










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



A research group centered around Kobe University Graduate School of Medicine’s Professor Yamada Hideto and Associate Professor Tanimura Kenji (Department of Obstetrics and Gynecology), and Professor Arase Hisashi et al. of Osaka University’s Research Institute for Microbial Diseases (RIMD) have revealed for the first time in the world the high frequency of a novel autoantibody in women suffering from recurrent pregnancy loss.
Joint research by Kobe University and Osaka University in 2015 previously discovered that this autoantibody also causes diseases such as thrombosis.
It is expected that these results will contribute towards the illumination of the underlying mechanisms behind recurrent pregnancy loss, thrombosis and pregnancy complications such as hypertensive disorders; leading to the development of new treatment methods.
The findings were published online on June 25 in the American College of Rheumatology’s journal ‘Arthritis and Rheumatology‘.
- Those with recurrent pregnancy loss (RPL) experience repeated miscarriages and stillbirths, making them unable to give birth to a healthy baby.
- In Japan, approximately 1,400,000 women are believed to suffer from RPL. In more than half of cases the cause cannot be determined, making it difficult to prescribe a treatment method. This is a big problem, especially in light of Japan’s low birthrate and aging population.
- This study was based at Kobe University, which is putting a lot of effort into RPL research, and covered 227 patients in five university hospitals across the country. Approximately a quarter of these patients tested positive for the novel autoantibody (neo-self antibody). Furthermore, almost one fifth tested positive for the neo-self antibody among patients for whom common tests had been unable to determine the cause.
- It is expected that these results can illuminate the underlying mechanisms behind RPL, thrombosis and hypertensive disorders of pregnancy, leading to the development of new treatment methods. Furthermore, it is hoped that these research results can be the key to solving the low birthrate and aging population issues.
Recurrent pregnancy loss (RPL) is a disorder in which the sufferer is able to get pregnant yet experiences repeated miscarriages and stillbirths, making them unable to give birth to a healthy baby. There is believed to be an estimated 1,400,000 women with RPL in Japan.
Given Japan’s low birthrate and aging population, this is an important issue to overcome. However, at present the cause remains unknown in over half of patients, making it difficult to know how to cure RPL in many cases.
However, joint research conducted by RIMD’s Professor Arase Hisashi and Kobe University’s Associate Professor Tanimura Kenji discovered a completely new autoantibody that causes antiphospholipid syndrome.
This syndrome can lead to the occurrence of thrombosis (in which blood clots can block up blood vessels in vital organs, endangering life such as by causing a stroke), miscarriages, and hypertensive disorders of pregnancy, which can endanger the lives of pregnant women. These findings were published as a thesis in the American Society of Hematology’s journal ‘Blood’ in 2015.
RPL involving repeated miscarriages is also a clinical manifestation of antiphospholipid syndrome. As previously mentioned, the cause of RPL is unknown in over half of cases, however the association between this neo-self antibody and RPL had not been investigated.
This research group hypothesized that the neo-self antibody could be connected to RPL in cases with unexplained causes.
With this in mind, blood samples were collected from RPL patients across five university hospitals (Kobe University, University of Toyama, Okayama University, The University of Tokyo, and Hyogo Medical University), and the samples were tested for neo-self antibodies at RIMD.
For the first time in the world, the research group conducted this clinical study to illuminate the association between RPL and the neo-self antibody.
This study was led by Kobe University, which is putting a lot of effort into RPL research, and involved testing for neo-self antibodies with the consent of outpatient couples affected by RPL at five university hospitals nationwide.
At the same time, a detailed examination into the cause of RPL in each case was carried out. This included testing for thyroid disorders, a chromosome analysis of each couple, and blood tests to investigate conditions that make blood clots form easily, such as the presence of antiphospholipid antibodies.
Also, tests were carried out on the couples for genetic complexes that are associated with susceptibility to various diseases, such as Human Leukocyte Antigen (HLA) Class II. (For example, it is known that those with HLA-DR4 are prone to RPL).
The neo-self antibody testing method used in the study is a patented technology developed by this research group.
