











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



La sindrome respiratoria acuta grave coronavirus 2 (SARS-CoV-2) sta causando la nuova pandemia di malattia da coronavirus del 2019 ( COVID-19 ) che ha infettato più di 5 milioni di persone e ucciso più di 350.000 persone in tutto il mondo.
L’indagine sistematica dei farmaci approvati clinicamente è una priorità per migliorare gli esiti della malattia e investire risorse per passare alla produzione su vasta scala.
La ricerca di una terapia efficace è in corso attivamente, ma attualmente ha un successo limitato. Forse dovremmo guardare fuori dagli schemi e considerare ciò che è nascosto in bella vista, come le ragioni biologiche per cui le donne sono relativamente protette dal COVID-19 rispetto agli uomini.
Questa recensione evidenzia prove sperimentali che gli ormoni steroidei 17β-estradiolo (E2) e progesterone (P4), ad alte concentrazioni fisiologiche, sono potenti immunomodulatori e sostiene che la terapia steroidea acuta con la combinazione di E2 e P4 può rappresentare un’opzione terapeutica sicura e praticabile che deve essere testato in studi clinici per mitigare gravi esiti COVID-19.
Malattia da coronavirus – 2019 La mortalità è inferiore nelle donne rispetto agli uomini
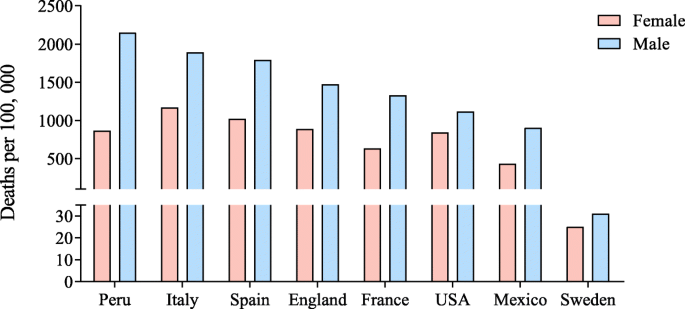
Dall’inizio del 21 ° secolo, 2 precedenti epidemie mortali di betacoronavirus zoonotici hanno superato le barriere delle specie per infettare gli esseri umani e hanno mostrato la stessa apparente protezione femminile da esiti gravi.
La prima epidemia di SARS-CoV è emersa nel 2002 nella provincia del Guangdong, in Cina, e tra 1755 pazienti ricoverati a Hong Kong il tasso di mortalità era del 13% nelle donne rispetto al 22% negli uomini (1). Durante l’epidemia di coronavirus (MERS-CoV) in corso in Medio Oriente, iniziata nel 2012 in Arabia Saudita, tra i 425 casi segnalati, l’insorgenza della malattia è stata inferiore tra le donne (38% dei casi) e il tasso di mortalità è stato del 23% per le donne rispetto al 52% per gli uomini (2).
Oggi in Cina, Europa e Stati Uniti, la gravità e la mortalità del COVID-19 è costantemente inferiore nelle donne rispetto agli uomini (3-8).
Prendendo le serie più rappresentative fino ad oggi, nella coorte di 1099 pazienti ospedalizzati COVID-19 a Wuhan, in Cina, solo il 42% dei pazienti erano donne (4). Tra i casi gravi (cioè, ricoverati in un’unità di terapia intensiva, che richiedono ventilazione meccanica, o fatali), le donne rappresentavano il 32% dei pazienti (4).
Allo stesso modo, le donne rappresentavano solo il 18% di tutti i ricoveri di COVID-19 nelle unità di terapia intensiva nella regione Lombardia (9). A New York City, su 5700 pazienti ricoverati, le donne rappresentavano il 33% dei casi e il 39% dei decessi (7).
L’International Severe Acute Respiratory and Emerging Infections Consortium (ISARIC) in uno studio prospettico osservazionale di coorte su oltre 17000 pazienti nel Regno Unito ha riferito che tra i pazienti ospedalizzati, le donne rappresentavano solo il 40%, con una mortalità inferiore del 20% rispetto agli uomini (10).
Sebbene l’avanzare dell’età sia associato a un maggior rischio di mortalità in entrambi i sessi, la protezione femminile rimane evidente (11). Un’analisi dei dati COVID-19 in Italia, Spagna, Germania, Svizzera, Belgio e Norvegia rivela che tra tutti i gruppi di età superiore ai 20 anni, i tassi di mortalità sono maggiori per i maschi rispetto alle femmine (12).
Al contrario, le differenze maschi-femmine nel tasso di infezioni confermate da SARS-CoV-2 dipendono dall’età in tutti i paesi, essendo maggiori tra le donne di età compresa tra 10 e 50 anni e maggiori tra i maschi di età inferiore a 10 anni e di età superiore a 50 anni (12) .
Interpretiamo questi dati per suggerire che le differenze sessuali biologiche contribuiscono alla protezione di parte femminile contro la morte, ma il rischio di esposizione associato al sesso può influenzare i tassi di infezione in modo diverso per maschi e femmine in età diverse.
Sorge quindi una domanda su quali fattori biologici sono protettivi nelle donne rispetto agli uomini, e come possiamo sfruttare questi fattori modificabili per mitigare la morbilità e la mortalità COVID-19 ?
Ormoni sessuali e infezioni virali
Gli ormoni gonadici non solo sono coinvolti nella differenziazione degli organi riproduttivi, ma esercitano anche una regolazione specifica del sesso su più tessuti, incluso il cervello e quelli del sistema immunitario [52a, 72a, 73a].
Negli esseri umani, la costituzione del cromosoma sessuale (XX / XY) determina il sesso e il gene della regione determinante il sesso Y (SRY) presente sul cromosoma Y è il gene regolatore principale della differenziazione sessuale [74a]. SRY guida la differenziazione del testicolo attivando i geni a valle e la cascata di attivazione genica innesca la gonade a secernere testosterone.
Il testosterone secreto aiuta ulteriormente a differenziare i sistemi riproduttivi maschili. Si suggerisce anche che il testosterone raggiunga il cervello e organizzi le reti neuronali. Il testosterone può mediare l’espressione genica direttamente legandosi al recettore degli androgeni (AR) o indirettamente dopo la conversione in estrogeni da parte dell’enzima aromatasi [73a, 75a].
