")










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Elderly people are more prone to infectious diseases as the function of their immune system continuously declines with progression of age.
This becomes especially apparent during seasonal influenza outbreaks or the occurrence of other viral diseases such as COVID-19. As the efficacy of vaccination in the elderly is strongly reduced, this age group is particularly vulnerable to such infectious pathogens and often shows the highest mortality rate.
In addition to the age-related immune decline aged individuals are commonly affected by frailty that negatively impacts quality-of-life. Even though the average life-expectancy for humans continuous to rise, living longer is often associated with age-related health issues.
Important role of belly fat in aging processes identified
Researchers from the Department for BioMedical Reserarch (DBMR) and the Institute of Pathology at the University of Bern as well as the University Hospital Bern (Inselspital) have set out to identify new approaches to improve health-span in a fast-growing aging population.
For many years scientists speculated that chronic low-grade inflammation accelerates aging processes and the development of age-related disorders.
An international team of researchers under Bernese guidance has now demonstrated that visceral adipose tissue, known as belly fat, crucially contributes to the development of chronic low-grade inflammation. Scientist around Dr. Mario Noti, formerly at the Institute of Pathology of the University of Bern and Dr. Alexander Eggel from the Department for BioMedical Research (DBMR) of the Universität of Bern reported that certain immune cells in the belly fat play and an essential role in regulating chronic low-grade inflammation and downstream aging processes.
They could show, that these immune cells may be used to reverse such processes. The findings of this study have been published in the scientific journal Nature Metabolism and were further highlighted by a News and Views editorial article.
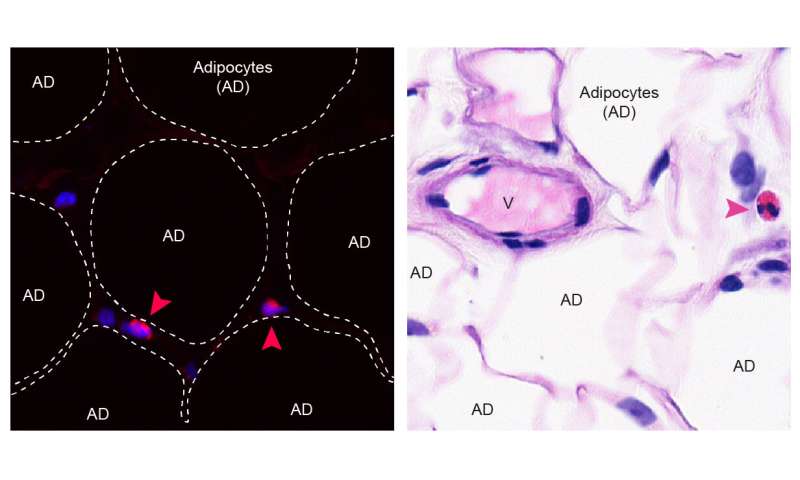
Belly fat as a source of chronic inflammation
The team around Dr. Noti and Dr. Eggel could demonstrated that a certain kind of immune cells, known as eosinophils, which are predominantly found in the blood circulation, are also present in belly fat of both humans and mice.
Although classically known to provide protection from parasite infection and to promote allergic airway disease, eosinophils located in belly fat are responsible to maintain local immune homeostasis.
With increasing age the frequency of eosinophils in belly fat declines, while the number of pro-inflammatory macrophages increases. Owing to this immune cell dysbalance, belly fat turns into a source of pro-inflammatory mediators accumulating systemically in old age.
Eosinophil cell therapy promotes rejuvenation
In a next step, the researchers investigated the possibility to reverse age-related impairments by restoring the immune cell balance in visceral adipose tissue.
“In different experimental approaches, we were able to show that transfers of eosinophils from young mice into aged recipients resolved not only local but also systemic low-grade inflammation,” says Dr. Eggel.
“In these experiments, we observed that transferred eosinophils were selectively homing into adipose tissue,” adds Dr. Noti. This approach had a rejuvenating effect on the aged organism.
