Zanoni I., Granucci F. Migratory, and not lymphoid-resident, dendritic cells maintain peripheral self-tolerance and prevent autoimmunity via induction of iTreg cells. Blood. 2012;120:1237–1245. doi: 10.1182/blood-2011-09-379776. [PubMed] [CrossRef] [Google Scholar]
66. Hunter C.A., Jones S.A. IL-6 as a keystone cytokine in health and disease. Nat. Immunol. 2015;16:448–457. doi: 10.1038/ni.3153. [PubMed] [CrossRef] [Google Scholar]
67. Kim B., Kim T.H. Fundamental role of dendritic cells in inducing Th2 responses. Korean J. Intern. Med. 2018;33:483–489. doi: 10.3904/kjim.2016.227. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
68. Agalioti T., Villablanca E.J., Huber S., Gagliani N. TH17cell plasticity: The role of dendritic cells and molecular mechanisms. J. Autoimmune. 2018;87:50–60. doi: 10.1016/j.jaut.2017.12.003. [PubMed] [CrossRef] [Google Scholar]
69. Sehrawat S., Rouse B.T. Interplay of Regulatory T Cell and Th17 Cells during Infectious Diseases in Humans and Animals. Front. Immunol. 2017;8:341. doi: 10.3389/fimmu.2017.00341. [PMC free article] [PubMed] [CrossRef] [Google Scholar]
70. Eisenstein E.M., Williams C.B. The T(reg)/Th17 cell balance: A new paradigm for autoimmunity. Pediatr. Res. 2009;65:26R–31R. doi: 10.1203/PDR.0b013e31819e76c7. [PubMed] [CrossRef] [Google Scholar]
71. Rajesh A., Wise L., Hibma M. The role of Langerhans cells in pathologies of the skin. Immunol. Cell Biol. 2019 doi: 10.1111/imcb.12253. [PubMed] [CrossRef] [Google Scholar]
More information: JodiRae DeDreu, Caitlin J. Bowen, Caitlin M. Logan, Sonali Pal-Ghosh, Paola Parlanti, Mary Ann Stepp, and A. Sue Menko, “An immune response to the avascular lens following wounding of the cornea involves ciliary zonule fibrils,” The FASEB Journal, DOI: 10.1096/fj.202000289R , 2020.



{kind=link}