











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Alcune delle domande fondamentali nell’evoluzione rimangono senza risposta, come quando e come i gruppi estremamente diversi di animali – ad esempio i rettili – si sono evoluti per la prima volta.
Per settantacinque anni, le radiazioni adattative – l’evoluzione relativamente rapida di molte specie da un singolo antenato comune – sono state considerate la causa principale della diversità biologica, comprese le origini dei principali piani corporei (caratteristiche strutturali e di sviluppo che identificano un gruppo di animali) e nuovi lignaggi.
Tuttavia, la ricerca passata che esaminava questi rapidi tassi di evoluzione era in gran parte limitata dai metodi utilizzati e dalla quantità di dati disponibili.
In un articolo pubblicato oggi su Nature Communications, un gruppo di ricerca guidato dall’Università di Harvard ha esaminato il più grande insieme di dati disponibili dei principali gruppi di rettili viventi ed estinti (come rettili marini, tartarughe, lucertole e gli antenati di dinosauri e coccodrilli) per affrontare domanda di lunga data di come le radiazioni adattative hanno modellato l’evoluzione dei rettili.
Utilizzando le informazioni sul DNA di specie moderne e centinaia di caratteristiche anatomiche di specie moderne e fossili per l’analisi statistica, lo studio ha rilevato che i periodi di rapido cambiamento anatomico durante l’origine dei gruppi di rettili spesso precedono quando questi gruppi si sono diversificati in centinaia o migliaia di specie. Ciò contraddice le idee di lunga data di radiazione adattativa nella biologia dell’evoluzione.

“I nostri risultati suggeriscono che l’origine dei principali gruppi di rettili, sia vivi che estinti, è stata contrassegnata da tassi molto rapidi di cambiamento anatomico, ma che alti tassi di evoluzione non si allineano necessariamente con la diversificazione tassonomica”, ha affermato il primo autore Dr. Tiago Simões, Membro post-dottorato nel laboratorio di Stephanie Pierce, Professore associato presso il Dipartimento di Biologia Organistica ed Evolutiva dell’Università di Harvard.
Simões e Pierce hanno rivelato che i tassi di evoluzione e la varietà morfologica nei rettili prima dell’estinzione di massa Permiano-Triassico – la più grande estinzione di massa di tutti i tempi – erano ugualmente alti, o persino più alti, che dopo l’evento.
Poiché la diversità delle specie di rettili era molto più bassa durante il Permiano rispetto al Triassico, questi risultati indicano che i rapidi tassi di evoluzione non devono coincidere con la rapida diversificazione tassonomica, come previsto dalla teoria classica della radiazione adattativa. I due possono essere disaccoppiati.
Il team, che includeva anche il dottorato di ricerca. la studentessa Oksana Vernygora e il professor Michael Caldwell dell’Università di Alberta, hanno inoltre scoperto che i tassi di evoluzione accelerati corrispondono all’origine di piani corporei dei rettili unici, ma che adattamenti funzionali molto simili nei rettili possono derivare da tassi di evoluzione variabili.
“Sorprendentemente”, ha detto Pierce, “rettili che hanno evoluto armature protettive simili come tartarughe o corpi serpentini come serpenti, mostrano tassi di evoluzione radicalmente diversi, indicando che l’origine e l’evoluzione di piani corporei unici è eterogenea attraverso l’evoluzione.”
“I nostri risultati mostrano anche che l’origine dei serpenti è caratterizzata dai tassi più rapidi di cambiamento anatomico nella storia dell’evoluzione dei rettili “, ha affermato Simões. “Ma, ciò non coincide con aumenti della diversità tassonomica [come previsto dalle radiazioni adattative] o alti tassi di evoluzione molecolare.”
La discrepanza tra evoluzione morfologica e molecolare supporta l’idea che le sequenze di DNA che codificano proteine non sembrano essere correlate a cambiamenti su larga scala dell’anatomia.
Sebbene siano necessarie molte più ricerche per capire come evolvono i piani corporei, il team ipotizza che le regioni codificanti non proteiche del genoma possano essere responsabili di rapidi cambiamenti morfologici, poiché queste parti sono più libere di mutare e assumere nuovi ruoli funzionali.
“È chiaro per noi che per far avanzare la nostra comprensione dei principali modelli in evoluzione abbiamo bisogno di ulteriori studi in grado di misurare i tassi evolutivi fenotipici e molecolari, i tempi di origine e la diversità fenotipica attraverso grandi scale temporali”, ha affermato Simões.
Simões e colleghi continuano a sviluppare nuovi metodi e stanno espandendo i loro dati indietro nel tempo per esaminare le origini degli amnioti, il gruppo che comprende sia rettili che mammiferi.
Di particolare interesse è individuare quando nel tempo geologico questi due gruppi di animali divergevano e in che modo estinzione, diversificazione e adattamento hanno modellato la loro storia evolutiva negli ultimi 300+ milioni di anni.
“Sono entusiasta di continuare le mie ricerche per svelare le prime dinamiche evolutive dei due gruppi di animali di maggior successo sul pianeta”, ha affermato Simões. “Mi sto anche concentrando sul miglioramento dei protocolli disponibili per analizzare i dati morfologici e costruire alberi evolutivi più robusti, compreso il momento di origine dei principali lignaggi dei vertebrati”.
Evoluzione dei rettili
I rettili formano un enorme gruppo di fossili e vertebrati viventi, di dimensioni variabili da piccoli serpenti a filo a dinosauri sauropodi, che sono gli animali più grandi mai vissuti sulla terra.
Nel tempo i rettili si sono evoluti in forme uniche, come tartarughe, serpenti e dinosauri, ma hanno anche assunto l’aspetto e le abitudini di altri gruppi di vertebrati, come squali e delfini. Come con altre classi di animali, i gruppi di rettili che si ritiene condividano un antenato comune sono noti come lame.
L’applicazione della cladistica ha cambiato le idee su come classificare gli organismi. Ad esempio, poiché si pensa che discendano da piccoli dinosauri bipedi, gli uccelli sono ora inclusi nei rettili. Ma i sinapsidi (una volta chiamati rettili simili a mammiferi) sono classificati con i mammiferi.
Poiché così tanti animali diversi sono inclusi nel termine rettili, sono difficili da definire come un singolo gruppo. I rettili sono amnioti, cioè sono tetrapodi (vertebrati a quattro zampe) con un amnione che circonda e protegge l’embrione in via di sviluppo.
I rettili diversi dagli uccelli e dai loro antenati mancano di piume vere e tutti mancano di capelli veri. Le caratteristiche comuni (sebbene variabili) tra i rettili includono il fatto che non possono regolare la loro temperatura
internamente (con le possibili eccezioni di alcuni dinosauri e tutti gli uccelli), che hanno una vasta copertura di squame o placche ossee o entrambi (eccezioni individuali si verificano in molti gruppi principali), che hanno un cuore a tre camere (con le eccezioni di coccodrilli e, possibilmente, altri arcosauri) e che hanno 12 coppie di nervi cranici.
I principali gruppi di rettili considerati qui sono Anapsida (“rettili a stelo”, tartarughe e altri gruppi primitivi), Euryapsida (notosauri marini, plesiosauri, placodonts e ittiosauri) e Diapsida.
L’ultimo gruppo comprende i lepidosauria (sfenodontiani, come la tuatara e i suoi parenti fossili; lucertole; e serpenti) e gli Archosauria (pseudosuchiani; coccodrilli; pterosauri, noti anche come “rettili volanti” e dinosauri).
Ciascuno dei tre principali gruppi di rettili è definito sulla base del numero e della posizione di grandi aperture nella regione temporale del cranio dietro gli occhi (rispetto ad altre ossa del cranio).