This method involves producing cells where the complexes produced byβ2-glycoprotein I (a protein that is believed to be targeted by the antibody that causes antiphospholipid syndrome) and HLA Class II (the HLA type that increases susceptibility to antiphospholipid syndrome) are present on the cell surface.
A reaction is then conducted between the complexes and the patient’s blood to detect antibodies (neo-self antibodies), which bind to the complexes on the cell surface.
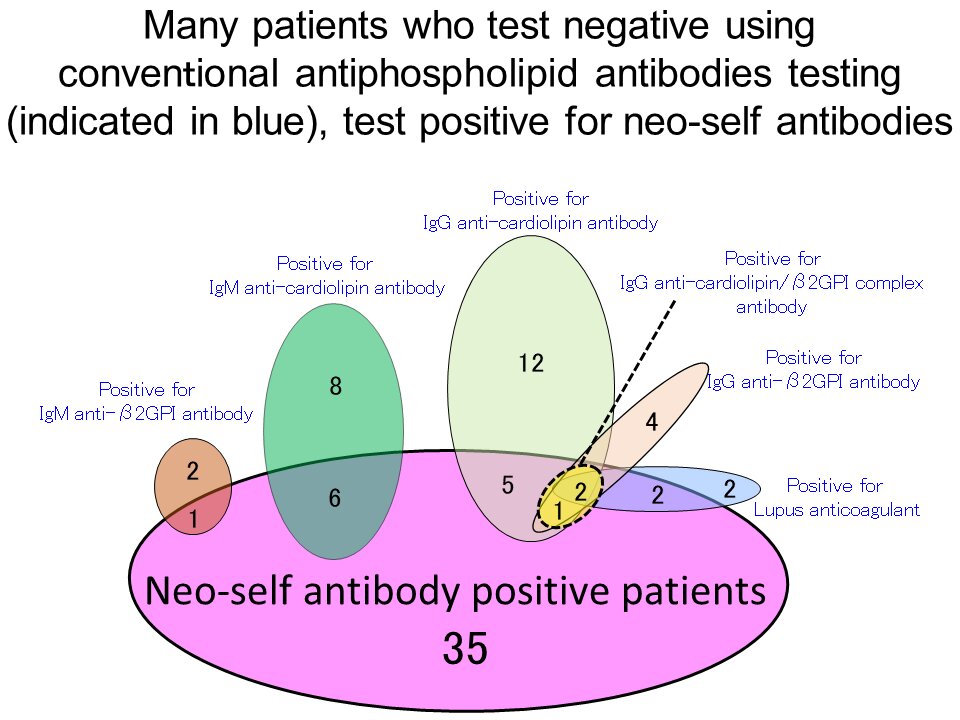
First of all, 208 women who did not have RPL and had previously given birth to healthy babies were tested for neo-self antibodies to establish normal levels.
Subsequently, it was found that 52 (23%) out of 227 patients with RPL tested positive for the neo-self antibody.
The results of the tests carried out to determine the causes of RPL were evaluated. The neo-self antibody was the factor with the highest frequency compared to all other factors that were tested, including uterine issues (such as malformation and myomas), thyroid dysfunction, and chromosomal abnormalities in each partner.
This indicts the possibility that this neo-self antibody is a major cause of RPL.
Even after testing for common RPL causes, the cause could not be determined in around half of the patients (121 women). However, 24 (20%) of the 121 women with unexplained RPL were found to test positive for only the neo-self antibody (Figure 1).
In particular, many who tested negative for antiphospholipid antibody criteria tested positive for the neo-self antibody (Figure 2).
When neo-self antibody positive women with RPL were compared with those RPL patients who tested negative, it was found that the frequency of those who had the HLA-DR4 gene, which predisposes the patient to RPL, was higher in neo-self antibody positive women than in negative women.
Currently, it is unclear as to why those with HLA-DR4 are prone to RPL. It is hoped that this study’s discoveries can serve as a key to understanding the reason.
By investigating the neo-self antibody, the researchers were able to illuminate the onset mechanism of RPL, in particular the onset mechanism of the disorder in some unexplained cases. This could lead to a solution for Japan’s low birthrate and aging population issues.
