









 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Australian researchers have discovered that removing copper from the blood can destroy some of the deadliest cancers that are resistant to immunotherapy using models of the disease.
While immunotherapy, a treatment that works through a patient’s immune system to kill the cancers, has proven to be a breakthrough for many cancer patients, offering real hope and for some even a cure – some cancers camouflage themselves from current immunotherapies by expressing the aptly titled Programmed Death Ligand or PD-L1.
Dr. Orazio Vittorio and his team from Children’s Cancer Institute in Sydney and UNSW Sydney published the findings today in Cancer Research, a journal of the American Association for Cancer Research.
It is known that cancer cells such as brain cancer “feed” on copper, often having up to six times the normal levels of the metal inside the tumor cells.
Dr. Vittorio and colleagues, including Professor Maria Kavallaris AM, studied tumor samples from more than 90 patients with neuroblastoma and 90 patients with gliomas.
Both these cancers have high mortality rates and to date have not responded well to cancer immunotherapy.
Neuroblastoma accounts for 15% of total childhood cancer deaths and only 50% of patients with high-risk neuroblastoma patient survive their disease. Glioblastoma has the worst survival rate of all cancers, with only 5% of patients surviving 5 years past their diagnosis.
According to Dr. Vittorio, these two cancers express PD-L1 as a way to hide from the immune system, explaining why these two cancers are so deadly.
By looking at the human biopsies the researchers found a correlation between high levels of copper and increased expression of PD-L1. The researchers then showed for the first time that copper levels could control the expression of PD-L1 in cancer cells.
The researchers went on to use an analog of a drug, called TETA, that is currently used in the treatment of Wilson’s Disease, which is a rare genetic disorder characterized by excess copper stored in various body tissues.
They used this drug in animal models of neuroblastoma and glioblastoma to reduce the amount to copper in the tumor cells, leading to a reduction in the expression of PD-L1.
“When these mice were given immunotherapy there was a significant reduction in the size of their tumors,” Dr. Vittorio said.
“Given that TETA is already in use in a number of clinical conditions and it is inexpensive and easy to manufacture, this may offer a viable treatment alternative for those cancers that are resistant to current immunotherapies.”
Neuroblastoma claims more lives of children younger than five than any other cancer. Children like Luciano who was diagnosed at 14 months, endured three operations and eight rounds of chemotherapy.
”We are lucky because he responded well to treatment, but there were so many kids who have been lost. This research will help give hope to more families and children in the future” his mother Maria said.
Tumors occur as a result of the complex interaction between malignant, stromal, immune cells, and vascular system, as these different components communicate with each other via cell–cell contact-dependent mechanisms, soluble messengers and metabolites (1, 2).
It is firmly established that the immune system can be reprogrammed by tumor cells to become ineffective, inactivated, or even acquire a tumor promoting phenotype (3).
In this special tumor microenvironment the macrophages are particularly abundant and play an important role in tumor development by modulating inflammation, immune suppression, and angiogenesis (2).
Many kinds of molecules including growth factors, inflammatory cytokines, chemokines, reactive oxygen, and nitrogen species (ROS and RNS, respectively) from tumor-associated macrophages (TAMs) are involved in the maintenance of a pro-tumorigenic microenvironment and in facilitating metastatic dissemination (3).
Recent evidences have highlighted the metabolic signals as important mediators of macrophage function in the crosstalk between cancer and the immune system (4–6). In this metabolic context, cancer patients are characterized by a variety of perturbations in homeostasis of metal ions such as zinc, iron, selenium, and copper both at intratumoral and systemic level (7, 8).
A large body of preclinical and clinical studies related to dietary deficiencies, indicates that this metal dysregulation triggers neoplastic transformation of cells and/or reduces anti-tumorigenic functions of immune cells by controlling a plethora of chemical and biological reactions (9).
Selenium, copper, zinc, and iron are chemical elements of particular interest given their natural presence in the environment (air, drinking water, and food) and their capacity to stabilize cellular structures, to protect the genome stability, to control metabolic enzymes, receptors, transcription factors at very small concentrations (8, 10).
The purpose of this review is to consider the contribution of these trace elements during neoplastic transformation and their involvement in tumor-induced immune evasion (7).
Here, we will focus on how metal ions modulate TAMs functions in sustaining immune-suppressive environment that protects tumor cell growth or conversely, how the activity of cancer cells influences TAMs via metallic interplay.
New pharmaceutical or dietary intervention strategies with the aim of restoring metal homeostasis, may in the future arise from an improved understanding of “metallic” crosstalk between macrophages and cancer cells.