I rettili anapsidi non hanno grandi aperture nella regione temporale del cranio e sono stati il primo gambo a ramificare il lignaggio dei rettili. I rettili euryapsid hanno un’unica apertura temporale nella parte superiore del cranio. I rettili diapsidi hanno due grandi aperture temporali, una sopra e una sotto un ponte osseo orizzontale.
Anapsida
I primi rettili sono noti dai primi Pennsylvanian (323–317 milioni di anni fa, o mya). Erano piuttosto piccoli e lucenti in apparenza, e i loro teschi, mascelle e strutture dentali indicano fortemente che erano insettivori.
In effetti, si pensa che si siano evoluti in tandem con gruppi di insetti che stavano iniziando a colonizzare la terra. Alcuni anfibi Pennsylvanian (323–290 mya) del gruppo dei microsauri si sono evoluti in insettivori che erano così superficialmente simili ai primi rettili che, per un certo periodo, sono stati classificati come tali.
L’uovo amniote si è evoluto nei primi rettili. Ciò consentì la prima vera occupazione della terra da parte dei tetrapodi, poiché l’uovo di amniote permise all’embrione di svilupparsi in un microcosmo acquatico fino a quando non fu pronto per la vita terrestre; questo ha spianato la strada all’enorme radiazione adattativa che alla fine ha avuto luogo tra i rettili.
Robert Carroll del Red-path Museum di Montreal, in Canada, ha sottolineato che i primi rettili probabilmente occupavano la terra prima che l’uovo di amniote fosse completamente sviluppato. Un’analogia può essere trovata in alcune moderne salamandre (piccoli anfibi un po ‘lucertola) che depongono minuscole uova non amniote in luoghi umidi terrestri, come sotto tronchi o in pile di foglie umide.
Queste uova si schiudono in minuscole repliche degli adulti anziché attraversare uno stadio larvale, come accade nelle rane e in altre salamandre. L’evoluzione dell’uovo amniote ha avuto luogo quando le membrane all’interno delle uova dei primi rettili sono state riorganizzate sotto forma di varie sacche e rivestimenti e la membrana esterna ha incorporato il calcio nella sua struttura per formare un guscio.
La conchiglia calcarea (calcarea) offriva protezione all’embrione in via di sviluppo ed era abbastanza porosa da consentire l’ingresso e l’uscita di elementi essenziali, come ossigeno e anidride carbonica.
Il cibo per l’embrione in via di sviluppo è stato fornito sotto forma di un sacco extra-embrionale pieno di tuorlo e un sistema di vasi sanguigni che ha permesso il trasferimento del tuorlo nell’embrione.
L’amnione stesso formava una sacca che conteneva un fluido all’interno del quale era sospeso l’embrione. Ciò ha fornito un “laghetto privato” (un termine usato dal defunto scienziato dell’Università di Harvard Alfred S. Romer, il leader mondiale nella paleontologia dei vertebrati dalla fine degli anni ’40 fino alla sua morte nel 1973) per il suo occupante e ha impedito alle fragili parti embrionali di aderire .
Un’unità composta da una parte dell’allantoide e dal corion membranoso accanto al guscio consentiva l’assorbimento di ossigeno e l’escrezione di anidride carbonica. Una sacca formata dall’allantoide immagazzinava i rifiuti azotati escreti dall’embrione. L’origine dell’uovo amniote fu uno degli eventi evolutivi più importanti che si siano mai verificati.
Passando ad altri rettili anapsidi, le tartarughe furono uno dei primi rettili a ramificarsi dal gambo dell’amnioto. Le tartarughe sono note per la prima volta dal tardo Triassico (227-206 mya) ma probabilmente si sono evolute nel Permiano (290-248 mya).
A parte la protezione, il guscio di tartaruga (che è essenzialmente una porzione del suo scheletro capovolto) ha altre funzioni critiche. Una grande percentuale dei globuli rossi della maggior parte dei vertebrati terrestri si forma nel midollo delle ossa lunghe.
Le tartarughe, tuttavia, hanno bisogno di zampe robuste per sostenere i loro gusci; quindi, le loro ossa degli arti sono molto dense, con poco spazio, se del caso, per il midollo che produce globuli rossi.
È stato dimostrato che il guscio di tartaruga è pieno di canali e cavità in cui i globuli rossi sono prodotti in quantità. Inoltre, anziché essere solo uno scudo inerte, il guscio di tartaruga è il sito del metabolismo del calcio ed è importante nel processo di regolazione della temperatura, assorbendo il calore durante il processo di crogiolarsi (prendere il sole).
Altri attributi delle tartarughe includono la capacità di alcuni di essi di assorbire ossigeno nell’acqua attraverso chiazze di pelle sottile sul corpo, nel rivestimento della bocca o all’interno della cloaca, un’estensione terminale della parete intestinale.
Alcune tartarughe possono congelare solido in inverno e scongelarsi in primavera senza effetti dannosi, un processo che si svolge anche in varie specie di rane. La capacità delle tartarughe di sopravvivere a gravi lesioni è ben nota e molte specie possono esistere in assenza di ossigeno per lunghi periodi di tempo.
Le tartarughe di cuoio hanno un flusso sanguigno controcorrente corrente simile a quello dei mammiferi di acque profonde e possono immergersi nel mare a grandi profondità e rimanere attive
a temperature molto fredde. La temperatura di incubazione delle uova di molte specie di tartaruga determina il sesso dei boccaporti. In alcuni casi i “nidi freddi” producono femmine e i “nidi caldi” producono maschi, ma può verificarsi anche il contrario. Le uova di tartaruga hanno grandi quantità di tuorlo rispetto a quelle di molti altri vertebrati. Questo “cibo per uova” sostiene i giovani durante lunghi periodi di incubazione.
L’origine delle tartarughe è in qualche modo in dubbio. Un antico anapside con costole espanse, Eunotosaurus, una volta fu proposto come forma ancestrale, ma il cranio di Eunotosaurus non era simile alla tartaruga. D’altra parte, lo scheletro del corpo dei primi rettili chiamati procolofonidi aveva un guscio e il cranio era un po ‘tortuoso.
Owenetta, un procolofonide trovato nell’alto permiano (256-248 mya) del Sud Africa ha nove personaggi avanzati nel cranio e uno nell’omero che sono condivisi con Proganochelys, un’indiscutibile tartaruga triassica (248-206 mya). Owenetta manca di una conchiglia, tuttavia; così è stato suggerito che i cambiamenti del cranio negli antenati delle tartarughe hanno preceduto quelli che hanno portato all’origine del guscio.
Le vere tartarughe (ordine Testudine) sono composte da tre sottordini principali: i proganochelydians, i pleurodirans e i criptodiran.
I proganochelydians sono i più primitivi e sono noti dal tardo Triassico al primo Giurassico (206-180 mya). La conchiglia è simile a quella delle tartarughe moderne, tranne per il fatto che ha ossa extra e la testa e gli arti non possono essere ritratti efficacemente al suo interno. Al cranio mancano i denti tranne alcuni al palato. I pleurodirani e i criptodirani non hanno denti nel cranio e possono ritrarre la testa, il collo e la coda nel guscio.
Nelle pleurodire il collo oscilla lateralmente quando viene retratto, in modo che la tartaruga guardi fuori con un solo occhio. Nei cryptodires il collo si piega su se stesso quando viene retratto, in modo che la tartaruga guardi con entrambi gli occhi. Stranamente, la differenziazione del collo in queste tartarughe non si è verificata fino al tardo Cretaceo (99–65 mya).
Riflettendo la loro origine nel Gondwana, si trovano solo criptodadi nell’emisfero settentrionale, mentre un numero significativo di pleurodire si verifica nell’emisfero meridionale. Seaturtles e tartarughe dal guscio morbido, animali familiari in gran parte del mondo oggi, sono comparsi nell’alto giurassico (159-144 mya); quando nel tardo Cretaceo nacquero le specializzazioni al collo, tutte le tartarughe erano essenzialmente moderne.
