










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



With an alarm code, we can enter a building without bells going off. It turns out that the SARS coronavirus 2 (SARS-CoV-2) has the same advantage entering cells.
It possesses the code to waltz right in.
On July 24 in Nature Communications, researchers at The University of Texas Health Science Center at San Antonio (UT Health San Antonio) reported how the coronavirus achieves this.
The scientists resolved the structure of an enzyme called nsp16, which the virus produces and then uses to modify its messenger RNA cap, said Yogesh Gupta, Ph.D., the study lead author from the Joe R. and Teresa Lozano Long School of Medicine at UT Health San Antonio.
“It’s a camouflage,” Dr. Gupta said. “Because of the modifications, which fool the cell, the resulting viral messenger RNA is now considered as part of the cell’s own code and not foreign.”
Deciphering the 3-D structure of nsp16 paves the way for rational design of antiviral drugs for COVID-19 and other emerging coronavirus infections, Dr. Gupta said.
The drugs, new small molecules, would inhibit nsp16 from making the modifications.
The immune system would then pounce on the invading virus, recognizing it as foreign.
“Yogesh’s work discovered the 3-D structure of a key enzyme of the COVID-19 virus required for its replication and found a pocket in it that can be targeted to inhibit that enzyme.
This is a fundamental advance in our understanding of the virus,” said study coauthor Robert Hromas, MD, professor and dean of the Long School of Medicine.
Dr. Gupta is an assistant professor in the Department of Biochemistry and Structural Biology at UT Health San Antonio and is a member of the university’s Greehey Children’s Cancer Research Institute.
In lay terms, messenger RNA can be described as a deliverer of genetic code to worksites that produce proteins.
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has caused the coronavirus disease (COVID-19) global pandemic 1, which has currently led to more than 10 million confirmed cases and more than 500 thousands deaths in over 200 countries according to the World Health Organization (www.who.int).
Coronaviruses have long been a threat, but recent developments show that they should be classified as extremely dangerous pathogens and that we must develop effective means to suppress and treat the diseases caused by these viruses2.
Currently, the arsenal of approved treatments for diseases caused by coronaviruses is rather limited and therefore there is a pressing need for the discovery and development of therapeutic agents for treatment of COVID-19 and other coronavirus infections3.
Directly acting antiviral agents provide a backbone for treatment of numerous viral disease such as hepatitis B and C and AIDS 4,5 and such a compound, remdesivir, was also very recently FDA approved for emergency treatment of COVID-19 patients.
These therapeutics directly aim at a certain viral protein and, therefore, a deeper understanding of the function of individual viral proteins is needed to derive future therapies of COVID-19 and other possible coronavirus infections.
Coronaviruses have the largest genomes of all RNA viruses. In particular, the genome of SARS-CoV-2 has ~29 800 bases, which encodes 4 structural and 16 nonstructural proteins (nsp1–nsp16) that are essential for the lifecycle of this virus6,7.
SARS-CoV-2 exploits the cell environment to its full advantage for its use and replication8. Importantly, viral RNA must be protected from the cellular innate immunity.
One of the most important elements that ensures the integrity of viral RNA is the cap, a specific arrangement at the 5 ‘end of the RNA molecule that consists of a N-methylated guanosine triphosphate and C2′-O-methyl-ribosyladenine (type 1 cap, Fig. 1).
This arrangement resembles the native mRNA of the host cells, stabilizes the RNA, and ensures effective process of its translation 9,10,11.
In human cells, however, the cap is installed on newly transcribed mRNA already in the nucleus, to which coronaviruses do not have access. Instead, they possess their own cap-synthesizing enzymes.
Clearly, this process is essential for the survival and further replication of viral RNA in cells. In principle, four different processes are necessary for installation of a type 1 cap on RNA (either human mRNA or coronavirus RNA).
First, the γ-phosphate from a 5′-triphosphate end of the nascent RNA is removed by 5′-RNA triphosphatase. Second, a guanosine monophosphate (GMP) is attached to the formed 5′-diphosphate end of RNA by a guanylyltransferase using GTP as the source of GMP. Finally, the methylation steps take place.
In this case, two separate enzymatic steps are required: one for N-7 methylation of the GTP nucleobase (N-7 methyltransferase) and the other for C2′-O methylation of the following nucleotide.
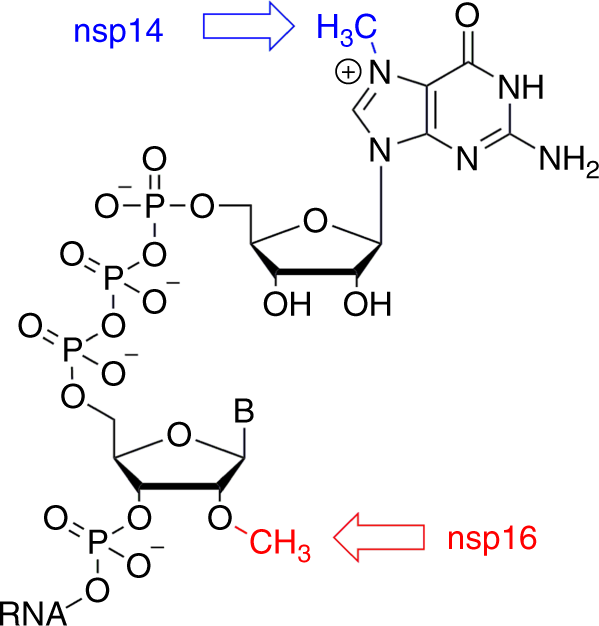
Coronaviruses use sequence installation of the cap that is performed by several nonstructural proteins (nsp) encoded by their genome. For coronaviruses, nsp10, 13, 14, and 16 appear to be involved in this process 12.