D’altra parte, estrogeni e progesterone sono ormoni importanti nelle donne che portano alla regolazione differenziale del sistema riproduttivo e immunitario [51a].
Esistono tre diversi tipi di estrogeni prodotti nelle femmine: estrone (E1), 17β-estradiolo (E2) ed estriolo (E3). E2 è la forma predominante prodotta dalle ovaie e il livello degli ormoni oscilla durante l’ovulazione e la gravidanza. Gli estrogeni agiscono attraverso i recettori degli estrogeni (ER) che esistono in due forme, ERα e ERβ [76a].
L’espressione di ERα ed ERβ è stata identificata nelle cellule immunitarie umane, inclusi linfociti B e T, mastociti, macrofagi, cellule dendritiche, monociti e cellule natural killer [77a, 78a, 79a].
È stato dimostrato che l’espressione di ERs è cellula specifica poiché ERα è risultato essere la forma predominante nelle cellule T CD4 + e ERβ era la forma predominante nelle cellule B [78a].
L’espressione di ERα specifica per sesso ed età è stata identificata nei monociti umani con una maggiore espressione nelle femmine e nei maschi in post-menopausa rispetto alle femmine in pre-menopausa [78a].
Tuttavia, l’espressione ER dipendente dal sesso non è stata osservata nei linfociti B e T. Poiché non vi era alcuna differenza nell’espressione di ER nei linfociti T e B maschili e femminili, gli autori hanno sostenuto che le differenze di sesso nella risposta immunitaria potrebbero non essere un effetto diretto degli estrogeni ma potrebbero essere indirette attraverso l’ormone di rilascio delle gonadotropine [78a].
Nakada et al. l’utilizzo di un modello murino ha mostrato che i livelli di ERα RNA erano più alti nelle cellule staminali ematopoietiche maschili (HSC) rispetto alle HSC femminili e che il livello di ERβ era basso sia nelle HSC maschili che femminili.
Gli autori hanno anche scoperto che le HSC nelle femmine si dividono più frequentemente che nei maschi. È interessante notare che la delezione condizionale di ERα ha determinato una ridotta proliferazione di HSC nelle femmine ma non nei maschi [80a].
Maschi e femmine sono sotto l’influenza di diversi ambienti ormonali.
È stato dimostrato che il testosterone ha un effetto immunosoppressivo mentre gli estrogeni hanno un effetto immunosoppressore [60a].
È stato scoperto che il testosterone inibisce la differenziazione delle cellule T helper [81a] e si correla positivamente con la carica virale del virus dell’encefalite equina venezuelana nei macachi [82a].
È noto anche che il testosterone riduce la reattività al vaccino antinfluenzale [83]. L’analisi dei livelli di testosterone nelle femmine ha mostrato che il livello è solitamente basso nelle femmine affette da malattie autoimmuni rispetto alle femmine sane [55a].
Inoltre, l’ablazione degli androgeni nei topi maschi ha mostrato un miglioramento delle prestazioni delle cellule immunitarie nei confronti del cancro alla prostata [84a]. L’ablazione degli androgeni altera anche gli organi immunitari poiché è stato riportato che il timo e la milza aumentano il peso [85a].
Sebbene la popolazione di cellule T CD4 periferiche mature sia diminuita, quella che ha raggiunto la milza ha mostrato una maggiore attivazione [85a]. Ciò suggerisce che gli ormoni sessuali maschili (androgeni) potrebbero portare a suscettibilità e gravità alle infezioni patogene.
È indicato che i pazienti affetti da cancro alla prostata che sono sotto terapia di deprivazione androgenica (ADT) per regolare la produzione di androgeni hanno ridotto il rischio di infezione da SARS-CoV-2 rispetto ai pazienti che non hanno ricevuto ADT [86a].
Gli autori hanno sostenuto che [86a] poiché l’espressione di TMPRSS2 è indotta da androgeni [87a], l’ADT potrebbe sottoregolare TMPRSS2 e questo a sua volta potrebbe abbassare l’infezione da SARS-CoV-2.
Si suggerisce che l’ADT potrebbe essere utile per COVID-19 e poiché questa malattia progredisce rapidamente, l’intervento di ADT sarà utile durante la fase iniziale dell’infezione virale e non nelle fasi successive [88a].
Tuttavia, l’espressione di TMPRSS2 nei polmoni umani maschili e femminili non è diversa [89a, 90a] e nei modelli di topo, il trattamento con enzalutamide, un antagonista dell’AR, non ha prodotto una diminuzione dell’espressione polmonare di TMPRSS2 [89a]. Quindi, l’uso di antagonisti AR per regolare l’espressione di TMPRSS2 per COVID-19 garantisce ulteriori ricerche.
L’estrogeno, d’altra parte, fornisce protezione contro i patogeni in quanto ha proprietà antivirali in diverse infezioni virali tra cui HIV, virus dell’epatite C, Ebola e citomegalovirus umano [91a]. È stato dimostrato che gli estrogeni inibiscono la replicazione del virus dell’influenza A nelle cellule epiteliali nasali in coltura isolate da topi femmina [91a].
È interessante notare che l’effetto protettivo degli estrogeni non è stato osservato nelle cellule epiteliali nasali isolate da topi maschi [91a]. Poiché la SARS si replica prevalentemente nelle vie aeree, livelli più elevati di estrogeni nelle femmine possono aumentare la protezione dalle infezioni da SARS [63a].
L’inibizione della funzione ER utilizzando ICI 182.780 antagonista ER ha provocato infezioni SARS-CoV più elevate nelle femmine. Tuttavia, la gonadectomia o il trattamento con l’antagonista AR flutamide non ha influenzato la morbilità o la mortalità nei topi maschi a seguito di infezione da SARS-CoV [63a].
Sulla base di ciò, è stato suggerito che la segnalazione del recettore degli estrogeni gioca un ruolo importante nell’infezione e nella mortalità da coronavirus, mentre gli androgeni non svolgono un ruolo nella patogenesi [63a]. Ciò suggerisce che la segnalazione degli estrogeni è fondamentale nella regolazione dell’infezione virale e potrebbe essere uno dei motivi per cui le femmine mostrano un rapido recupero e una bassa mortalità da COVID-19.