Alcuni grandissimi abitanti vivevano nei mari mesozoici (248-65 mya) durante il periodo in cui i dinosauri dominavano la terra. Archelon of the Cretaceous (144–65 mya) aveva una lunghezza della conchiglia di 6,3 piedi (1,9 m). Ma la più grande tartaruga conosciuta è una tartaruga pleurodire d’acqua dolce (gruppo collo laterale) con una lunghezza della conchiglia di 7,6 piedi (2,3 m). Appropriatamente chiamata Stupendemys, questa tartaruga è stata raccolta durante una gita ad Harvard in Venezuela. L’animale proveniva da sedimenti pliocenici (5,3-1,8 mya), non molto tempo fa in termini di tempo geologico.
Diverse strane lame di rettili si ramificarono dal gambo anapside, tra cui l’elefante, i pareiasauri terrestri e i sottili mesosauri marini. I pareiasauri (tra questi, il noto genere Scutosaurus) sono stati trovati nel Medio e Alto Permiano (269-248 mya) in Africa, Europa occidentale, Russia e Cina. Erano lunghi fino a 3 m (10 piedi) e avevano una posizione eretta (a differenza di quella di altri amnioti dell’epoca) e arti robusti che sostenevano il corpo massiccio.
La testa di parei era bassa e spessa, con ossa fortemente scolpite che proteggevano gli occhi e il cervello minuscolo. I denti avevano corone compresse a forma di foglia come quelle delle moderne lucertole erbivore mangiatori di foglie; quindi, i pareiasauri furono probabilmente i primi grandi erbivori terrestri.
I mesosauri, d’altra parte, erano i primi rettili veramente acquatici. Questi animali gracili erano lunghi circa 0,9 m di lunghezza, circa un terzo costituiti dalla coda. I mesosauri fossili sono conosciuti solo dalle coste adiacenti dell’Africa meridionale e del Sud America orientale, riflettendo il fatto che Gondwana si divise nei due continenti.
Nei mesosauri il muso era molto lungo e la bocca era piena di denti lunghi e simili ad aghi. Apparentemente questi denti formavano un sistema di tensione specializzato che permetteva agli animali di nutrirsi di piccoli crostacei, probabilmente quelli che si trovano negli stessi letti fossili dei mesosauri. La lunga coda compressa probabilmente era usata per la propulsione nel nuoto. Le vertebre della coda posteriore presentano pianure fratturate, indicando che potrebbe essersi verificata un’autonomia caudale (spargimento volontario dell’estremità della coda durante lo stress); questa funzione potrebbe aver permesso loro di sfuggire alla presa dei predatori. Inizialmente si pensava che il cranio dei mesosauri avesse un’apertura temporale, ma questo è stato successivamente smentito.
Euryapsida
Per quanto riguarda i rettili euryapsid, troviamo quattro gruppi altamente adattati di rettili marini, alcuni dei quali hanno raggiunto proporzioni pesanti. Si ritiene che gli euryapsidi si siano evoluti dai diapsidi, avendo perso l’apertura temporale inferiore nel processo.
Erano una parte molto importante dell’ambiente marino durante il Mesozoico e, in un certo senso, erano dominanti in quel contesto quanto i dinosauri erano sulla terra. Un argomento che viene spesso trascurato nel corso delle discussioni sulle ragioni della morte dei dinosauri alla fine del Cretaceo riguarda le ragioni dietro l’estinzione di tutti i rettili marini euryapsid alla fine del Cretaceo.
Nothosaurs visse dal Triassico in tarda età, ma era più comune nel Triassico medio (242–227 mya). Erano relativamente piccoli rispetto ai plesiosauri che li seguivano. I nothosauri avevano il collo e gli arti moderatamente lunghi modificati come pinne. Si pensa che siano stati possibili equivalenti ecologici dei moderni sigilli e lontre. I nothosauri avevano denti affilati e conici modificati per la cattura dei pesci. La struttura della cintura scapolare del notosauro è unica tra i rettili; fornisce poco spazio per l’attacco dei muscoli del corpo che supportano il tronco, come accade nei rettili terrestri.
Si pensa che i plesiosauri si siano evoluti dai non dinosauri nel Triassico medio. Persistettero fino alla fine del Cretaceo, ma furono più abbondanti nel Giurassico medio (180-159 mya) e lentamente diminuirono di numero fino alla loro estinzione.
I plesiosauri avevano le gambe modificate come pagaie che si agitavano su e giù come ali acquatiche. Questa azione non solo li ha spinti attraverso l’acqua, ma ha anche dato loro un passaggio che ha permesso loro di “volare” attraverso il mare allo stesso modo di seaturtles e pinguini.
Le ossa della spalla e le cinture dell’anca dei plesiosauri si espandevano notevolmente sotto, formando un’armatura sul fondo dell’animale che non lasciava “ventre molle” per l’attacco di predatori come gli squali. Due tipi generali di tipi di corpo erano prominenti nei plesiosauri, forme a collo lungo e forme a collo corto.
Nel primo gruppo, una piccola testa era posizionata alla fine di un collo molto lungo. Il corpo era pesante e bulboso. I denti erano conici e affilati; per questo motivo si presume che questi plesiosauri si nutrano principalmente di pesci.
Il clade elasmosauride dei plesiosauri dal collo lungo raggiungeva una lunghezza di 12,2 m (40 piedi) e aveva enormi pale e teste molto piccole. Alcuni ricercatori hanno suggerito che i mostri di Loch Ness (se effettivamente esistevano) erano plesiosauri dal collo lungo. Se questo è vero, dal Mesozoico devono essersi verificati notevoli cambiamenti fisiologici che hanno permesso loro di adattarsi alle acque ghiacciate.
I plesiosauri dal collo corto non hanno praticamente alcun collo e una testa massiccia. È stato suggerito che fossero gli equivalenti ecologici delle orche dei tempi attuali, poiché erano carnivori consumati. Il Kronosaurus, il più grande rettile marino che sia mai vissuto, era un animale enorme che raggiungeva una lunghezza di almeno 42 piedi (12,8 m).
Questo animale è stato trovato nella proprietà di un allevatore in Australia e finì nel Museum of Comparative Zoology di Harvard. Uno dei problemi della raccolta di un animale così grande è come e dove esporlo. Come spesso accade, una stanza del museo ha dovuto essere ristrutturata per mettere in mostra questa “balena assassina rettiliana”.
I Placodont erano rettili marini con corpi corti e robusti che vivevano dal Triassico medio al superiore (242-206 mya). Gli arti erano solo moderatamente a paddle. Una volta è stato suggerito che i segnaposto fossero collegati piuttosto da vicino ai nonosauri, ma non ci sono davvero buone prove a sostegno di questa ipotesi.
La caratteristica più distintiva dei segnaposto è la forma dei loro denti, che sono appiattiti piuttosto che appuntiti. Nel noto genere Placodus, i denti lungo il margine della regione della guancia e al palato sono grandi e molto piatti, mentre i denti anteriori lunghi e stretti sporgono dall’estremità del muso piuttosto ristretto.
Si pensa che Placodus abbia usato i denti anteriori per afferrare i molluschi e quelli posteriori per schiacciarli. Alcuni segnaposto, come Henodus, erano superficialmente come le tartarughe, in quanto avevano un guscio superiore composto da numerose piccole ossa poligonali. Un guscio inferiore non era presente.
I rettili marini più specializzati erano gli ittiosauri, il cui corpo assunse l’aspetto di moderni tonni, squali e focene. Ichthyosaurs visse dai primi tempi del Triassico al Medio Cretaceo (248-112 mya).