As a consequence, aged animals showed significant improvements in physical fitness as assessed by endurance and grip strength tests. Moreover, the therapy had a rejuvenating effect on the immune system manifesting in improved vaccination responses of aged mice.
Translating findings into clinics
“Our results indicate that the biological processes of aging and the associated functional impairments are more plastic than previously assumed,” states Dr. Noti. Importantly, the observed age-related changes in adipose immune cell distribution in mice were also confirmed in humans.
“A future direction of our research will be to now leverage the gained knowledge for the establishment of targeted therapeutic approaches to promote and sustain healthy aging in humans,” says Dr. Eggel.
Neutrophils and Eosinophils
Neutrophils are the most abundant population of white blood cells in humans, playing important roles in the inflammatory process and primary immune response [123]. They have been mainly described as short-living immune cells and for their role in the clearance of extracellular pathogens.
However, this concept has been overstepped with the discoveries of their capacity to live much longer than expected and to produce several key cytokines, chemokines and Fc receptors [124].
Nowadays, evidence has demonstrated important contributions of neutrophils both in the promotion and inhibition of tumor development and growth [125]. This dichotomy suggests that neutrophils are composed of diverse cellular subsets. However, the identification of these subgroups has been trivial due to the lack of robust surface cell markers [125], limiting their characterization of physical properties and functionality [126].
Neutrophils are thought to support tumor growth and progression. Higher neutrophils to lymphocytes ratio represent a poor prognostic marker for a variety of cancer types such as lung and breast cancer [126].
Indeed, neutrophils can support metastatic colonization in a mouse model of breast cancer, orchestrating the formation of the premetastatic niche in the lung [127]. Neutrophils are recruited in the tumor microenvironment by a series of attracting CXC cytokines released by cancer cells [124].
Furthermore, neutrophils are able to sustain tumor progression through the release of proangiogenic factors such as VEGF and MMP9 [128,129] and, by maintaining an immunosuppressive tumor microenvironment through the expression of the arginine-consuming enzyme Arg1 and ROS production [130].
Arginine depletion and ROS directly inhibit T cell antitumor activity and, therefore, neutrophils can negatively impact the efficacy of T-cell-based cancer immunotherapy (Figure 2).
As mentioned above, neutrophils can also exert cytotoxic activity against tumor cells. During the first phases of tumor formation when the immunosuppressive environment is not yet completely established [125] neutrophils recognize and kill cancer cells through ADCC.
Neutrophils contribute to tumor control and tumor eradication also by regulating the activation and function of adaptive immune cells like CD4 or CD8 T cells [124,131]. Indeed, neutrophils also function as APC and thus can directly interact with T cells, supporting a robust antitumor T cell activity. [132–136].
Conversely, neutrophils can recruit and activate Treg cells following the secretion of cytokines like CCL17 and promoting an immunosuppressive environment (Figure 2) [137]. However, in the tumor microenvironment neutrophils, as well macrophages, exert generally a tolerogenic activity and consequently hampering the effectiveness of T-cell-based immunotherapy.
In this context, TGFβ can polarize neutrophils toward an immunosuppressive phenotype and hinder CD8 T cell tumor infiltration [133]. Blocking TGFβ in the tumor boosts CD8 T cell infiltration and CD8 T cell-dependent antitumoral response [133].
On the contrary, IFNβ has been shown to promote neutrophil antitumor activity and recruitment in the tumor microenvironment [125,138].
Blocking the immunosuppressive function of tumor-infiltrating neutrophils has shown to improve the effectiveness of T-cell-based immunotherapies, such as ICB and ACT.
In a murine model of melanoma, the inhibition of the receptor tyrosine kinase c-MET (HGF-R, hepatocyte growth factor receptor) reduces neutrophil recruitment in the tumor microenvironment and in the draining lymph nodes in response to immunotherapy (Figure 2). Consequently, c-MET inhibition enhances the response to ICB and ACT [139].
Meanwhile, the inhibition of the fatty acid transporter protein 2 (FATP2) abrogate neutrophil immunosuppressive activity and response to ICB [140]. In addition, under certain stress conditions, neutrophils can release DNA-based structures, named neutrophil extracellular traps (NETs), which enable neutrophils to capture and kill extracellular pathogens [141].