Testicolo e clearance virale
Il testicolo è un organo immuno-privilegiato poiché sia gli allo che gli autoantigeni non sono in grado di provocare una risposta immunitaria. Questa caratteristica è importante per mantenere le cellule germinali immunogeniche lontane dalla risposta immunitaria [92a].
Il sistema immunitario non controllato può reagire con l’antigene di superficie sugli spermatozoi noto come antigene meiotico delle cellule germinali (MGCA), e questo può provocare infertilità [92a]. Sebbene il testicolo sia un organo immuno-privilegiato, può suscitare un’immunità innata quando i patogeni microbici si infiltrano nell’organo.
È noto che i virus inclusi l’HIV, il citomegalovirus e la parotite infettano i testicoli e provocano disturbi ai testicoli [93a]. Inoltre, virus inclusi Zika, Ebola e Marburg sono stati isolati da campioni di sperma e si sa che sono trasmessi sessualmente [94a].
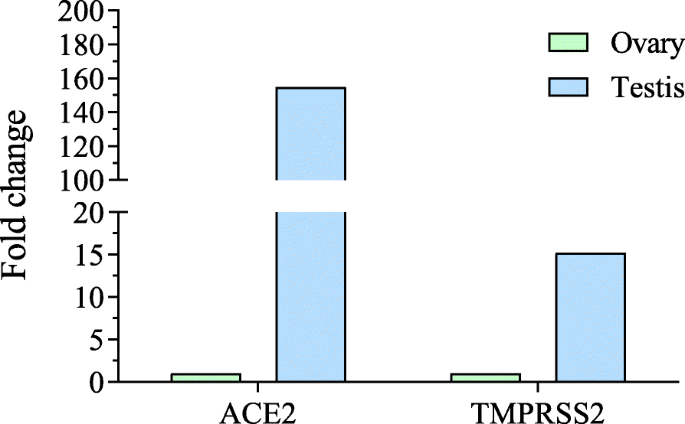
Shastri e colleghi hanno dimostrato che i maschi nelle famiglie richiedono più tempo per riprendersi dal COVID-19 rispetto agli altri membri della famiglia. Gli autori hanno osservato che i testicoli avevano un’alta espressione di ACE2 sia a livello di mRNA che di proteine [26a].
Gli autori hanno suggerito che il coronavirus può entrare nel testicolo e di conseguenza portare a una maggiore carica virale e richiedere più tempo per la clearance virale. I soggetti totali in questo studio erano solo 68 (48 maschi e 20 femmine) con un’età media di 37 anni. Per supportare l’ipotesi dell’autore è necessario uno studio approfondito con un numero maggiore di pazienti.
In un altro studio, il materiale genetico del coronavirus è stato rilevato nei campioni di sperma di maschi infettati da coronavirus [95a]. L’analisi dei dati di sequenziamento dell’RNA ottenuti dal database NCBI [96] ha mostrato che sia ACE2 che TMPRSS2 erano altamente espressi nel testicolo rispetto all’ovaio (Fig. 4).
Ciò supporta l’osservazione che il coronavirus potrebbe entrare nel testicolo umano. Anche gli ormoni sessuali nei maschi sono stati alterati a seguito dell’infezione da SARS-CoV-2. L’analisi di 81 uomini infetti con SARS-CoV-2 ha mostrato un aumento dell’ormone luteinizzante (LH), ma il rapporto tra testosterone e LH e ormone follicolo-stimolante con LH era diminuito [97a].
Sebbene l’RNA del coronavirus sia stato rilevato nei campioni di sperma, le particelle virali attive non sono state finora isolate dai testicoli. Quindi, la teoria del coinvolgimento del testicolo nella clearance virale ritardata e nell’elevata mortalità negli uomini [26a] dovrebbe essere presa con cautela.
Il disturbo della regolazione immunitaria nel testicolo può provocare l’orchite, una condizione in cui i leucociti si infiltrano nel testicolo e danneggiano i tubuli seminiferi e di conseguenza provocano infertilità. Quindi, se il testicolo è un fattore che contribuisce alla prognosi sfavorevole e all’elevata mortalità negli uomini deve essere esaminato in modo più approfondito.
Nell’epidemia di SARS nel 2002, è stata osservata una mortalità più elevata nei maschi rispetto alle femmine.
Gli individui infetti hanno mostrato più danni d’organo e nei maschi, il testicolo è stato anche affetto da distruzione delle cellule germinali, mancanza di spermatozoi, membrana basale ispessita e infiltrazione leucocitaria [98a]. Tuttavia, non sono state rilevate particelle virali o RNA virale nei campioni di tessuto ed è stato suggerito che il danno del testicolo mediato dal virus fosse dovuto a una risposta immunitaria.
Ruolo della tempesta di citochine proinfiammatorie nella malattia da coronavirus – Risultati del 2019
Gli esiti gravi di COVID-19 sono associati a risposte immunitarie innate ritardate ed esagerate, tra cui ipercitochina e infiltrazione di cellule infiammatorie nei polmoni. La nostra attuale comprensione della malattia, che è in rapida evoluzione mentre scriviamo questa recensione, è che i pazienti con COVID-19 non muoiono per i danni causati dalla replicazione del virus, muoiono per le conseguenze di una cosiddetta “tempesta di citochine” (13 -16).
Nel tentativo di proteggere il corpo dalla SARS-CoV-2, le cellule immunitarie si infiltrano nei polmoni, causando iperattivazione di monociti e macrofagi e un’elevata produzione di citochine proinfiammatorie (p. Es., Interleuchina-6 [IL-6], interleuchina-1β [IL -1β], fattore di necrosi tumorale α [TNF-α]) e chemochine (p. Es., Monocyte chemoattractant protein-1 (MCP-1 / CCL2]) (15).
La tempesta di citochine è anche associata a linfopenia e uno studio su 21 pazienti di Wuhan ha riportato una diminuzione delle cellule T CD4 + e CD8 +, nonché una produzione soppressa di interferone γ da parte delle cellule T CD4 +, che era associata alla gravità COVID-19 (15) .