This study has shown that the novel autoantibody (neo-self antibody) that was discovered by Associate Professor Tanimura and Professor Arase et al. is a major cause of RPL.
Next, the researchers aim to generate a drug which inhibits neo-self antibodies or suppresses their production. In addition, it is hoped that research into neo-self antibodies will illuminate the onset mechanism for obstetrics patients with conditions, such as unexplained hypertensive disorders of pregnancy and fetal growth restriction, where the cause is unknown.
This could lead to the development of treatment methods for these patients. Furthermore, it is possible that there is a neo-self antibody that triggers many common autoimmune disorders, such as rheumatism. Such a discovery could be revolutionary for both rheumatology and immunology.
Recurrent pregnancy loss (RPL) is the loss of two or more pregnancies before 24 weeks of gestation, according to the European Society of Human Reproduction and Embryology [1]. The Practice Committee of the American Society for Reproductive Medicine defines RPL as two or more failed clinical pregnancies [2].
RPL is a challenge for the clinical and scientific community. In fact, in only around 50% of RPL cases can defined causes/risk factors can be found, including advanced maternal age, genetic abnormalities, selected maternal autoantibodies, endocrine dysfunctions, and uterine abnormalities [3]. The remaining RPL cases currently are unexplained (uRPL) [3,4].
It is believed that a significant, although not exactly quantified, proportion of RPL is associated with immune etiologies [5] and that in these cases the pregnancy losses can occur through persistent disturbances in several immune pathways [6]. In this context, a relevant role could be played by the endometrium.
The endometrium has a crucial role in reproduction. It is the maternal tissue that comes into direct contact with the embryo and allows for its proper implantation, survival, and development, processes in which it actively participates.
Extensive investigation has been carried out in the last decades to clarify the biomolecular mechanisms which make the endometrium receptive to the embryo, as well as the specific cell types and the cellular pathways involved in endometrial receptivity. However, our knowledge on these mechanisms is still largely incomplete.
The currently available information indicates that the endometrium is a unique tissue in which a series of events – collectively called decidualization – occur in order to ensure, in a dynamic fashion, the correct environment for the developing conceptus.
Emerging evidence indicates that a pivotal role in endometrial remodeling and maternal tolerance towards the embryo is played by several cells of the innate and adaptive immune system, which have or acquire specific characteristics when they are in or reach the endometrium, together with a growing number of immunoregulatory molecules.
The experimental and clinical evidence also suggests that derangements occurring in the endometrial immune environment can be involved in several important reproductive dysfunctions, such as recurrent implantation failure (RIF) and recurrent pregnancy loss (RPL) of otherwise unexplained etiology.
This review summarizes the major contributions of the immune system in the physiology of the endometrial function and the current knowledge on the major known alterations of the endometrial immune system with specific application to RPL in women.
Physiological Endometrial Function and the Immune System
Endometrial Remodeling and Decidualization
The endometrium is the maternal tissue that comes into direct contact with the embryo, which is immunologically different from the mother, being usually considered as a semi allograft, or even as a complete allograft in the case of oocyte donation [7].
The major function of the endometrium is to create and ensure an optimal endocrine/paracrine, immune, and molecular environment that allows for proper apposition, attachment, implantation, invasion and development, and full maturation of the embryo.
To accomplish this mission, the endometrium needs to develop a series of unique and striking adaptive changes collectively termed decidualization, resulting in profound morphological and functional reprogramming of the endometrial stromal cells that differentiate into highly specialized cells with secretory capabilities.
Among the eutherian – placental mammals – these changes are particularly significant in species with an invasive type of placenta, specifically in humans who have the most invasive type of placentation.
In fact, the extent of decidualization is proportional to the degree of invasiveness of the embryo [8]. While endometrial transformation in most animal species occurs in response to the presence of the embryo, in a few species – including humans – it takes place well in advance of the presence of the embryo and, therefore, is – at least initially – under exclusive maternal hormonal control.