This cell death phenomenon is nowadays called NETosis [142] and it represents a mechanism adopted by neutrophils to modulate adoptive T cell immunity. NETs are able to interact with DCs enhancing their maturation [143,144], resulting in the upregulation of costimulatory molecules such as CD40, CD80/CD86 and MHCII and proliferation of CD4 T cells [145].
Furthermore, NETs can directly prime T cells leading to increased expression of activation markers such as CD69 and CD25 [146]. Notably, T cells primed by NETs were able to respond to specific antigens even in the absence of costimulatory signals.
It is worth noting that this scenario is common in the tumor microenvironment, and, therefore, NETosis can be exploited to induce or to modulate the activation of tumor-specific T cells. However, neutrophilia and NETs formation support disease progression and metastasis formation in lung and colon cancer, and blocking NETosis resulted in decreased cancer cell dissemination [147]).
Furthermore, it has been shown that NETs can protect cancer cells against T cells and NK cell-mediated cytotoxicity [148]. Tumors cells are able to recruit neutrophils and to sustain NETosis through the secretion of neutrophils chemoattractants, like ligands of the chemokine receptors CXCR1 and CXCR2 [149].
NETs coated tumor cells were physically protected against adaptive immune cells, impairing T cells and NK cells motility and blocking them from reaching the tumor. Notably, pharmacological inhibition of NETosis sensitizes tumors to ICB therapy [149].
This evidence highlights the potential therapeutic opportunities in targeting neutrophils combined with immunotherapies. However, neutrophil contribution to tumor development and progression still needs to be fully elucidated [124] and neutrophil-targeting therapies capable of boosting T-cell-based immunotherapy have not yet reached clinical applications.
Eosinophils participate in tissue repair processes and in the immune response against parasites. Moreover, eosinophils play a role in several diseases like allergic asthma, autoimmune disorders and cancer [150,151].
Although eosinophils were described in tumor samples at the end of the 19th century and are known to infiltrate several tumor types [152], their function in tumor progression and in the tumor microenvironment is still subject of investigation [153].
Studies have shown increased eosinophil count upon ICB therapy in melanoma, prostate and lung cancer patients which positively correlated with the disease prognosis [154–157]. The release of DAMPS by necrotic cells in the tumor stroma is proposed to be responsible for eosinophil recruitment (eosinophilia) [158].
Indeed, eosinophil infiltration in the tumor microenvironment of melanoma patients treated with ICB positively correlates with increased TILs infiltration and better patient outcome [159].
Activated eosinophils are able to remodel and normalize tumor vasculature, promoting T cell infiltration and boosting ACT efficacy [160]. In addition, eosinophils can support antitumor immunity modulating macrophage polarization. Notably, proinflammatory cytokines such as INFγ and TNFα are produced by activated eosinophils.
Therefore, when transferred together with T cells, activated eosinophils promote the polarization of TAM toward the M1-like phenotype, contributing to the establishment of a proinflammatory tumor microenvironment [160].
Due to their tumor-homing properties, eosinophils are becoming novel potential targets to improve T-cell-based immunotherapies. In the preclinical mouse model of hepatic, prostate and breast cancer, inhibition of the dipeptidyl-peptidase 4 (DPP4) with the FDA approved drug sitagliptin enhances host eosinophil-mediated antitumoral activity and contribute to ICB efficacy [161].
These promising studies can pave the way for new therapeutic strategies aimed to enhance T-cell-based immunotherapy through DPP4 inhibition (Figure 2) [162].

Reference :
123. Mestas, J.; Hughes, C.C. Of Mice and Not Men: Differences between Mouse and Human Immunology.
J. Immunol. 2004, 172, 2731–2738.
124. Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the Activation and Regulation of Innate and Adaptive Immunity. Nat. Rev. Immunol. 2011, 11, 519–531.