L’effusione locale di chemochine e citochine attrae più cellule infiammatorie, come neutrofili e monociti, nel tessuto polmonare, provocando lesioni polmonari. Ironia della sorte, la tempesta di citochine è il risultato del sistema immunitario che risponde alle infezioni nel tentativo di proteggere l’ospite, ma si traduce in sindrome da distress respiratorio acuto e insufficienza multiorgano (13, 14).
Si ipotizza che l’aumento della produzione e l’IL-6 locale e sistemica elevata siano centrali per lo sviluppo della tempesta di citochine (17, 18).
Di conseguenza, strategie terapeutiche mirate alla risposta infiammatoria come il blocco di IL-6 (19) o il trapianto di cellule staminali mesenchimali per ripristinare la tolleranza immunitaria (20) stanno mostrando risultati preliminari promettenti per mitigare la tempesta di citochine. Qui discutiamo un paradigma in cui la terapia con gli ormoni steroidei E2 e P4 potrebbe mitigare questa risposta immunitaria infiammatoria innata viralmente indotta.
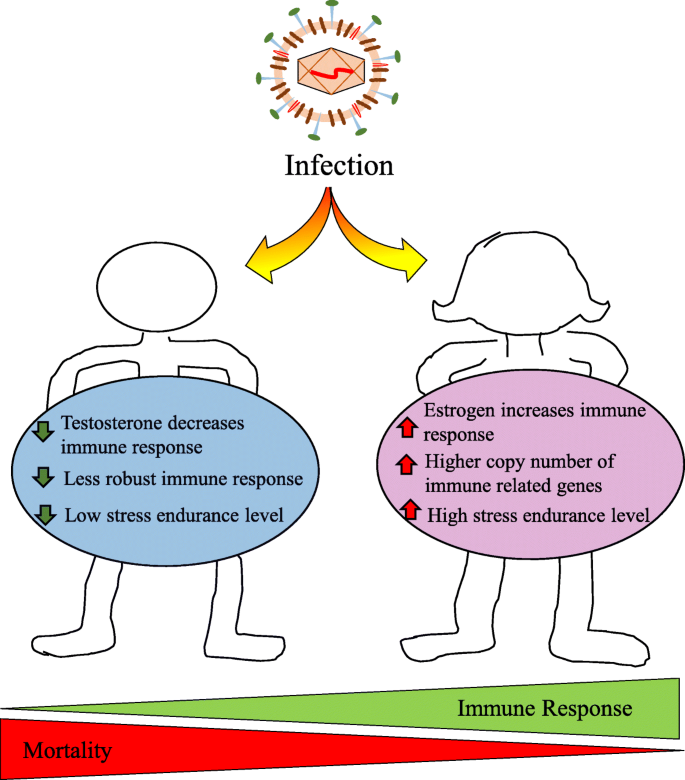
Le femmine generalmente mostrano una maggiore risposta immunitaria ai virus
Le femmine generalmente sviluppano risposte immunitarie più elevate rispetto ai maschi.
Nel 1967, Butterworth et al hanno riferito che le donne producono livelli più elevati di immunoglobuline IgG e IgM circolanti rispetto agli uomini (21), il che è stato successivamente confermato da più studi. Di conseguenza, a seguito della vaccinazione contro l’influenza, la febbre gialla, la rosolia, il morbillo, la parotite, l’epatite, l’herpes simplex 2, la rabbia, il vaiolo e i virus della dengue, le risposte anticorpali protettive sono due volte più elevate nelle donne rispetto agli uomini (22).
Le donne hanno anche frequenze più elevate di cellule T helper CD4 + rispetto agli uomini (23).
Le ragioni biologiche per cui le femmine sviluppano una risposta immunitaria più robusta rispetto ai maschi contro i patogeni, compresi i virus, probabilmente spiegano la protezione femminile osservata dagli esiti fatali del COVID-19. In primo luogo, le femmine godono del vantaggio genetico di 2 cromosomi X ed essendo un mosaico di geni legati all’X (cioè, che esprimono casualmente alleli ereditati dalla madre o dal padre), inclusi più di 60 geni di risposta immunitaria (24).
Al contrario, i maschi hanno un solo cromosoma X ereditato dalla madre. Diversi studi dimostrano che le malattie genetiche associate ad alleli deleteri legati all’X sono più frequentemente osservate nei maschi (25).
In generale, non dovrebbe esserci alcun effetto del dosaggio associato alla posizione di 2 cromosomi X nelle femmine. L’inattivazione incompleta dei geni immunoregolatori sul cromosoma X nelle femmine, tuttavia, può causare uno squilibrio del dosaggio genico tra i sessi (26, 27), che è implicato nelle malattie autoimmuni di parte femminile (28) e nell’efficacia del vaccino (29).
Il cromosoma Y ha anche funzioni immunoregolatrici (30) che sono collegate agli esiti dell’influenza, almeno nei topi (31).
Gli steroidi sessuali sono potenti immunomodulatori e le diverse concentrazioni di estrogeni, P4 e androgeni tra donne e uomini, oltre alla genetica descritta in precedenza, possono influenzare le risposte immunitarie COVID-19 e gli esiti infiammatori.
Ciò è particolarmente importante perché malattie acute e gravi, come COVID-19, possono alterare la funzione dell’asse gonadico ipotalamo-ipofisario e diminuire la produzione endogena di estrogeni e P4.
Gli ormoni sono anche suscettibili di intervento terapeutico. Successivamente, discuteremo l’immunomodulazione fornita da elevate concentrazioni sieriche fisiologiche di estrogeni e P4 in relazione all’infezione da SARS-CoV-2. Questa conoscenza di base è fondamentale per apprezzare i potenziali benefici che il trattamento E2 e P4 potrebbe fornire nel contesto dell’iperinfiammazione mediata da SARS-CoV-2 e della sindrome da distress respiratorio acuto.
Estrogeni, progesterone e funzione immunitaria
I recettori degli estrogeni (ER) sono espressi in tutte le cellule immunitarie, fungendo da regolatori trascrizionali della funzione cellulare.
Nelle cellule mononucleate del sangue periferico umano, i linfociti T CD4 + esprimono livelli più elevati di RNA messaggero ERα rispetto a ERβ, mentre le cellule B esprimono livelli più elevati di ERβ rispetto all’RNA messaggero ERα (32).