The primary function of nsp13 is the unwinding of the viral RNA during replication. Therefore, it is considered to be essentially the helicase. However, it is also a protein with 5′-RNA triphosphatase activity responsible for cleaving monophosphate at the 5′-end of the nascent RNA to provide a diphosphate13.
There is still no clear evidence that any of the coronavirus proteins possess the guanylyltransferase functionality associated with the cap creation12. Nsp14 and nsp16 are responsible for the methylation of the cap on the guanine of the GTP and the C2′ hydroxyl group of the following nucleotide, respectively.
Both nsp14 and nsp16 are S-adenosylmethionine (SAM)-dependent methyltransferases (MTases) and seem to be essential for the viral lifecycle7. In particular, nsp16 appears to be a very promising molecular target from the perspectives of medicinal chemistry and drug design.
It has been shown that this 2′-O methyltransferase (MTase) is indispensable for replication of coronaviruses in cell cultures 14,15. Enzymatic activities of both these MTases (nsp14 and nsp16) are significantly enhanced by nsp10, which is a necessary cofactor for their proper function 15,16,17,18,19.
Here we report on the crystal structure of SARS-CoV-2 nsp10-nsp16 in complex with sinefungin, a pan- MTase inhibitor originally isolated from Streptomyces griseoleus 20.
The structure reveals an overall fold similar to SARS-CoV nsp10-nsp16, and, importantly, reveals atomic details in how sinefungin inhibits the nsp16 MTase. This provides the starting point for specific inhibitor design.
References
1.Coronaviridae Study Group of the International Committee on Taxonomy of V. The species Severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 5, 536–544 (2020).Article Google Scholar
2.Zumla, A., Chan, J. F., Azhar, E. I., Hui, D. S. & Yuen, K. Y. Coronaviruses – drug discovery and therapeutic options. Nat. Rev. Drug Discov. 15, 327–347 (2016).CAS Article Google Scholar
3.Totura, A. L. & Bavari, S. Broad-spectrum coronavirus antiviral drug discovery. Expert Opin. Drug Disco. 14, 397–412 (2019).Article Google Scholar
4.De Clercq, E. A cutting-edge view on the current state of antiviral drug development. Med. Res. Rev. 33, 1249–1277 (2013).Article Google Scholar
5.Martinez, M. G., Villeret, F., Testoni, B. & Zoulim, F. Can we cure hepatitis B virus with novel direct-acting antivirals? Liver Int.: Off. J. Int. Assoc. Study Liver 40 Suppl 1, 27–34 (2020).CAS Article Google Scholar
6.Wu, A. et al. Genome composition and divergence of the novel coronavirus (2019-nCoV) originating in China. Cell host microbe 27, 325–328 (2020).CAS Article Google Scholar
7.Snijder, E. J., Decroly, E. & Ziebuhr, J. The nonstructural proteins directing coronavirus rna synthesis and processing. Adv. virus Res. 96, 59–126 (2016).CAS Article Google Scholar
8.Ziebuhr, J. The coronavirus replicase: insights into a sophisticated enzyme machinery. Nidoviruses: Control Sars Other Nidovirus Dis. 581, 3–11 (2006).CAS Article Google Scholar
9.Bradrick, S. S. Causes and consequences of flavivirus RNA methylation. Front Microbiol. 8, 2374 (2017).Article Google Scholar
10.Dong, H. et al. Flavivirus RNA methylation. J. Gen. Virol. 95, 763–778 (2014).CAS Article Google Scholar
11.Ramanathan, A., Robb, G. B. & Chan, S. H. mRNA capping: biological functions and applications. Nucleic Acids Res. 44, 7511–7526 (2016).Article Google Scholar
12.Chen, Y. & Guo, D. Molecular mechanisms of coronavirus RNA capping and methylation. Virol. Sin. 31, 3–11 (2016).CAS Article Google Scholar
13.Ivanov, K. A. et al. Multiple enzymatic activities associated with severe acute respiratory syndrome coronavirus helicase. J. Virol. 78, 5619–5632 (2004).CAS Article Google Scholar
14.Daffis, S. et al. 2′-O methylation of the viral mRNA cap evades host restriction by IFIT family members. Nature 468, 452–456 (2010).ADS CAS Article Google Scholar
15.Decroly, E. et al. Coronavirus nonstructural protein 16 is a cap-0 binding enzyme possessing (nucleoside-2′O)-methyltransferase activity. J. Virol. 82, 8071–8084 (2008).CAS Article Google Scholar
16.Sawicki, S. G. et al. Functional and genetic analysis of coronavirus replicase-transcriptase proteins. PLoS Pathog. 1, e39 (2005).Article Google Scholar
17.Minskaia, E. et al. Discovery of an RNA virus 3′->5′ exoribonuclease that is critically involved in coronavirus RNA synthesis. Proc. Natl Acad. Sci. USA 103, 5108–5113 (2006).ADS CAS Article Google Scholar
18.Ma, Y. et al. Structural basis and functional analysis of the SARS coronavirus nsp14-nsp10 complex. Proc. Natl Acad. Sci. USA 112, 9436–9441 (2015).ADS CAS Article Google Scholar
19.Decroly, E. et al. Crystal structure and functional analysis of the SARS-coronavirus RNA cap 2′-O-methyltransferase nsp10/nsp16 complex. PLoS Pathog. 7, e1002059 (2011).CAS Article Google Scholar
More information: Structural basis of RNA cap modification by SARS-CoV-2, Nature Communications (2020). DOI: 10.1038/s41467-020-17496-8



{kind=link}