Il cranio degli ittiosauri è aerodinamico, con un lungo muso; gli occhi sono molto grandi. Il corpo è a forma di fuso e gli arti sono ridotti a pinne. La pinna caudale è simile a un pesce. Le singole vertebre nel midollo spinale hanno la forma di dischi biconcave molto corti e compatti, molto simili nell’aspetto a quelli dei moderni squali. Si stima che alcuni ittiosauri fossero molto attivi e potessero nuotare tra 30 e 40 mph (48,3-64,4 km / h). Perché gli ittiosauri si siano estinti nel Medio Cretaceo (circa 121–99 mya), molto prima dei dinosauri e di altri rettili marini, rimane un mistero.
Diapsida
Oltre ad avere due aperture nella regione temporale del cranio, i diapsidi hanno in genere gli arti posteriori più lunghi degli arti anteriori. Il più antico diapside conosciuto era un piccolo animale simile a una lucertola con una lunghezza del corpo (meno la lunga coda) di circa 20,3 cm. Questo esile animale, chiamato Petrolacosaurus, fu raccolto dal tardo pennsylvaniano (circa 303–290 mya) del Kansas.
Due lame distinte, la lepidosauria e l’archosauria, si ramificarono presto dal tronco diapside. Questi due gruppi sono caratterizzati da modelli contrastanti di locomozione e postura.
I lepidosauri mantennero la postura tentacolare e il movimento laterale diretto degli arti trovati nei tetrapodi primitivi. L’ondulazione laterale della colonna vertebrale era anche un importante metodo di locomozione per la maggior parte dei lepidosauri, raggiungendo il suo più alto grado di sviluppo nei serpenti.
Un’altra caratteristica importante del lepidosauro era rappresentata da ossa del cranio vagamente separate, che permettevano di ingoiare intere prede. D’altra parte, gli archosauri presentavano un’ondulazione laterale limitata o assente nella colonna vertebrale e gli arti venivano portati in una posizione più direttamente sotto il tronco. Queste modifiche hanno raggiunto il loro massimo grado di sviluppo nei dinosauri e negli pterosauri.
Le specie rappresentative dei tre gruppi della lepidosauria indirizzate qui, sfenodontidi, lucertole e serpenti, sono attualmente vive. Per quanto riguarda gli sfenodontidi, i tuatara (Sphenodon) della Nuova Zelanda sono gli unici membri viventi di questo gruppo un tempo numeroso. Un articolo di giornale risalente agli anni ’40 spiegava che la tuatara assomigliava a una lucertola ma in realtà era un “fossile vivente”.
Esistono diverse differenze tra sfenodontidi e lucertole, che si sono separate l’una dall’altra nel Triassico, probabilmente all’inizio dell’epoca. Negli sfenodontidi, i muscoli della mascella sono massicci, il che consente loro di avere un morso più forte ma più lento delle lucertole. I denti sfenodontidi sono fusi alla mascella così saldamente (condizione acrodont) che la mascella ha un aspetto a dente di sega, come se i denti fossero semplicemente dentellature dell’osso stesso.
I muscoli della mascella sono meno massicci nelle lucertole e si trovano più indietro nella bocca, producendo un morso più debole ma più veloce. La maggior parte delle lucertole ha denti leggermente allentati all’interno della mascella (condizione del pleurodonte) e questi denti sono sostituiti frequentemente nella maggior parte delle specie.
Gli sfenodontidi erano i lepidosauri dominanti del Giurassico (206-144 mya), ma diminuirono drasticamente nel Cretaceo quando le lucertole iniziarono a diversificarsi ampiamente. Solo le due specie di tuatara neozelandese sono sopravvissute fino ad oggi, altre si sono estinte alla fine del Cretaceo.
La tuatara è attiva a temperature molto più basse rispetto alla maggior parte delle lucertole e le uova hanno un periodo di gestazione di circa nove mesi prima di essere deposte. Il periodo di incubazione per le uova è di circa 15 mesi, il più lungo di tutti i rettili viventi conosciuti.
La crescita nei giovani è lenta e gli animali non raggiungono la maturità sessuale fino a quando non hanno circa 20 anni. La crescita lenta continua quindi fino a quando l’animale ha un’età compresa tra 50 e 60 anni. Piuttosto che avere emipeni (doppio pene) come lucertole e serpenti, il tuatara maschio trasferisce lo sperma alla femmina da un’estensione dell’intestino chiamata cloaca. Mentre la maggior parte delle lucertole sembra guardare attraverso una persona, la tuatara ha uno sguardo diretto, con grandi occhi marroni che sembrano più mammiferi che rettiliani.
Lucertole e serpenti sono considerati un singolo clade, lo Squamata (rettili in scala). Sia le lucertole che i serpenti hanno forme senza gambe con una struttura a mascella che consente loro di ingoiare prede intere. I serpenti, tuttavia, hanno portato queste tendenze all’estremo.
Le prime lucertole sono rappresentate da un animale noto come Paliguana, del tardo Permiano del Sud Africa. Il fatto che le lucertole avessero una struttura della mascella più efficace, un migliore udito e una migliore locomozione probabilmente hanno permesso loro di sfruttare gli habitat occupati da altri tetrapodi simili a lucertole, come gli sfenodontidi. La maggior parte delle lucertole moderne, ad eccezione del drago di Komodo (una lucertola che può abbattere cervi), non ha raggiunto grandi dimensioni. Ma nel Cretaceo, le lucertole ancestrali si sono evolute in mosasaur ecologicamente importanti, le lucertole marine che sono cresciute fino a 9,1 m di lunghezza. Una lucertola terrestre gigante del passato era la lucertola monitor Megalania (probabilmente 20 piedi o 6,1 m di lunghezza), il principale predatore nel Pleistocene (1,8-0,1 mya) dell’Australia. Inizialmente si pensava che i fossili di Megalania fossero quelli dei dinosauri, ma Max K.
I serpenti hanno avuto origine molto più tardi nella documentazione fossile rispetto alle lucertole, a volte durante il Cretaceo medio-alto. I quattro fossili che si avvicinano maggiormente all’origine dei serpenti sono Pachyrhachis, Podophis, Lapparentophis e Dinilysia.
I primi tre sono del Cretaceo medio, ma Dinilysia, il più completo e ben studiato dei quattro, è del tardo Cretaceo. Gli squamati marini Pachyrhachis e Podophis sono stati considerati i serpenti più primitivi da alcuni ricercatori, perché la configurazione delle ossa del cranio ricorda quella dei serpenti viventi, ma hanno uno scheletro degli arti posteriori ben sviluppato.
Il serpente terrestre Lapparentophis, spesso chiamato il “serpente più antico”, è rappresentato solo da vertebre, ma sono certamente vertebre di serpenti, con tutte le modifiche uniche che si trovano nei serpenti viventi generalizzati. Dinilysia ha un teschio che è un mosaico di lucertole e personaggi di serpenti primitivi, ma le sue vertebre sono chiaramente simili a quelle di un serpente simile a un boa. Sfortunatamente, l’intero quadro dell’evoluzione dei primi serpenti è stato confuso da argomenti contorti e gargarismi, il problema, come sempre, è la somiglianza di base di serpenti e lucertole.
I serpenti primitivi erano dominanti nel mondo fino al Miocene, quando i serpenti moderni sostituivano rapidamente i tipi meno avanzati. Tre fattori probabilmente hanno giocato un ruolo: il ritorno di climi caldi ed equi alle latitudini più alte a seguito del deterioramento climatico nell’Oligocene (circa 33-23 mya), la notevole diffusione degli habitat delle praterie in tutto il mondo e l’evoluzione di molti gruppi di roditori che potrebbe essere sfruttato dai serpenti come cibo. I più grandi serpenti moderni includono i boa del Nuovo Mondo e i pitoni del Vecchio Mondo. Il gigantesco serpente simile a un pitone Wonambi viveva nel Pleistocene dell’Australia, insieme alla lucertola gigante Megalania.