Le cellule T CD8 + del sangue periferico ei monociti esprimono livelli bassi ma comparabili di entrambi gli ER (32). La terapia con E2, che porta a concentrazioni sieriche equivalenti all’ovulazione o alla gravidanza, possiede azioni immunomodulatorie e antinfiammatorie benefiche nei topi e nell’uomo (rivisto in [24, 33]).
Nella maggior parte dei modelli sperimentali umani o di roditori, le azioni antinfiammatorie di E2 sull’immunità innata includono la soppressione della produzione di citochine proinfiammatorie, ad esempio IL-6, IL-1β e TNF-α, da parte di monociti e macrofagi (a fattore principale nella tempesta di citochine COVID-19) e una forte inibizione di CCL2, prevenendo così la migrazione delle cellule immunitarie innate nelle aree infiammate, in particolare neutrofili e monociti.
E2 stimola la produzione di cellule T helper CD4 + di citochine antinfiammatorie, ad esempio interleuchina 4 (IL-4), interleuchina 10 (IL-10) e interferone γ.
In generale, concentrazioni elevate di E2 favoriscono le risposte antinfiammatorie di tipo T helper di tipo 2 (Th2). L’E2 riduce la produzione di interleuchina 17 da parte delle cellule helper Th17 proinfiammatorie. E2 aumenta l’espansione delle cellule T regolatorie (Treg) promuovendo così la tolleranza immunitaria. E2 stimola anche la produzione di anticorpi da parte delle cellule B (Figura 1).

Azione antinfiammatoria e immunomodulante dell’estradiolo e del progesterone. Elevate concentrazioni fisiologiche di 17β-estradiolo (E2) sopprimono la produzione di citochine proinfiammatorie, ad esempio interleuchina 6 (IL-6), interleuchina 1β (IL-1β) e fattore di necrosi tumorale α (TNF-α) e chemochina CCL2 dai macrofagi, impedendo così la migrazione di neutrofili e monociti nelle aree infiammate. Il progesterone (P4) inibisce anche la produzione di citochine proinfiammatorie IL-1β e interleuchina 12 da parte dei macrofagi e delle cellule dendritiche. Alte concentrazioni di E2 o P4 stimolano la produzione di cellule T helper CD4 + di citochine antinfiammatorie, ad esempio interleuchina 4 e interleuchina 10, e favoriscono risposte antinfiammatorie di tipo Th2. E2 e P4 migliorano anche l’espansione delle cellule T regolatorie (Treg), promuovendo così la tolleranza immunitaria. Finalmente,
C’è una forte evidenza nella malattia ossea metabolica e nell’epatopatia indotta da virus che gli estrogeni inibiscono la patogenesi della malattia attraverso la soppressione della produzione di IL-6.
Ad esempio, gli estrogeni inibiscono lo sviluppo degli osteoclasti e la funzione di riassorbimento nell’osso inibendo la trascrizione e la produzione del gene IL-6 (34).
Inoltre, l’incidenza del carcinoma epatocellulare (HCC) indotto da epatite B cronica nell’uomo mostra una forte predominanza maschile.
Si ritiene che l’IL-6 sia un componente chiave nella tumorigenesi associata all’infiammazione dell’HCC (35).
In uno studio retrospettivo su donne in postmenopausa con epatite cronica C, la progressione verso la fibrosi epatica è stata ridotta nelle donne che hanno assunto la terapia con estrogeni in menopausa, rispetto alle donne che non l’hanno fatto (36). In un modello di ratto di HCC indotto chimicamente, i maschi hanno prodotto più IL-6 dalle cellule di Kupffer del fegato ed erano più inclini a HCC rispetto alle femmine (37). Gli estrogeni proteggevano i maschi dall’HCC tramite l’inibizione della produzione di IL-6 da parte delle cellule di Kupffer.
In un modello murino di infiammazione polmonare acuta mediante instillazione di lipopolisaccaride batterico, i maschi e le femmine ovariectomizzate hanno mostrato una maggiore infiltrazione polmonare di cellule polimorfonucleate con elevata produzione di IL-6, IL-1β e ICAM-1 (molecola di adesione intercellulare-1), che è stato ridotto dal trattamento E2 di maschi e femmine ovariectomizzate (38).
Nei modelli preclinici di infezione influenzale, gli estrogeni mostrano potenti azioni immunomodulatorie che portano a una risposta immunitaria innata più appropriata nei polmoni, che è associata a una diminuzione delle citochine proinfiammatorie e delle risposte chemochine prima che si sviluppi la malattia clinica (39-41).
Nelle colture primarie di cellule epiteliali nasali umane, i composti estrogenici, incluso E2, che segnalano attraverso ERβ riducono significativamente la replicazione del virus influenzale (42). Inoltre, SARS-CoV-2 e SARS-CoV producono entrambi polmoniti mortali con la stessa apparente protezione femminile.
In un modello murino di infezione da SARS-CoV, topi femmine hanno sviluppato titoli di virus inferiori, minore infiltrazione con monociti infiammatori, macrofagi e neutrofili che producono meno citochine infiammatorie (IL-6, IL-1β e TNF-α) e chemochine (CCL2) , con conseguente danno polmonare più lieve e una mortalità femminile inferiore (20%) rispetto ai maschi (80%) (43), una distribuzione per sesso simile a quella osservata nei pazienti con SARS.
È importante sottolineare che la produzione endogena di E2 nelle femmine di topo è stata determinante in questa protezione. La castrazione dei maschi non ha avuto alcun effetto sulla malattia, mentre la rimozione chirurgica delle ovaie o il trattamento con fulvestrant, antagonista del pronto soccorso, in topi femmine infettati da SARS-CoV ha prodotto gli stessi danni polmonari e tasso di mortalità dei maschi.
Inoltre, il trattamento di topi ovariectomizzati con il modulatore selettivo del recettore degli estrogeni (SERM) approvato dalla Food and Drug Administration (SERM) tamoxifene, un agonista misto dell’ER e un antagonista prescritto per il trattamento del cancro al seno, ha ripristinato la protezione femminile.