Copper
Copper is an essential transition metal required for fundamental metabolic processes, but it can be toxic if present in excess (90, 91).
As catalytic cofactor of many enzymes, it is involved in the mitochondrial electron transport chain (cytochrome c oxidase), in the detoxification of reactive oxygen species (superoxide dismutase 1 and 3), in the conversion of hydroperoxides into hydroxides (glutathione peroxidase), in melanin formation (tyrosinase), and in “ferroxidation” (ceruloplasmin) (91).
Copper ions are also fundamental for proteins involved in cell signaling pathways, cell differentiation and death, and for enzymes involved in nervous system physiology. This metal ion plays a crucial role in the development and maintenance of immune function (29, 92).
Indeed copper-deficient patients display decreased numbers of myeloid precursors in the bone marrow and susceptibility to infections (29, 93).
The recommended daily intake of copper in healthy adults is 0.9 mg/day (94).
A reduced intake of copper causes neutropenia, anemia, hypotonia, deterioration of the nervous system, neurodegenerative disorders, and severe intellectual disabilities.
Whereas the overload of copper, mainly in the liver, brain, and kidney, results in redox copper toxicity (e.g., liver cirrhosis) (91, 95). Various studies suggest a strong involvement of altered copper and cupro-proteins levels in cancer (96, 97).
Copper has the ability to catalyze redox reactions and during its dysregulation reactive oxygen species are generated so excessively that act as precursors for neoplastic transformation and metastasis formation (91, 98).
Many types of cancer (brain, multiple myeloma, acute lymphoblastic leukemia, lung, reticulum cell sarcoma, cervical, breast, and stomach cancer) show increased intratumoral levels and/or altered overall distribution of copper (97).
Role in Cancer Cells
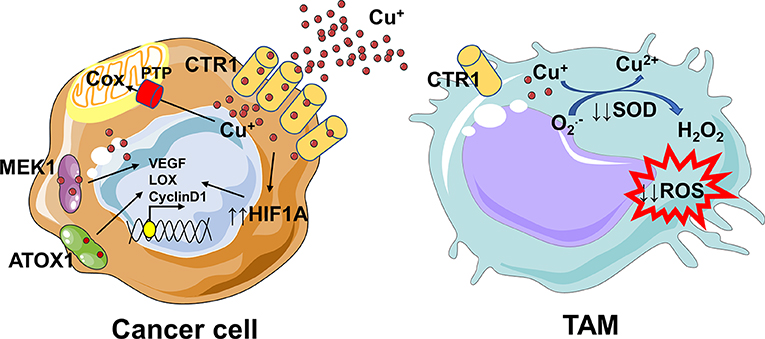
An analysis of the human copper proteome in 18 different tumor types has revealed several copper genes like CTR1, ATOX1, ATP7B, COX17 to be up-regulated (91). The reduced copper (including the dietary pool) is transported inside the cells via CTR1, a high affinity membrane copper transporter.
The increased copper flow via CTR1 is followed by loading onto copper chaperone ATOX1, which acts as a copper-dependent transcription factor promoting the transcription of cyclin D1 and prompting cell replication (91, 99).
Furthermore, copper binds to copper chaperones like COX17 and SCO2, which deliver it to mitochondria and to target proteins involved in trans Golgi network, including ATP7A, and ATP7B (100).
Since copper is essential for the activity of cytochrome c oxidase (Cox), mitochondria rely on the phosphate carrier SLC25A3 for its uptake (101), and on labile copper pool in endoplasmic reticulum as additional source (91, 102).
The mitochondrial phosphate carrier SLC25A3 has been associated with chronic myeloid leukemia progression and might play a role in copper imbalance (103). MEK1 being a copper-binding protein has led to the hypothesis that this metal ion is involved in the RAS-RAF-MEK-ERK pathway, required for cell proliferation, and tumorigenesis (104).
Copper not only binds to proteins directly involved in cancer progression, but also indirectly modulates their expression or activation. Copper inhibits prolyl hydroxylase thus stabilizing HIF-1α and increasing the transcription of various angiogenic genes (e.g., ceruloplasmin and VEGF) (105) and genes involved in the epithelial to mesenchymal transition (e.g., LOX) (91, 106).
The copper-dependent enzyme LOX catalyzes the cross-linking of collagen and elastin in the extracellular matrix (ECM) and interacts with MEMO1 (Mediator of cell Motility 1), another copper-dependent redox enzyme (107). MEMO1 is involved in cell migration through modulation of the cytoskeleton and formation of adhesion sites.