125. Granot, Z. Neutrophils as a Therapeutic Target. in Cancer. Front. Immunol. 2019, 10, 1710.
126. Sagiv, J.Y.; Michaeli, J.; Assi, S.; Mishalian, I.; Kisos, H.; Levy, L.; Damti, P.; Lumbroso, D.; Polyansky, L.; Sionov, R.V.; et al. Phenotypic Diversity and Plasticity in Circulating Neutrophil Subpopulations in Cancer. Cell Rep. 2015, 10, 562–573.
127. Wculek, S.K.; Malanchi, I. Neutrophils Support Lung Colonization of Metastasis-Initiating Breast Cancer Cells. Nature 2015, 528, 413–417.
128. Scapini, P.; Morini, M.; Tecchio, C.; Minghelli, S.; Di Carlo, E.; Tanghetti, E.; Albini, A.; Lowell, C.; Berton, G.; Noonan, D.M.; et al. CXCL1/Macrophage Inflammatory Protein-2-Induced Angiogenesis In Vivo is Mediated by Neutrophil-Derived Vascular Endothelial Growth Factor-A. J. Immunol. 2004, 172, 5034–5040.
129. Nozawa, H.; Chiu, C.; Hanahan, D. Infiltrating Neutrophils Mediate the Initial Angiogenic Switch in a Mouse Model of Multistage Carcinogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 12493–12498.
130. Schmielau, J.; Finn, O.J. Activated Granulocytes and Granulocyte-Derived Hydrogen Peroxide are the Underlying Mechanism of Suppression of T-Cell Function in Advanced Cancer Patients. Cancer Res. 2001, 61, 4756–4760.
131. Nicolas-Avila, J.A.; Adrover, J.M.; Hidalgo, A. Neutrophils in Homeostasis, Immunity, and Cancer. Immunity 2017, 46, 15–28.
132. Ashtekar, A.R.; Saha, B. Poly’s Plea: Membership to the Club of APCs. Trends Immunol. 2003, 24, 485–490.
133. Fridlender, Z.G.; Sun, J.; Kim, S.; Kapoor, V.; Cheng, G.; Ling, L.; Worthen, G.S.; Albelda, S.M. Polarization of tumor-associated neutrophil phenotype by TGF-beta: “N1” versus “N2” TAN. Cancer Cell 2009, 16, 183–194.
134. Eruslanov, E.B.; Bhojnagarwala, P.S.; Quatromoni, J.G.; Stephen, T.L.; Ranganathan, A.; Deshpande, C.; Akimova, T.; Vachani, A.; Litzky, L.; Hancock, W.W.; et al. Tumor-Associated Neutrophils Stimulate T Cell Responses In Early-Stage Human Lung Cancer. J. Clin. Investig. 2014, 124, 5466–5480.
135. Radsak, M.; Iking-Konert, C.; Stegmaier, S.; Andrassy, K.; Hänsch, G.M. Polymorphonuclear Neutrophils as Accessory Cells for T-Cell Activation: Major Histocompatibility Complex Class II Restricted Antigen-Dependent Induction of T-Cell Proliferation. Immunology 2000, 101, 521–530.
136. Stoppacciaro, A.; Melani, C.; Parenza, M.; Mastracchio, A.; Bassi, C.; Baroni, C.; Parmiani, G.; Colombo, M.P. Regression of an Established Tumor Genetically Modified to Release Granulocyte Colony-Stimulating Factor Requires Granulocyte-T Cell Cooperation and T Cell-Produced Interferon Gamma. J. Exp. Med. 1993, 178, 151–161.
137. Mishalian, I.; Bayuh, R.; Eruslanov, E.; Michaeli, J.; Levy, L.; Zolotarov, L.; Singhal, S.; Albelda, S.M.; Granot, Z.; Fridlender, Z.G. Neutrophils recruit regulatory T-cells into tumors via secretion of CCL17–a new mechanism of impaired antitumor immunity. Int. J. Cancer 2014, 135, 1178–1186.
138. Eash, K.J.; Greenbaum, A.M.; Gopalan, P.K.; Link, D.C. CXCR2 and CXCR4 Antagonistically Regulate Neutrophil Trafficking from Murine Bone Marrow. J. Clin. Investig. 2010, 120, 2423–2431.