If conception does not occur, in response to falling circulating levels of progesterone the decidualized endometrium in these species is shed and menstruation ensues. This initiates the beginning of a new cycle.
The majority of recent information on the role of the endometrium and on the functional changes occurring in the normal endometrium stems from the histologic investigations performed on human endometrial biopsies, as well as from studies carried out in vivo in animal species (particularly baboons and gene knockout mice) and in vitro in primary cell cultures of human endometrial stromal cells (HESCs).
Despite the considerable differences among the species, the overall emerging picture is that decidualization of the endometrium is a process involving profound cell reprogramming, tissue remodeling, changes in gene expression and post-translation regulation, and alterations in cell signaling pathways.
Considerable changes and modulation of the activity of the immune cells at a local level also occurs. These events involve a large number of molecular mediators and effectors. A summary of the major endometrial changes characteristic of decidualization in women is described in Table 1.
Table 1
Simplified summary of the major changes characterizing the decidualization of the endometrium.
| Morphological (Tissutal and Cell) Changes |
| Secretory transformation of endometrial glands, compaction of surface epithelial cells, stromal edema, stromal cell proliferation, differentiation of fibroblast-like stromal cells into epithelioid-like cells, massive leukocyte infiltration (mainly uterine natural killer cells and mast cells), spiral arteries remodeling |
| Extracellular Matrix Changes |
| Increased production by decidualized stromal cells of collagen IV, fibronectin, laminin, decorin, heparan-sulphate proteoglycans, IGFBP-1, PRL, LEFTY-2, αvβ3 integrin, osteopontin |
| Changes in the Expression of Genes Involved in the Following Cellular Functions |
| Cell cycle regulation, cytoskeleton remodeling, oxidative stress response, ion and water transport, response to steroid hormone, deposition of extracellular matrix (ECM), modulation of transcription, epigenetic patterning, post-translation modification, growth factor, angiogenesis, cytokine and chemokine signaling, immune modulation |
| Cell Signaling and Pathways Involved in the Decidualization |
| cAMP/PKA pathway, progesterone signaling pathway, Nodal pathway, Notch signaling pathway, ERK1/2 pathway, WNT/β-catenin pathway, cSRC pathway, JAK-STAT pathway, lipid signaling (endocannabinoid system), TGFβ signaling pathway, BMP2-WNT4 signaling cascade, phosphatidylinositol 3-kinase/AKT pathway, Ras/Raf1/MAPK pathway, EPAC1 and EPAC2 signaling |
LEFTY-2: left-right determination factor 2; IGFBP-1: insulin like growth factor binding protein-1; PRL: prolactin; cAMP: cyclic adenosine monophosphate; PKA: protein kinase A; ERK1/2: extracellular signal-regulated kinase 1/2; JAK: Janus kinase; STAT: signal transducer and activator of transcription; TGFβ: transforming growth factor-β; BMP-2: bone morphogenetic protein-2; AKT: protein kinase B; MAPK: mitogen-activated protein kinase; EPAC1 and 2: exchange protein directly activated by cAMP 1 and 2.
The detailed description of all the changes and the mediators involved in decidualization is beyond the scope of the present article and has been discussed in several excellent reviews recently [9,10,11,12,13]. Here, the major focus is to evaluate the significance of the endometrial changes, particularly decidualization, in relation to the role played by the immune system.
Decidualization can be evolutionarily considered to have two, not mutually exclusive functions. On the one hand, it can be considered as the biological solution of a maternal–fetal conflict in which the invasiveness of the embryo, aimed to maximally ensure the expansion of fetal genes (partially different from the maternal ones) is contrasted by the need of maternal genes to ensure their own expansion in the present and future pregnancies [14].
On the other hand, it can be viewed as the adaptive response to the maternal need of controlling its relevant investment in a pregnancy bearing a single, high quality fetus for a prolonged period of time [15].
This concept is supported by robust evidence indicating that the decidualized endometrium acts as a biosensor of the quality of the embryo [16,17], that implantation is characterized by a cooperation between the embryo and the decidualized endometrium [18], and that the loss of endometrial plasticity can be implicated in unexplained RPL [19].