Furthermore, copper ions activate the endothelial Nitric Oxide Synthetase (eNOS), thus increasing the production of the vasodilator nitric oxide (NO) (108). Other studies are required to unveil the mechanisms by which these proteins within the cell are loaded with copper.
The dysregulation of these protein functions could be the priming for processes such as, creation of pre-metastatic niches, escape of immune defense, and angiogenesis. Understanding the mechanism of these genes and protein may open up exciting avenues for developing them as potential cancer therapeutic targets.
Role in Macrophages
Copper is an essential element for immunomodulatory functions (29). As a component of the SOD enzyme, which catalyzes the production of H2O2 from superoxide, it sustains the activity of neutrophils and monocytes, and regulates macrophage antimicrobial functions by contributing to ROS-dependent killing capacity (29, 109).
Indeed its deficiency leads to a defective respiratory burst, impaired phagocytosis, and killing ability, with consequent susceptibility to recurrent pulmonary and urinary infections as well as septicaemia (29, 110, 111).
Macrophages activated with proinflammatory cytokines (IFNγ and TNFα) and LPS show increased copper uptake via CTR1, increased copper accumulation within the phagosomes due to bactericidal Fenton reactions, and finally increased ceruloplasmin activity (112).
The copper-containing ferroxidase ceruloplasmin promotes iron export via FPN, thus starving intracellular bacteria of this essential element (29, 113).
Furthermore, M1-like macrophages display also an increased copper transport to the mitochondria via COX17 for energy production, to SOD1 for antioxidant defense or to Atp7a for protein synthesis (29, 112).
The literature on the role of copper in modulating M2-like macrophages and/or in sustaining TAMs function into tumor microenvironment is absent.
The “Metallic” Cross-Talk Between Macrophages and Cancer Cells
There are not evidences on the copper crosstalk between cancer cells and macrophages, thus in this context we can only speculate. Several studies suggest a strong copper addiction of cancer cells (114, 115), that probably deprives TAMs of this essential element.
Since copper is essential for sustaining the pro-inflammatory phenotype of macrophages (29, 113), its removal from tumor microenvironment could be responsible for a shift toward a pro-tumoral M2-like phenotype and for an immunosuppressive environment (Figure 3).
Overall, our understanding of how copper controls TAMs-cancer cells interplay requires further investigation, with the aim to plan in the future a better dietary intervention or to find novel targets and innovative therapeutic agents.
Opportunities to Improve Cancer Therapy
The strong connection between copper and tumor development, as well as metastization has encouraged scientists to design and synthesize new copper-complexing agents to be used in chemotherapy with lower side effects (79, 91).
The copper-binding compounds used as anticancer agents are divided in two groups: copper chelators, which sequester copper ions from cells, and copper ionophores, which vehicle copper inside cells increasing its intracellular levels and priming cytotoxic effects through multiple pathways (116, 117).
The copper complexing species tetrathiomolybdate (TTM), disulfiram, and clioquinol have been employed in clinical trials, but only TTM has given the most promising results (117). In the latest years, the fact that copper is a limiting factor for multiple phases of tumor progression, has led the scientists to the identification of plant based natural molecules with chelating properties, able to exert antitumoral effects or improve the efficacy of already known drugs, with low side effects (91, 97).
These compounds in the presence of copper act as pro-oxidants and produce reactive oxygen species so excessively to induce DNA degradation (91, 118). The effects of copper, copper oxide nanoparticles, and copper chelate have been evaluated not only on cancer cells but also on macrophages (88, 119).
Chatterjee et al., discovered a novel copper chelate, copper N-(2-hydroxy acetophenone) glycinate (CuNG), able to reprogram TAMs in a proinflammatory type which in turn converts Treg and Th2 cells in anti-tumorigenic Th1 cells (120–122).
This compound triggers in TAMs ROS-mediated activation of MAPKs and ERK1/2 pathways which lead to upregulation of IL-12 and simultaneous downregulation of TGF-β and IL-10 production (121).
We may speculate on a bivalent role of these redox-active compounds like CuNG in a clinical approach. The sustained generation of ROS on the one hand would induce apoptosis of cancer cells, on the other hand would trigger proimmunogenic macrophages.
REFERENCE LINK : https://www.frontiersin.org/articles/10.3389/fonc.2020.00646/full
More information: Florida Voli et al. Intra-tumoral copper modulates PD-L1 expression and influences tumor immune evasion., Cancer Research (2020). DOI: 10.1158/0008-5472.CAN-20-0471



{kind=link}