139. Glodde, N.; Bald, T.; van den Boorn-Konijnenberg, D.; Nakamura, K.; O’Donnell, J.S.; Szczepanski, S.; Brandes, M.; Eickhoff, S.; Das, I.; Shridhar, N.; et al. Reactive Neutrophil Responses Dependent on the Receptor Tyrosine Kinase c-MET Limit Cancer Immunotherapy. Immunity 2017, 47, 789–802.e9.
140. Veglia, F.; Tyurin, V.A.; Blasi, M.; De Leo, A.; Kossenkov, A.V.; Donthireddy, L.; To, T.K.; Schug, Z.; Basu, S.; Wang, F.; et al. Fatty Acid Transport Protein 2 Reprograms Neutrophils in Cancer. Nature 2019, 569, 73–78.
141. Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535.
142. Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel Cell Death Program Leads to Neutrophil Extracellular Traps. J. Cell Biol. 2007, 176, 231–241.
143. Sangaletti, S.; Tripodo, C.; Chiodoni, C.; Guarnotta, C.; Cappetti, B.; Casalini, P.; Piconese, S.; Parenza, M.; Guiducci, C.; Vitali, C.; et al. Neutrophil Extracellular Traps Mediate Transfer of Cytoplasmic Neutrophil Antigens to Myeloid Dendritic Cells toward ANCA Induction and Associated Autoimmunity. Blood 2012, 120, 3007–3018.
144. Papadaki, G.; Kambas, K.; Choulaki, C.; Vlachou, K.; Drakos, E.; Bertsias, G.; Ritis, K.; Boumpas, D.T.; Thompson, P.R.; Verginis, P.; et al. Neutrophil Extracellular Traps Exacerbate Th1-Mediated Autoimmune Responses in Rheumatoid Arthritis by Promoting DC Maturation. Eur. J. Immunol. 2016, 46, 2542–2554.
145. Scozzi, D.; Wang, X.; Liao, F.; Liu, Z.; Zhu, J.; Pugh, K.; Ibrahim, M.; Hsiao, H.-M.; Miller, M.J.; Yizhan, G.; et al. Neutrophil Extracellular Trap Fragments Stimulate Innate Immune Responses that Prevent Lung Transplant Tolerance. Am. J. Transpl. 2019, 19, 1011–1023.
146. Tillack, K.; Breiden, P.; Martin, R.; Sospedra, M. T Lymphocyte Priming by Neutrophil Extracellular Traps Links Innate and Adaptive Immune Responses. J. Immunol. 2012, 188, 3150–3159.
147. Rayes, R.F.; Mouhanna, J.G.; Nicolau, I.; Bourdeau, F.; Giannias, B.; Rousseau, S.; Quail, D.; Walsh, L.; Sangwan, V.; Bertos, N.; et al. Primary Tumors Induce Neutrophil Extracellular Traps with Targetable Metastasis Promoting Effects. JCI Insight 2019, 4, e128008.
148. Flemming, A. Tumours Use NETs as Physical Shields. Nat. Rev. Immunol. 2020, 20, 352–353.
149. Teijeira, A.; Garasa, S.; Gato, M.; Alfaro, C.; Migueliz, I.; Cirella, A.; de Andrea, C.; Ochoa, M.C.; Otano, I.; Etxeberria, I.; et al. CXCR1 and CXCR2 Chemokine Receptor Agonists Produced by Tumors Induce Neutrophil Extracellular Traps that Interfere with Immune Cytotoxicity. Immunity 2020, 52, 856–871.e8.
150. Melo, R.C.N.; Liu, L.; Xenakis, J.J.; Spencer, L.A. Eosinophil-Derived Cytokines in Health and Disease: Unraveling Novel Mechanisms of Selective Secretion. Allergy 2013, 68, 274–284.
151. Rosenberg, H.F.; Dyer, K.D.; Foster, P.S. Eosinophils: Changing Perspectives in Health and Disease. Nat. Rev. Immunol. 2013, 13, 9–22.