Archosauria
Gli archosauri, o “rettili dominanti”, si sono ramificati in una serie impressionante di gruppi importanti, tra cui pseudo-tali, coccodrilli, pterosauri e dinosauri. Il clade dell’archosauro può essere definito da due aperture temporali nel cranio, una (fenestra antorbitale) tra l’occhio e la narice e un’altra (fenestra mandibolare) nella parte posteriore della mascella inferiore.
Molti degli altri personaggi dell’archosauro riflettono i cambiamenti scheletrici associati a una postura più eretta e un movimento da davanti a dietro degli arti. Robert Carroll ha sottolineato che i lignaggi all’interno dell’assemblea degli archosauri erano tutti distinti l’uno dall’altro quando apparvero per la prima volta come fossili.
I coccodrilli viventi sono imparentati con la Pseudosuchia, un gruppo eterogeneo di antichi arcosauri a volte simili a coccodrilli che sono collegati ai veri coccodrilli principalmente sulla base dell’avere la cosiddetta struttura normale del coccodrillo del tarso (caviglia).
La Pseudosuchia è una delle due maggiori pale che si ramificarono dai primi Archosauria. Tutti gli pseudosuchiani avevano un’ampia armatura esterna composta da placche ossee. Il gruppo comprende i rauisuchidi, i fitosauri e gli aetosauri.
I rauisuchidi sono rettili triassici medi e tardivi che avevano una posizione più o meno verticale e crescevano fino a 6,1 m di lunghezza. Ticinosuchus, dalla prima parte del Triassico medio svizzero, è una delle forme più conosciute. Nello scheletro degli arti, la caviglia e il piede avanzarono al livello di quelli dei coccodrilli moderni.
Presumibilmente, la posizione eretta si è sviluppata indipendentemente dal lignaggio che ha portato ai dinosauri. Ticinosuchus aveva un’armatura di due file di piccole placche ossute che si estendevano lungo il tronco e un’unica fila nella parte superiore e inferiore della coda. Denti affilati e penetranti incastonati indicano che questo animale era un carnivoro.
I fitosauri si sono verificati abbondantemente nel tardo Triassico del Nord America, dell’India e dell’Europa. Sebbene non fossero veri coccodrilli, li assomigliavano in forma corporea e probabilmente avevano uno stile di vita simile. I fitosauri avevano un muso molto lungo, con denti affilati e penetranti in orbite lungo il margine delle mascelle. Sebbene le aperture nasali fossero in cima alla testa, erano posizionate molto indietro sul muso piuttosto che sulla punta, come nei veri coccodrilli. Sia il tronco che la coda avevano un’ampia copertura di armatura dermica. Una varietà di altri rettili contemporanei sono stati trovati nel contenuto dello stomaco dei fitosauri, documentando così le loro abitudini carnivore. La documentazione fossile di fitosauri è limitata al tardo Triassico.
Gli aetosauri formarono un gruppo distinto noto anche solo dal tardo Triassico. Invece di denti appuntiti, gli aetosauri avevano denti piccoli a forma di foglia che suggeriscono una dieta erbivora. Il noto aetosauro Stagnolepus, era un bizzarro
bestia con zampe corte e posizione eretta. Il corpo era rotondo e la coda massiccia. Il corpo era coperto da grandi placche quadrangolari che formavano un’ampia armatura lungo la schiena, si estendeva lungo i lati, copriva il ventre e circondava la coda.
La testa era piccola rispetto al corpo e ristretta in un breve rostro anteriore ai denti, ricoperto da un’area gonfia che indicava che l’animale aveva il naso di un piccione. La relazione tra gli aetosauri e le altre pale dell’archosauro è poco compresa.
I coccodrilli sono gli unici arcosauri giganti sopravvissuti e i principali predatori in molti ambienti acquatici di tutto il mondo oggi. Un recente sondaggio condotto in Florida ha riportato che un alligatore maschio era lungo 4,3 metri e pesa 429 kg. Coccodrilli molto più lunghi sono stati trovati nei tropici dell’emisfero australe.
Perché i coccodrilli siano sopravvissuti ai cicli di estinzione verificatisi tra gli altri grandi archosauri è una questione di congetture. I primi coccodrilli erano terrestri e includevano forme come Gracilisuchus del Triassico medio del Sud America, che camminava su due zampe, e il Terrestrisuchus a quattro zampe del tardo Triassico d’Europa, che aveva uno scheletro corporeo estremamente gracile e doveva essere un corridore veloce. Terrestrisuchus aveva le dimensioni di un coniglio ma era un carnivoro che probabilmente si affrettava a entrare e uscire dai nascondigli del Triassico in cerca di prede delle dimensioni di un topo.
I mesosuchiani del Giurassico assomigliavano molto più ai moderni coccodrilli che alle forme triassiche. Molti di loro sono diventati semiacquatici e alcuni hanno invaso l’ambiente marino.
Il processo di lasciare la terra per diventare semiacquatico era importante in termini di cambiamenti del corpo avvenuti in queste forme transitorie. Famiglie moderne di coccodrilli, Alligatoridae (alligatori e caimani), Crocodylidae (coccodrilli moderni) e Gavialidae (il gaviale), sono noti per risalire al tardo Cretaceo. Gli alligatoridi hanno un muso largo, con un numero relativamente grande di denti un po ‘spuntati.
I coccodrilli hanno un muso più lungo e più sottile, con un numero significativo di denti appuntiti. I gavialidi hanno un muso allungato e denti aghiformi, così come i garfish; i gavialidi sono mangiatori di pesce consumati. Alcuni coccodrilli cretacei crebbero fino a dimensioni molto grandi e probabilmente predarono i dinosauri. L’imperatore di Sarcosuchus dall’Africa probabilmente raggiunse una lunghezza di 12 m (40 piedi) e alcuni dal Texas erano anche così a lungo.
I coccodrilli moderni erano molto più diffusi nel mondo all’inizio del Terziario (circa 65-34 mya) di quanto lo siano oggi, il loro declino probabilmente derivante dal deterioramento climatico nell’era cenozoica.
I coccodrilli sono gli unici rettili viventi che danno la vera cura dei genitori ai loro piccoli, inclusa la protezione del nido, aiutando i giovani a uscire dalle uova rompendo le uova con le mascelle, portando i giovani dal nido all’acqua in bocca e proteggendoli nel acqua per un po ‘. Si verifica anche la comunicazione vocale tra genitori e giovani.
L’ornitodirano sviluppò un collo che poteva essere piegato a forma di S e un piede stretto e compresso. Un ramo dell’Ornithodira, il Pterosauria (rettili volanti) del Mesozoico, aveva un quarto dito molto lungo e molto allargato sulla mano, che sosteneva una membrana ad ala.
L’altro ramo dell’Ornithodira, che includeva Lagosuchus e i dinosauri, aveva le gambe allungate della parte inferiore (tibiae e metatarsali) e un osso della coscia (femore) con la testa girata verso l’interno in modo da poter entrare in una presa profonda nella cintura dell’anca (cintura pelvica). Questa presa consentiva alle gambe di sostenere la grande cintura dell’anca, che a sua volta sosteneva il corpo.
I pterosauri furono i primi vertebrati a sviluppare un vero volo battente le ali. Sono conosciuti dal tardo Triassico ed erano diventati abbastanza diversi da questo momento. Il fatto che gli pterosauri avessero un forame nel cranio davanti agli occhi (forame antorbitale), gambe disposte principalmente direttamente sotto il corpo e un’articolazione della caviglia simile a un dinosauro indica la loro stretta relazione con Lagosuchus e i dinosauri. Ma il fatto che avessero un quinto osso metatarsale adunco e un lungo quinto dito suggerisce che si sono discostati dai primi arcosauri prima che i dinosauri (che avevano il quinto dito ridotto o perso) si fossero separati.