Many key events occurring in the endometrium during decidualization implantation and in the decidua throughout pregnancy are realized with the relevant contribution of the immune cells and are mediated by many immunoregulatory molecules.
Emerging evidence suggests that derangements in the normal immune function can occur in these tissues in subsets of women with RPL.
Immune Cells and Their Functions in the Normal Endometrium and Decidua
A great body of experimental work has been carried out to investigate the specific populations of immune cells in the endometrium throughout the menstrual cycle, during implantation, and early pregnancy; their specific roles in early pregnancy together with the regulatory molecules that are expressed are being characterized.
This information is summarized in Table 2 and Table 3. Likewise, a growing number of mechanisms are proposed to explain the development of maternal tolerance towards the immunologically different conceptus.
These proposed mechanisms—schematically summarized in Table 4 and illustrated in Figure 1a,b—could be the basis to explain, understand, and, possibly, treat the immune-mediated RPL, that likely represent a substantial proportion of all unexplained RPL. However, it is actually extremely difficult to disentangle the most important mechanisms from the accessory ones.
The emerging overall picture strongly suggests that in normal pregnancy, the maternal immune system undergoes considerable modulation in at least many (if not all) key components in order to develop tolerance towards the foreign paternal antigens of the fetus and an immunologically favorable environment for the fetus while at the same time maintaining full responsive capacity against other foreign antigens [20,21,22,23].
The pregnancy-related changes in the immune cells and system occurring in the endometrium and decidua are only partially determined. Nevertheless, with regard to the success of implantation, the most important changes known so far in the endometrial immune cells involve macrophages, uterine natural killer (NK) cells, dendritic cells (DCs), T cells, especially cytolytic T cells (CTLs), whose specific roles have been, at least in part, determined.

Schematic representation of the changes occurring in the human endometrium and in local immune cell trafficking in the normal state and in recurrent pregnancy loss (RPL). (a) Endometrium in the secretory phase of the menstrual cycle in the absence of the embryo; (b) endometrium in the presence of a normally implanting embryo; (c) endometrial immune derangements in RPL. M: macrophages; uNK: uterine natural killer cell; iDC: immature uterine dendritic cell; mDC: mature uterine dendritic cell; N: neutrophil granulocyte; Treg: regulatory T cell; MC: mastocyte; Teff: effector T cells; ESC: endometrial stromal cell; BV: blood vessel.
Table 2
Overview of the relevant cells of the innate immune system in the endometrium and decidua with their major reproductive functions.
| Cell Type | Phenotype | Density (%) of Leukocyte Population in the Endometrium | Changes during the Menstrual Cycle and in Early Pregnancy | Preferential Location in the Endometrium | Relevant Molecules Secreted | Suggested Functions | References |
|---|---|---|---|---|---|---|---|
| Uterine Natural killer Cells (uNK) | CD3−CD56brightCD16− (predominant phenotype) | 30%–40% of stromal cells 70% of endometrial leukocytes in the late luteal phase (LP) and in early pregnancy | Progressively increase from the follicular phase (FP) to the LP. Maximal density in late LP and in gestational decidua | Surround the arteries and the glands | IFN-γ, VEGF, PlGF, TGF-β, TNF-α, IL-10, GM-CSF, IL-1β, LIF, CSF-1, AP-2 | Tissue (spiral arteries) remodeling, enhancement of angiogenesis, control of trophoblast invasion | [10,20,24,25,26,27,28,29] |
| Macrophages (Mφ) | CD68+ | 20%–25% of total leukocytes in the decidua | Progressively increase from the FP to the LP. Maximal density before menstruation and in pregnancy. Acquire tolerogenic phenotype | Scattered throughout the endometrium; preferentially found around the glands and at implantation site | TGF-β, IL-10, IDO, PGE2 | Involved in corpus luteum maintenance, blastocyst implantation, spiral arteries remodeling, control of trophoblast invasion, protection of the fetus against intrauterine infection | [20,29,30,31,32,33,34,35] |