Questo studio indica che in un modello murino di infezione da SARS-CoV, gli ormoni ovarici e soprattutto gli estrogeni proteggevano le femmine dalla polmonite letale e il tamoxifene imitava la protezione di parte femminile. Uno screening di più composti approvati dalla Food and Drug Administration per l’attività anticoronavirus ha identificato il tamoxifene e il toremifene (un altro SERM) tra i primi 10 farmaci più efficaci e sicuri nell’inibire le infezioni da MERS-CoV e SARS-CoV in vitro (44).
Il toremifene inibisce anche l’infezione da virus Ebola in vitro e in vivo nei topi (45). Il meccanismo d’azione del toremifene sembra correlato alla struttura anfifila cationica multipla della molecola che ostacola la fase tardiva dell’ingresso o della fusione del virus.
Presi insieme, questi risultati suggeriscono che E2 e SERM correlati hanno 2 potenziali meccanismi di azione protettiva contro le polmoniti mediate da SARS-CoV nei topi:
1) una diminuzione dipendente dagli estrogeni nella risposta immunitaria innata mortale e tempesta di citochine nei polmoni, prevenendo così l’insufficienza respiratoria, e
2) specifico per SERM, un’inibizione diretta off-target della replicazione di SARS-CoV e degli effetti citopatici.
Il P4 è un altro importante ormone immunomodulatore e antinfiammatorio prodotto ad alti livelli dalla placenta durante la gravidanza. I recettori del progesterone sono espressi nella maggior parte delle cellule immunitarie, comprese le cellule epiteliali, i macrofagi, le cellule dendritiche, i linfociti, i mastociti e gli eosinofili (24).
Tuttavia, P4 può anche segnalare tramite i recettori glucocorticoidi e mineralcorticoidi. P4 inibisce la produzione di citochine proinfiammatorie IL-1β e interleuchina 12 da parte di macrofagi umani e roditori e cellule dendritiche. Il progesterone favorisce l’inclinazione delle risposte delle cellule T helper CD4 + dal tipo Th1 al tipo Th2 e la produzione di citochine anti-infiammatorie IL-4 e IL-10 (24, 46, 47).
Il trattamento delle cellule del sangue del cordone ombelicale con P4 aumenta la percentuale di cellule FOXP3 + Treg (promuovendo così la tolleranza immunitaria), mentre diminuisce le frequenze delle cellule Th17 proinfiammatorie (Fig.1). La somministrazione di P4 a concentrazioni che imitano la fase luteale a topi femmine adulte prive di progesterone ha conferito protezione dalla polmonite letale del virus dell’influenza A (48).
In questi topi, il trattamento con P4 ha ridotto l’ambiente infiammatorio dei polmoni, migliorato la funzione polmonare e promosso la proliferazione cellulare e la riparazione polmonare, che hanno portato a un recupero più precoce, senza effetti sulla carica virale.
È interessante notare che, in questo caso, il trattamento con P4 ha promosso un recupero più rapido aumentando il fattore di crescita trasformante β, IL-6, l’interleuchina 22 e il numero di cellule Th17 regolatorie che esprimono CD39.
È importante sottolineare che il progesterone ha promosso la riparazione del tessuto polmonare sovraregolando il fattore di crescita epidermico amfiregulina nei polmoni (48).
Sebbene l’infezione da virus dell’influenza A sia diversa e produca una reazione immunitaria diversa da quella indotta da SARS-CoV-2 (p. Es., L’effetto benefico di IL-6), questo studio fornisce importanti informazioni sugli effetti immunomodulatori e curativi di P4. Inoltre, P4 sembra anche mostrare attività antivirale nelle cellule VeroE6 infettate con SARS-CoV-2 (49).
Gravidanza e malattia da Coronavirus – 2019
Durante la gravidanza, le risposte immunitarie innate e adattive passano da un fenotipo infiammatorio a uno antinfiammatorio per evitare il rigetto fetale e favorire il trasferimento passivo di anticorpi materni al feto (rivisto in [50]).
Questi effetti, che sono rilevanti per la protezione COVID-19, sono ampiamente mediati da E2 e P4. Durante la gravidanza, livelli aumentati di E2 sopprimono molte risposte immunitarie infiammatorie citotossiche e innate ma stimolano la produzione di anticorpi da parte delle cellule B (33, 51).
In effetti, una delle caratteristiche immunologiche più importanti della gravidanza è l’aumento delle risposte delle cellule B con una maggiore produzione di anticorpi dovuta alla doppia stimolazione da parte di estrogeni e P4, la cui produzione è massima nel terzo trimestre (33, 51).
P4 stimola anche la sintesi del fattore di legame indotto dal progesterone (PIBF) da parte dei linfociti, che promuove la differenziazione delle cellule T CD4 + in cellule Th2 che secernono citochine antinfiammatorie, tra cui IL-4, interleuchina 5 e IL-10 (52).
Questo spiega perché durante la gravidanza le malattie causate da cellule B / anticorpi, come il lupus eritematoso sistemico, si aggravano; mentre le malattie guidate dalle cellule T con risposte immunitarie citotossiche e innate, come l’artrite reumatoide o la sclerosi multipla, migliorano (50, 53).
Le donne incinte non sono protette dall’infezione da SARS-CoV-2, ma sembrano essere relativamente protette dagli esiti gravi di SARS-CoV-2. Attualmente, gli studi che valutano gli esiti di COVID-19 durante la gravidanza non hanno ancora separato gli esiti che si verificano durante la gravidanza (cioè, quando le concentrazioni di E2 e P4 sono elevate) da quelli nell’immediato periodo postpartum (cioè, quando le concentrazioni di E2 e P4 non sono rilevabili).
In una serie retrospettiva cinese di 82 donne (28 donne incinte, 54 donne non gravide in età riproduttiva) ricoverate in ospedale a Wuhan con COVID-19 confermato, le donne incinte hanno mostrato gravità della malattia, tempo di eliminazione del virus e durata della degenza ospedaliera comparabili rispetto a quelli riproduttivi. donne anziane non gravide (54).
Gli autori hanno concluso che le donne incinte infettate da SARS-CoV-2 hanno decorso clinico e risultati comparabili rispetto alle donne di controllo. Tuttavia, in questo studio, le donne non gravide hanno ricevuto più terapie antivirali, corticosteroidi e immunoglobuliniche rispetto alle donne in gravidanza e quindi i gruppi non erano comparabili in termini di trattamenti e risultati correlati.