I pterosauri andavano dalle forme molto piccole alle creature di gran lunga più grandi che siano mai volate. Sono divisi in due gruppi principali, i rhamphorhynchoids primitivi e i pterodattiloidi più avanzati. I rhamphorhyncoids sono noti per la prima volta dal tardo Triassico ed erano dominanti in tutto il Giurassico.
Avevano una faccia corta, un collo corto e una coda lunga. Alcuni rhamphorhyncoids erano piccoli quanto i passeri inglesi. Fin dall’inizio, i rhamphorhyncoids avevano sviluppato vari personaggi specializzati per il volo, tra cui uno sterno (sterno) che aveva una chiglia simile a una barca che sosteneva i muscoli che sbattevano le ali, come negli uccelli. Altre ossa simili a quelle degli uccelli includevano la scapola (scapola), la coracoide (osso della cintura della spalla dritta) e l’omero (osso del braccio superiore), tutti specializzati per contribuire al volo attivo e svolazzante.
I pterodattiloidi apparvero nel tardo giurassico e durarono fino alla fine del Cretaceo. Questi rettili avevano una faccia molto più lunga, un collo più lungo e una coda più corta dei rhamphorhynchoids. Il cranio era altamente modificato nel genere cretaceo Pteranodon, che aveva un lungo rostro (muso) simile a una vescica davanti all’occhio e una proiezione simile a una lama quasi altrettanto lunga dietro l’occhio.
D’altra parte, il Pterodaustro, proveniente dal Cretaceo superiore dell’Argentina, non aveva praticamente alcun teschio dietro l’occhio, ma aveva mascelle grandi, allungate e ricurve verso l’alto. La mascella inferiore aveva denti molto lunghi e ben stretti che si ritiene abbiano teso piccoli animali invertebrati dall’acqua allo stesso modo delle balle nelle balene.
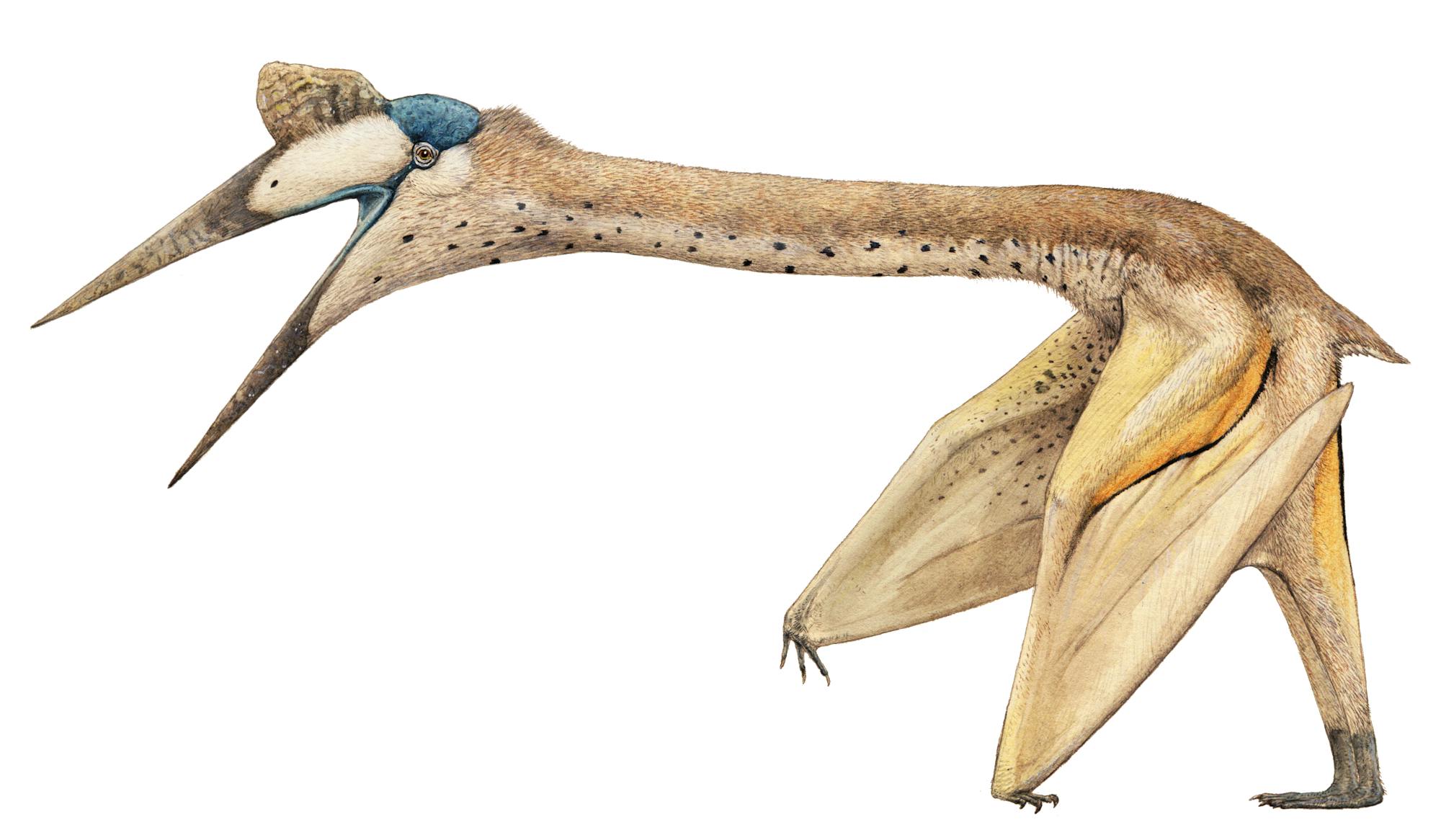
Quetzalcoatlas, dal tardo Cretaceo del Texas, era l’animale più grande di sempre a volare, con un’apertura alare di oltre 35 piedi (10,7 m). Prima della scoperta del Quetzal-coatlas, lo zoologo britannico JZ Young suggerì che Pteranodon, con la sua apertura alare di 7 metri, era probabilmente l’animale più grande che potesse volare. Robert Carroll ha sottolineato che Quetzalcoatlas era ovviamente molto più pesante di Pteranodon.
È stato suggerito che il Quetzalcoatlas avrebbe potuto nutrirsi dei cadaveri dei dinosauri, come un gigantesco avvoltoio. I pterosauri erano molto diversi dagli uccelli, in quanto l’ala era composta da una membrana, qualcosa di simile a quello dei pipistrelli. La differenza è che negli pterosauri la membrana era sostenuta interamente da un quarto dito lungo e robusto. A differenza dei pterosauri, i pipistrelli hanno una membrana alare più ampia e si attacca agli arti posteriori.
Lagosuchus, dal Medio Triassico del Sud America, fornisce un collegamento strutturale tra i primi arcosauri e dinosauri. Lagosuchus era lungo solo circa 1 m (0,3 m) e aveva uno scheletro molto leggero, con arti lunghi e delicati; questo era un umile antenato dei giganteschi animali a venire.
A Lagosuchus gli arti posteriori erano molto più lunghi di quelli anteriori. Inoltre, la tibia (osso della parte inferiore della gamba) era molto più lunga del femore (parte superiore della gamba o osso della coscia). La cintura pelvica (cintura dell’anca) era composta da tre ossa che formavano una struttura triradiata (a tre punte). Pertanto, il lungo ilio era diretto in avanti, il lungo ischio era diretto all’indietro e il pube corto si trovava sopra le altre due ossa della cintura pelvica. Nel tarso (caviglia) c’era una cerniera tra le ossa tarsali superiore e inferiore. Tutte queste caratteristiche (che riflettono i cambiamenti negli arti e nelle cinture degli arti, principalmente la cintura pelvica e gli arti posteriori) hanno reso Lagosuchus il più simile a un dinosauro di uno qualsiasi degli archosauri primitivi.