| Mast Cells (MCs) | MCT; MCTC; MCC (endometrial MCs) | 3%–5% of total endometrial cells; | Unchanged throughout menstrual cycle; changes in phenotype during the menstrual cycle; activated in the early and midluteal phase | More prominent in the basal endometrial compartment | VEGF | Initiation of menstruation Enhance tissue and spiral artery remodeling, support implantation and angiogenesis | [20,36,37] |
| Uterine Dendritic Cells (DCs) | CD1a+ (immature, tolerogenic DCs); CD83+ (mature DCs) | Density of immature DCs in the endometrium is higher than that of mature DCs; DC 1%–2% of the immune cells in the decidua | Immature DCs increase from FP to LP; peak in the menstrual phase (controversial finding); No changes in mature DCs with the menstrual cycle | Both mature and immature DCs are found mainly in the basal layer of the endometrium in the LP; Scattered through the gestational decidua In mice grouped in cluster-like structures | TGF-β, IL-10, IDO | Involved in the maternal acceptance of the embryo, trophoblast invasion and differentiation; Uterine remodeling, angiogenesis; Determine the differentiation of T cell progenitors into Tregs and expansion and activation of Tregs | [29,33,38,39,40,41,42,43] |
| Neutrophil (N) Granulocytes | CD11b+; CD16b+; CD66c+ | 1% to 6%–15% of endometrial cells | Considerably increase in the late (premenstrual) LP | Endometrial Stroma | CCL2, CXCL8, TNF-α, IL-6, VEGF | Menstruation, tissue breakdown and repair, Proangiogenic and tolerogenic in the pregnant decidua |
Table 3
Overview of the cells of the adaptive immune system in the endometrium and decidua with their major reproductive functions.
| Cell Type | Phenotype | Density (%) of Leukocyte (CD45+) Population in the Endometrium | Changes during the Menstrual Cycle and in Early Pregnancy | Preferential Location in the Endometrium | Relevant Molecules Secreted | Suggested Functions | References |
|---|---|---|---|---|---|---|---|
| BLymphocytes | CD45+ CD19+ | 0.2%–4.5% | Very slight increase in late LP | Clusters among stromal cells in the perimenstrual period | IL-10 | Still undetermined; Potentially implicated in early pregnancy | [20,47,48] |
| T Lymphocytes | CD45+ CD3+ | 1%–2% to 28% | Reportedly decreased or unchanged from follicular phase (FP) to luteal phase (LP) | Lymphoid aggregates; scattered throughout epithelium and stroma | Variable according to the specific cell subset | Protective or harmful for the embryo according to the specific cell subset | [24,29,48,49] |
| T Helper Lymphocytes (Th1, Th2, Th17, Tregs) | CD45+ CD3+ CD4+ | 3.8%–21.4% | No clear variations reported | Present in uterine mucosa as unique aggregates surrounding a B cell core | TNF-α and IFN-γ (by Th1); IL-4 (by Th2); IL-8 (by Th17) | Th1 produce inflammatory cytokines; Th2 produce anti-inflammatory cytokines; Th17 has pro-inflammatory effects (for Tregs see the dedicated section in the table) | [33,48,50] |
| T Cytotoxic Lymphocytes | CD45+ CD3+ CD8+ | 4.4%–34.5% to 66% | Significantly decrease from FP to LP | Lymphoid aggregates | Release cytotoxic substances (granzymes, perforin) | Potentially harmful to the embryo. Blocked in successful pregnancy | [47,48,49,51] |
| Tregs | CD4+ CD25+ FOXP3+ | Not clearly defined in humans | Expansion in preimplantation endometrium; Increase in the decidua at implantation site and in early pregnancy until midgestation | Insufficient data in human preimplantation endometrium | Galectin-1, TGF-β, IL-10, HO-1 | Essential in the control of an excessive maternal inflammatory response at the implantation site; Involved in maternal immune tolerance to fetal allograft particularly in early pregnancy; Block maternal effector T cells Involved in the regulation of maternal vascular remodeling | [20,33,49,50,52,53,54,55,56,57] |
CD: cluster of differentiation; FP: follicular phase; LP: luteal phase; IFN-γ: interferon gamma; VEGF: vascular endothelial growth factor; PlGF: placental growth factor; TNF-α: tumor necrosis factor alpha; IL-10: interleukin-10; GM-CSF: granulocyte macrophage colony stimulating factor; IL-1β: interleukin-1β; LIF: leukemia inhibitory factor; CSF-1: colony-stimulating factor 1; AP-2: endocytic adaptor protein 2; IDO: indoleamine 2,3-dioxygenase; PGE2: prostaglandin E2; CCL2: chemokine C-C motif ligand 2; CXCL8: C-X-C motif ligand 8; IL-6: interleukin-6; IL-4: interleukin-4; IL-8: interleukin-8; HO-1: heme oxygenase-1.