Una più ampia revisione retrospettiva di 118 donne incinte ammesse per polmonite COVID-19 in Cina ha riportato solo 9 casi (8%) di polmonite grave con ipossiemia. In particolare, in 6 di queste donne, inclusa 1 che necessitava di ventilazione meccanica, l’esacerbazione della polmonite si è verificata durante il periodo postpartum, dopo che le concentrazioni sieriche di E2 e P4 erano già diminuite (55).
Pertanto, il numero effettivo di casi gravi in questo studio era 3 (2,5% della popolazione incinta), che è inferiore alla gravità del COVID-19 nelle donne cinesi in una fascia di età simile (circa il 6%) (2).
Infatti nell’unica serie pubblicata di 9 donne in gravidanza con COVID-19 fatale, un’analisi dettagliata dei casi rivela che 7 di queste donne sono peggiorate e sono morte nelle ore o nei giorni successivi al parto (56). Pertanto, sono necessari studi più ampi che affrontano la mortalità da COVID-19 durante la gravidanza rispetto al postpartum precoce come endpoint primario per determinare se l’ambiente ormonale del terzo trimestre è protettivo.
Immunomodulazione mediante terapie ormonali nelle donne
Il trattamento delle donne in postmenopausa con terapia ormonale menopausale (MHT) e l’uso di contraccettivi orali da parte delle donne in età riproduttiva sono accompagnati da cambiamenti fisiologici concomitanti associati ad un aumento delle concentrazioni di estrogeni e progestinici.
Pertanto, gli effetti di questi 2 ormoni non possono essere separati. La maggior parte degli studi che valutano l’effetto dell’MHT utilizzando E2 da solo o in combinazione con progestinici hanno mostrato che l’MHT inibisce la produzione di citochine proinfiammatorie (p. Es., TNF-α, IL-1β e IL-6) da parte delle cellule mononucleate del sangue periferico ex vivo o in vivo nel siero delle donne trattate con MHT (57-60).
Inoltre, l’E2 transdermico ha attenuato le risposte delle citochine proinfiammatorie a una provocazione infiammatoria (59). L’effetto antinfiammatorio della terapia E2 nelle donne in menopausa per quanto riguarda l’infiammazione sistemica di basso grado sembrava essere osservato principalmente dopo somministrazione transdermica piuttosto che orale di E2 e non è stato riprodotto da estrogeni equini coniugati. Il bazedoxifene appartiene a una nuova generazione di SERM utilizzati in combinazione con estrogeni nella terapia ormonale della menopausa orale.
Nelle femmine di topo obese, il trattamento con bazedoxifene riduce l’IL-6 e molteplici marker di infiammazione sistemica (61).
Tuttavia, questo effetto non è stato osservato in uno studio pilota randomizzato di 8 settimane di trattamento con estrogeni orali e bazedoxifene in donne obese in postmenopausa (62). Probabilmente, l’assenza di effetti benefici degli estrogeni somministrati per via orale sull’infiammazione sistemica è correlata al metabolismo epatico di primo passaggio dopo somministrazione di estrogeni per via orale, che aumenta la produzione di proteina C-reattiva e i marker di infiammazione (63).
La terapia estrogenica transdermica e orale con o senza progestinico aumenta il numero e l’attività dei linfociti B CD19 + (64). Di conseguenza, gli effetti stimolanti della terapia menopausale con estrogeni e progestinici sui linfociti B favoriscono la progressione del lupus eritematoso sistemico nelle donne in postmenopausa (33).
I recettori classici ERα, ERβ e P4 sono presenti sia nei pool extranucleari che nucleari nella maggior parte delle cellule (65). Fino a che punto ogni pool cellulare del recettore ha effetti collaborativi o unici sulla funzione immunitaria non è stato determinato e sarà di interesse per la progettazione di studi futuri che valutino l’effetto dei ligandi del recettore degli steroidi sessuali nel modulare le funzioni immunitarie.
Riproporre estrogeni e progesterone per mitigare la malattia da coronavirus – mortalità 2019?
Alte concentrazioni fisiologiche di E2 e P4 possono essere sinergizzate per mitigare la produzione di cellule immunitarie innate di citochine proinfiammatorie, promuovere le risposte antinfiammatorie delle cellule T e la tolleranza immunitaria e stimolare la produzione di anticorpi da parte delle cellule B (Fig.1).
Negli individui con COVID-19 confermato, la terapia ormonale acuta con E2 e P4 potrebbe mitigare la tempesta di citochine aumentando la produzione di anticorpi.
Pandemie come SARS-CoV-2 forniscono poco tempo per lo sviluppo di farmaci. Il riutilizzo di farmaci esistenti e approvati che sono già stati testati sugli esseri umani – e per i quali sono disponibili informazioni dettagliate sulla loro farmacologia, formulazione, dose e potenziale tossicità – fornisce un approccio rapido e sicuro per l’uso off-label di terapie potenzialmente salvavita.
Come discusso in precedenza, ci si aspetta che il trattamento acuto con E2 e P4 attenui le risposte immunitarie infiammatorie innate e allo stesso tempo stimoli le risposte delle cellule B e la produzione di anticorpi (33, 51) senza effetti collaterali evidenti.
Un vantaggio fondamentale di estrogeni, SERM e composti progestinici è la profonda conoscenza della loro efficacia clinica e tossicità accumulata in decenni di studi clinici e di base. La terapia ormonale è utilizzata da milioni di donne per la contraccezione e la prevenzione dei sintomi della menopausa.
È ampiamente disponibile negli ospedali, poco costoso, realizzabile in scala e può essere prescritto immediatamente. Mentre questa recensione è in fase di scrittura, 2 studi clinici stanno testando E2 (identificatore ClinicalTrials.gov NCT04359329) o P4 (identificatore ClinicalTrials.gov NCT04365127) individualmente in pazienti COVID-19.
Vale la pena considerare il potenziale beneficio della terapia ormonale da sola o in combinazione con farmaci antivirali o blocco di IL-6 come una modulazione immunitaria in studi clinici off-label monocentrici. In un’epidemia come questa, e mentre siamo in attesa dello sviluppo di un vaccino sicuro ed efficiente, l’indagine sistematica dei farmaci clinicamente approvati è una priorità per determinare quali composti possono mitigare la malattia e per investire risorse per iniziare la produzione su vasta scala.