Dinosauria I
dinosauri affascinano più persone di qualsiasi altro gruppo di rettili. Il raggiungimento di grandi dimensioni e diversità e il lungo dominio della terra da parte dei dinosauri formano gran parte di questo fascino. Come è vero per la grande apparizione grigia nel Don Giovanni di Mozart o nel mostro di Frankenstein, le persone adorano cose terribili e meravigliose allo stesso tempo. In effetti, il nome dinosauro deriva dalle radici che significa “terribile lucertola”.
Caratteristiche evolutive uniche sono evidenti nelle mani di alcuni dinosauri. L’articolazione tra il pollice e il palmo era strutturata in modo tale che quando la mano era chiusa, il pollice si piegava verso il palmo, indicando che la mano veniva utilizzata per afferrare e trattenere gli oggetti. In effetti, l’andatura bipede nei primi arcosauri lascia le mani libere per altre funzioni sia nei pterosauri che nei dinosauri.
Nella cintura pelvica del dinosauro (cintura dell’anca), l’acetabolo (presa dell’anca) era aperto o era composto da cartilagine in forme primitive, probabilmente aumentando la velocità e la gamma del movimento delle gambe dalla parte anteriore a quella posteriore. Inoltre, una o più vertebre sono state incorporate nella cintura pelvica, dandole più forza per una locomozione vigorosa.
Tre pale di dinosauro principali sono riconosciute dalla maggior parte dei paleontologi: i Theropoda, che erano bipedi e principalmente carnivori; il Sauropodomorpha, che aveva il collo lungo ed era erbivoro; e l’ornitischia, che comprendeva un diverso assemblaggio di erbivori a becco.
I primi membri di tutti e tre i gruppi sono conosciuti per la prima volta all’incirca nello stesso periodo nel tardo Triassico. Durante la prima parte del tardo Triassico, i dinosauri erano un importante gruppo emergente, ma in realtà erano dominati da arcosauri pseudosuchiani e sinapsi avanzati (mammiferi ancestrali con caratteristiche rettiliane).
Fu solo dopo la grande estinzione avvenuta alla fine del Triassico che i dinosauri divennero il gruppo dominante di grandi vertebrati terrestri. Questo dominio è durato fino a quando i dinosauri si sono estinti alla fine del Cretaceo.
I teropodi avevano personaggi di dinosauri più primitivi rispetto agli altri due gruppi. Ad esempio, i teropodi conservavano i loro denti simili a lama; da questa caratteristica si sa che erano carnivori. Tutti i teropodi conosciuti erano bipedi e molti mantenevano le mani afferranti.
I teropodi più primitivi conosciuti erano gli herrerasauridi del tardo Triassico di Argentina, Brasile e Nord America. Una specializzazione che questi animali condividevano con i successivi teropodi era un’articolazione nella mascella inferiore tra le porzioni portanti e non portanti. Questa innovazione probabilmente ha contrastato lo stress associato al morso di prede relativamente grandi.
Le due pale principali di Theropoda sono la Neotheropoda e la Celurosauria. Un neotopode precoce, la coelofisi del tardo triassico, era di corporatura gracile e aveva un nodo nella mascella superiore che si riteneva un adattamento per trattenere una piccola preda. Il Ceratosauro del tardo Giurassico era una forma robusta, con pomelli a forma di corno sulla parte superiore del cranio.
Lo Spinosauro del Cretaceo aveva un muso molto lungo e grandi denti conici (piuttosto che a lama); i denti alla fine del muso erano più grandi degli altri. È stato suggerito che Spinosaurus ha mangiato grandi pesci, perché alcune località fossili in cui sono stati raccolti gli spinosauri contenevano abbondanti resti di pesci. L’allosauro del giurassico, un enorme carnivoro bipede con la testa compressa, è presente in molti musei e rappresenta un ramo significativo del tronco teropode chiamato carnosauri.

La celurosauria, l’altro ramo principale dello stelo del teropode, differiva dai neotopodi in numerosi modi. La cavità cerebrale era relativamente più grande e le mani erano più sottili. La coda tendeva ad essere molto rigida.
Alcuni elementi ritenuti omologhi alle piume degli uccelli, chiamati protofeathers da alcuni ricercatori, sono stati trovati in celurosauri sia primitivi che avanzati, ed è stato suggerito che tutti i celurosauri primitivi potrebbero averli avuti. Diversi celurosauri giurassici e cretacei, sia piccoli che grandi, non erano ancestrali a lame più avanzate, ma un clade chiamato maniraptor divenne sempre più specializzato.
I maniraptor sono caratterizzati dallo sviluppo di un palato secondario e da numerosi cambiamenti nella struttura del caso cerebrale. Avevano anche mani molto sottili e meno vertebre di coda. Gli uccelli sono un sottoprodotto dell’evoluzione del maniraptor.
Esistono diversi importanti gruppi di maniraptor; tra questi, gli ornitomimosauri, come lo Struthiomimus, non sono antenati degli uccelli, ma mostrano una convergenza con struzzi moderni e altri uccelli incapaci di volare. I primi esempi avevano denti piccoli, ma i denti andarono persi in forme successive. Le dita delle mani formavano una struttura a gancio che poteva essere stata usata per afferrare i rami mentre gli animali cercavano cibo.
I tirannosauridi sono ben noti sotto forma di Tyrannosaurus rex, popolarmente noto come T. rex, così come altri grandi carnivori. Va sottolineato che i tirannosauridi si sono evoluti indipendentemente dall’Allosauro dalla faccia stretta e dai suoi parenti.
I tiarannosauridi sono caratterizzati da teste massicce, piuttosto che strette; denti stridenti (incisivi) nella parte anteriore delle mascelle; denti laterali ispessiti anziché compressi sui lati delle mascelle; e arti anteriori molto minimali con solo due artigli.
Questi animali correvano veloci per le loro dimensioni e solo con la mascella potevano uccidere la loro preda e renderla in porzioni abbastanza piccole da essere ingoiate. I primi tirannnosauridi del Cretaceo erano lunghi solo circa 3 m (10 piedi), ma quelli successivi come il Tyrannosaurus rex erano lunghi 40- 12 piedi (12,2-15,2 m) con un peso fino a sette tonnellate o più.
Altri maniraptor comprendono gruppi come gli oviraptorosauri, di cui Oviraptor è un genere ben noto. Molte specie di oviraptosauri sono caratterizzate da stemmi ornati e si pensa che abbiano covato i loro nidi alla maniera degli uccelli moderni.

I dromaeosauridi comprendono i noti generi Deinonychus, Velociraptor e il più grande Utahraptor. Questi animali avevano arti anteriori lunghi e afferranti e un artiglio grande, retrattile e curvo sulla seconda cifra del piede. I dromaeosauridi, come gli uccelli, avevano un pube rivolto all’indietro e alcuni ricercatori ritengono che sia vicino allo stelo dell’evoluzione degli uccelli.
Trovato tra i Sauropodomorpha sono i più grandi animali terrestri che siano mai vissuti; alcuni hanno raggiunto una lunghezza di circa 30 m. Tutti i sauropodi avevano il collo lungo e la testa piccola. Alcuni dei sauropodomorfi primitivi, noti come prosauropodi (come il Plateosaurus), trascorsero un po ‘di tempo a carponi, sebbene potessero ancora afferrare oggetti con la mano. Alla fine del Triassico, i prosauropodi erano comuni erbivori di grandi dimensioni. I lunghi colli davano ai sauropodomorfi l’accesso alle foglie degli alberi e le loro grandi dimensioni corporee consentivano loro di avere un sistema digestivo più ampio per elaborare la vegetazione e fungere da difesa contro i piccoli predatori.