152. Davis, B.P.; Rothenberg, M.E. Eosinophils and cancer. Cancer Immunol. Res. 2014, 2, 1–8.
153. Reichman, H.; Karo-Atar, D.; Munitz, A. Emerging Roles for Eosinophils in the Tumor Microenvironment. Trends Cancer 2016, 2, 664–675.
154. Delyon, J.; Mateus, C.; Lefeuvre, D.; Lanoy, E.; Zitvogel, L.; Chaput, N.; Roy, S.; Eggermont, A.M.; Routier, E.; Robert, C. Experience in Daily Practice with Ipilimumab for the Treatment of Patients with Metastatic Melanoma: An Early Increase in Lymphocyte and Eosinophil Counts is Associated with Improved Survival. Ann. Oncol. 2013, 24, 1697–1703.
155. Gebhardt, C.; Sevko, A.; Jiang, H.; Lichtenberger, R.; Reith, M.; Tarnanidis, K.; Holland-Letz, T.; Umansky, L.; Beckhove, P.; Sucker, A.; et al. Myeloid Cells and Related Chronic Inflammatory Factors as Novel Predictive Markers in Melanoma Treatment with Ipilimumab. Clin. Cancer Res. 2015, 21, 5453–5459.
156. McNeel, D.G.; Gardner, T.A.; Higano, C.S.; Kantoff, P.W.; Small, E.J.; Wener, M.H.; Sims, R.B.; DeVries, T.;Sheikh, N.A.; Dreicer, R. A Transient Increase in Eosinophils is Associated with Prolonged Survival in Men with Metastatic Castration-Resistant Prostate Cancer Who Receive Sipuleucel-T. Cancer Immunol. Res. 2014, 2, 988–999.
157. Tanizaki, J.; Haratani, K.; Hayashi, H.; Chiba, Y.; Nakamura, Y.; Yonesaka, K.; Kudo, K.; Kaneda, H.; Hasegawa, Y.; Tanaka, K.; et al. Peripheral Blood Biomarkers Associated with Clinical Outcome in Non-Small Cell Lung Cancer Patients Treated with Nivolumab. J. Thorac. Oncol. 2018, 13, 97–105.
158. Lotfi, R.; Herzog, G.I.; DeMarco, R.A.; Beer-Stolz, D.; Lee, J.J.; Rubartelli, A.; Schrezenmeier, H.; Lotze, M.T. Eosinophils Oxidize Damage-Associated Molecular Pattern Molecules Derived from Stressed Cells. J. Immunol. 2009, 183, 5023–5031.
159. Simon, S.C.S.; Hu, X.; Panten, J.; Grees, M.; Renders, S.; Thomas, D.; Weber, R.; Schulze, T.J.; Utikal, J.; Umansky, V. Eosinophil accumulation predicts response to melanoma treatment with immune checkpoint inhibitors. Oncoimmunology 2020, 9, 1727116.
160. Carretero, R.; Sektioglu, I.M.; Garbi, N.; Salgado, O.C.; Beckhove, P.; Hämmerling, G.J. Eosinophils Orchestrate Cancer Rejection by Normalizing Tumor Vessels and Enhancing Infiltration of CD8(+) T Cells. Nat. Immunol. 2015, 16, 609–617.
161. Hollande, C.; Boussier, J.; Ziai, J.; Nozawa, T.; Bondet, V.; Phung, W.; Lu, B.; Duffy, D.; Paradis, V.; Mallet, V.; et al. Inhibition of The Dipeptidyl Peptidase DPP4 (CD26) Reveals IL-33-Dependent Eosinophil-Mediated Control of Tumor Growth. Nat. Immunol. 2019, 20, 257–264.
162. Munitz, A.; Hogan, S.P. Alarming Eosinophils to Combat Tumors. Nat. Immunol. 2019, 20, 250–252.
More information: Brigger, D., Riether, C., van Brummelen, R. et al. Eosinophils regulate adipose tissue inflammation and sustain physical and immunological fitness in old age. Nat Metab (2020). doi.org/10.1038/s42255-020-0228-3



{kind=link}