Table 4
Proposed major mechanisms underlying maternal immune tolerance towards the embryo in normal pregnancy in the endometrium and decidua.
| Mechanism | Effect | References |
|---|---|---|
| Increased secretion of LIF and IL-1β by uterine macrophages | Enhancement of embryo attachment to endometrial epithelium | [58] |
| TGFβ production by maternal decidual macrophages | Suppression of EVT rejection mediated by NK cells | [59] |
| Production by macrophages of TGFβ, IL-10, IDO, and PGE2 | Immunoinhibitory and pro-tolerance actions | [33] |
| Specific characteristics of uNK cells in the endometrium and of dNK in pregnancy | Acquisition of a regulatory role rather than classic cell killing ability | [21,27,60] |
| dNK cells dampen Th17 cells through the production of IFN | Suppress Th17-induced inflammatory response | [61] |
| Galectin-1 production by dNK cells | Induces apoptosis of activated CD8+ T cells | [62,63,64] |
| Endometrial recruitment of innate immune cells (Mφ, DCs, and granulocytes) triggered by seminal fluid before implantation | Activation and expansion of Tregs which, in turn, creates a uterine microenvironment favorable for embryo implantation and enhances maternal tolerance towards paternal MHC antigens | [21,54,65,66,67] |
| Induction and expansion of tolerogenic DCs phenotype | Involved in the control and activation of Tregs | [33,54,68,69] |
| Reduced production of IL-12 by DCs | Priming of decidual CD4+ cells into a Th2 phenotype | [70,71] |
| EVT cells express HLA-C, HLA-E, and HLA-G but not HLA-A and HLA-B Lack of expression of MHC molecules by syncytiotrophoblasts | Inhibition of cytolytic activity of dNK cells against the trophoblasts Induction of NK senscence-Enhanced apoptosis of activated CD8+ cells | [70,72,73,74,75] |
| Trophoblast production of exosomes | Downregulation of maternal immunity towards the trophoblast | [76] |
| Development of a specific cytokine and chemokine network in the endometrium and decidua | Achievement of a correct immune cell recruitment and dialogue favoring embryo implantation and proper pregnancy evolution | [77] |
| Presence of asymmetric maternal Abs against paternal antigens | Bind trophoblast but are unable to trigger destructive immune response | [78] |
| Increased production of PIBF | Upregulation of Th2 cytokines production. Downregulation of dNK activity. Increase the production of Glycodelin A which induces apoptosis in T cells. These effects can contribute to trophoblast immune protection | [66,79] |
| Increased progesterone production in LP and in pregnancy | Expansion of Tregs and enhancement of their immunosuppressive actions | [33,78] |
| hCG production by syncytiotrophoblast | Recruitment of Tregs at the maternal–fetal interface | [80] |
EVT: extravillous trophoblast; HLA: human leukocyte antigen; MHC: major histocompatibility complex; Mφ: macrophages; DC: dendritic cells; PIBF: progesterone induced blocking factor; LP: luteal phase; hCG: human chorionic gonadotropin; Abs: asymmetric antibodies.
reference : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6862690/
More information: Kenji Tanimura et al, The β2‐glycoprotein I/HLA‐DR complex is the major autoantibody target in obstetric antiphospholipid syndrome, Arthritis & Rheumatology (2020). DOI: 10.1002/art.41410



{kind=link}