I sauropodomorfi chiamati Neosauropoda divennero un gruppo diversificato nel Giurassico e nel Cretaceo. Neosauropods
erano caratterizzati da gambe colonnari, fianchi larghi e denti specializzati. Le due principali pale di neosauropodi sono la Diplodocoidea e la macronaria. Diplodocoideans aveva denti sottili a forma di matita che presumibilmente venivano usati per raccogliere foglie e aghi dai rami degli alberi. Il cranio era lungo e inclinato e le narici erano posizionate dietro gli occhi. Le code dei diplodocoidi erano molto lunghe e frustate. Esempi familiari sono Apatosaurus (a lungo chiamato Brontosaurus) e Diplodocus. Le forme a collo più corto includono l’Amargasaurus del primo Cretaceo dell’Argentina.
Nella macronaria (“grandi narici”) i clade sono sauropodi con enormi narici che sono più grandi delle orbite. Parecchi membri di questo gruppo avevano anche denti a forma di spatola. I brachiosauri, compreso il noto genere Brachiosaurus, formano un clade che aveva arti anteriori molto più lunghi degli arti posteriori e un collo molto lungo, una condizione che ricorda le giraffe moderne.
La combinazione di lunghe zampe anteriori e un lungo collo ha permesso a questi animali di nutrirsi in cima agli alberi. Si stima che Sauroposeidon del Cretaceo dell’Oklahoma fosse lungo 80 piedi (24,4), circa 40 piedi (12,2) costituiti dal collo. I titanosauri erano giganteschi sauropodomorfi macronaristi che formano un clade a parte i brachiosauri. È stato scoperto che le forme avanzate in questo gruppo hanno sviluppato denti simili a quelli dei diplodocidi.
Gli Ornithischia sono la terza e ultima delle tre principali pale di dinosauro. Questi animali erano principalmente erbivori a quattro zampe corazzati, con becco, ma alcuni degli esempi primitivi erano almeno parzialmente bipedi. Le tre principali pale attualmente riconosciute all’interno di Ornithischia sono la Thyreophora (dinosauri corazzati), Ornithopoda (dinosauri con becco, becco e cresta) e Marginocephalia (dinosauri con elmo e cornuto).
Nella diversificazione dei tireoforani, si scopre che la modifica degli scudi corazzati in altre strutture difensive è una tendenza centrale. Lo Scutellosauro relativamente privo di corazza del primo Giurassico, ritenuto un membro basale del gruppo, aveva una fila di placche dermiche ossee dal collo alla coda.
Si pensa che questa caratteristica avrebbe potuto proteggerla dai piccoli predatori del tempo, ma probabilmente non dai grandi predatori che apparvero in seguito nel Giurassico. In effetti, le forme successive hanno modificato questa armatura dermica in piastre, punte e armatura di coda.
Gli Stegosauri del tardo Giurassico erano veri e propri dinosauri corazzati, in quanto avevano compresse compresse, spine coniche o armature di forma intermedia lungo il centro della schiena e code modificate come mazze appuntite. Alcuni stegosauri avevano punte di spalla e altri avevano masse di pomelli sulla gola. Gli anchilosauri avevano un’armatura dermica simile ad un armadillo fusa alla testa e proteggendo la maggior parte del corpo. Un gruppo di anchilosauri aveva la coda del club.
Gli ornitopodi possono essere distinti dagli altri ornitischia in quanto la fila di denti nella porzione anteriore della mascella superiore (premaxilla) è più depressa rispetto a quella nella restante mascella superiore. Le tendenze evolutive in questo gruppo includevano un aumento delle dimensioni e cambiamenti nelle articolazioni della mascella e dei denti che hanno portato a un modo macinante di masticazione.
Iguanodon, un antico ornitopodo scoperto per la prima volta dai primi Cretacei d’Inghilterra, un tempo era considerato una lucertola gigante, perché aveva denti dentellati simili a foglie come la moderna lucertola iguana. I dinosauri a becco d’anatra del tardo Cretaceo, con musi espansi alla fine, sono stati eccezionalmente ben studiati, e alcuni sono conosciuti letteralmente dalla culla (nido e uova) alla tomba.
Ogni specie di lambeosauri aveva una forma unica di cresta e tubi che producevano suoni all’interno del cranio. Queste caratteristiche hanno fornito segnali sia visivi che vocali che indicano un comportamento sociale complesso in questi animali.
L’ultimo gruppo di Ornithischia sono i marginocefali (dinosauri con elmo e cornuto), rappresentati sia da forme bipede che a quattro zampe. Il personaggio principale di questo clade era una cresta o una mensola di osso che si sovrapponeva alla parte posteriore del cranio. Questo gruppo è diviso in due lame, i pachycephalosaurs e i ceratopsians. I pachicefalosauri erano forme bipede con un grosso elmo di osso sopra la cassa del cervello; nelle forme più derivate, l’elmo formava una cupola ispessita.
Molti scienziati ritengono che la cupola sia stata utilizzata per contrastare gli scontri, come accade oggi tra le pecore Bighorn. Altri pensano che, poiché la cupola era dotata di un gran numero di vasi sanguigni, avrebbe potuto essere importante nella regolazione della temperatura.
I ceratopsiani avevano grandi becchi alla fine del muso. Un primitivo ceratopsian, Psittacosaurus, del primo Cretaceo aveva questo becco profondo ma era bipede e mancava il fronzolo di forme più avanzate. Le forme avanzate, chiamate neoceratopsiani, avevano uno scaffale modificato come un volant nella parte posteriore del cranio in cui erano attaccati i muscoli della mascella.
I neoceratopsiani avevano file di denti raggruppati in modo tale da fornire una superficie di taglio continua, ma non è noto su quale tipo di piante si nutrissero questi animali. I giganteschi ceratopsiani nordamericani avevano elaborato notevolmente le corna sul cranio.
Risorse – Libri
Benton, Michael J. Vertebrate Paleontology. 2a edizione. Oxford: Blackwell Science, 2000.
Benton, Michael J. e DAT Harper. Paleontologia di base. Londra: Addison Wesley Longman, 1997.
Carroll, Robert L. Modelli e processi nell’evoluzione dei vertebrati. Cambridge: Cambridge University Press, 1997.
Colbert, Edwin H., Michael Morales ed Eli C. Minkoff. Evoluzione dei vertebrati di Colbert. 5a edizione. New York: John Wiley and Sons, Inc., 2001.
Cowan, Richard. Storia della vita. Edizione 3d. Malden, MA: Blackwell Science, 2000.
Farlow, James O. e MK Brett-Surman, a cura di. Il dinosauro completo. Bloomington e Indianapolis: Indiana University Press, 1997.
Hallam, Arthur e PP Wignall. Estinzioni di massa e loro conseguenze. New York: Oxford University Press, 1997.
Holman, J. Alan. Vita vertebrata del passato. Dubuque, IA: William C. Brown Publishers, 1994.
-. Serpenti fossili del Nord America: origine, evoluzione, distribuzione, paleoecologia. Bloomington e Indianapolis: Indiana University Press, 2000.
Lucas, Spencer G. Dinosaurs, il libro di testo. Edizione 3d. Dubuque, IA: William C. Brown, editori, 2000.
Paul, Gregory S., ed. Il libro scientifico americano dei dinosauri. New York: St. Martin’s Press, 2000.
Pough, F. Harvey, Robin M. Andrews, John E. Cadle, Martha L. Crump, Alan H. Savitzky e Kentwood D. Wells. Erpetologia. 2a edizione. Upper Saddle River, NJ: Prentice Hall, 2001.
Sumida, Stuart S. e Karen LM Martin. Amniote Origins, Completando la transizione verso la terra. San Diego: Academic Press, 1997.
Weishampel, David B., Peter Dodson e Halszka Osmólska, a cura di. La Dinosauria. Berkeley: University of California Press, 1990.
Ulteriori informazioni: Tiago R. Simões et al., Dinamica megaevolutiva e tempistica dell’innovazione evolutiva nei rettili, Nature Communications (2020). DOI: 10.1038 / s41467-020-17190-9



{kind=link}