Riferimenti
References
1. Karlberg J, Chong DS, Lai WY. Do men have a higher case fatality rate of severe acute respiratory syndrome than women do? Am J Epidemiol. 2004;159(3):229-231. [PMC free article] [PubMed] [Google Scholar]
2. Alghamdi IG, Hussain II, Almalki SS, Alghamdi MS, Alghamdi MM, El-Sheemy MA. The pattern of middle east respiratory syndrome coronavirus in Saudi Arabia: a descriptive epidemiological analysis of data from the Saudi Ministry of Health. Int J Gen Med. 2014;7:417-423. [PMC free article] [PubMed] [Google Scholar]
3. Mauvais-Jarvis F, Bairey Merz CN, Barnes PJ, et al. . Sex and gender: modifiers of health, disease and medicine. Lancet. 2020;396. [PMC free article] [PubMed] [Google Scholar]
4. Guan WJ, Ni ZY, Hu Y, et al. . Clinical characteristics of coronavirus disease 2019 in China. N Engl J Med. 2020;382:1708-1720. [PMC free article] [PubMed] [Google Scholar]
5. Onder G, Rezza G, Brusaferro S. Case-fatality rate and characteristics of patients dying in relation to COVID-19 in Italy. JAMA. 2020;323(18):1775-1776. [PubMed] [Google Scholar]
6. COVID-19 National Emergency Response Center, Epidemiology and Case Management Team, Korea Centers for Disease Control and Prevention. Coronavirus disease-19: the first 7755 cases in the Republic of Korea. Osong Public Health Res Perspect. 2020;11(2):85-90. [PMC free article] [PubMed] [Google Scholar]
7. Richardson S, Hirsch JS, Narasimhan M, Crawford JM, McGinn T, Davidson KW, and the Northwell COVID-19 Research Consortium Presenting characteristics, comorbidities, and outcomes among 5700 patients hospitalized with COVID-19 in the New York City Area. JAMA. 2020;323(20):2052-2059. [PMC free article] [PubMed] [Google Scholar]
8. Klein S, Dhakal S, Ursin R, Deshpante S, Sandberg K, Mauvais-Jarvis F. Biological sex impacts COVID-19 outcomes. PLoS Pathog. 2020;16(6):e1008570. [PMC free article] [PubMed] [Google Scholar]
9. Grasselli G, Zangrillo A, Zanella A, et al. ; for the COVID-19 Lombardy ICU Network Baseline characteristics and outcomes of 1591 patients infected with SARS-CoV-2 admitted to ICUs of the Lombardy Region, Italy. JAMA. 2020;323(16):1574-1581. [PMC free article] [PubMed] [Google Scholar]
10. Docherty AB, Harrison EM, Green CA, et al. . Features of 16 749 hospitalised UK patients with COVID-19 using the ISARIC WHO clinical characterisation protocol. [Published online ahead of print April 28, 2020.] medRxiv. Doi:10.1101/2020.04.23.20076042. [PMC free article] [PubMed] [Google Scholar]
11. Scully E, Haverfield J, Ursin R, Tannenbaum C, Klein SL. Sex is a variable in immune responses and COVID-19 outcomes. Nat Rev Immunol. 2020;20:442-447. [PMC free article] [PubMed] [Google Scholar]
12. Marina S, Piemonti L. Gender and age effects on the rates of infection and deaths in individuals with confirmed SARS-CoV-2 infection in six European countries. SSRN website. https://ssrn.com/abstract=3576790 or http://dx.doi.org/10.2139/ssrn.3576790. Posted April 28, 2020. Accessed June 20, 2020.
13. Chousterman BG, Swirski FK, Weber GF. Cytokine storm and sepsis disease pathogenesis. Semin Immunopathol. 2017;39(5):517-528. [PubMed] [Google Scholar]
14. Ye Q, Wang B, Mao J. The pathogenesis and treatment of the ‘Cytokine Storm’ in COVID-19. J Infect. 2020;80(6):607-613. [PMC free article] [PubMed] [Google Scholar]
15. Chen G, Wu D, Guo W, et al. . Clinical and immunological features of severe and moderate coronavirus disease 2019. J Clin Invest. 2020;130(5):2620-2629. [PMC free article] [PubMed] [Google Scholar]
16. Mehta P, McAuley DF, Brown M, Sanchez E, Tattersall RS, Manson JJ; HLH Across Speciality Collaboration, UK COVID-19: consider cytokine storm syndromes and immunosuppression. Lancet. 2020;395(10229):1033-1034. [PMC free article] [PubMed] [Google Scholar]
17. Tanaka T, Narazaki M, Kishimoto T. Immunotherapeutic implications of IL-6 blockade for cytokine storm. Immunotherapy. 2016;8(8):959-970. [PubMed] [Google Scholar]
18. McGonagle D, Sharif K, O’Regan A, Bridgewood C. The role of cytokines including interleukin-6 in COVID-19 induced pneumonia and macrophage activation syndrome-like disease. Autoimmun Rev. 2020;19(6):102537. [PMC free article] [PubMed] [Google Scholar]
19. Luo P, Liu Y, Qiu L, Liu X, Liu D, Li J. Tocilizumab treatment in COVID-19: a single center experience. J Med Virol. 2020;92(7):814-818. [PMC free article] [PubMed] [Google Scholar]
20. Leng Z, Zhu R, Hou W. Transplantation of ACE2– mesenchymal stem cells improves the outcome of patients with COVID-19 pneumonia. Aging Dis. 2020;11(2):216-228. [PMC free article] [PubMed] [Google Scholar]
21. Butterworth M, McClellan B, Allansmith M. Influence of sex in immunoglobulin levels. Nature. 1967;214(5094):1224-1225. [PubMed] [Google Scholar]
22. Klein SL, Jedlicka A, Pekosz A. The Xs and Y of immune responses to viral vaccines. Lancet Infect Dis. 2010;10(5):338-349. [PMC free article] [PubMed] [Google Scholar]
23. Amadori A, Zamarchi R, De Silvestro G, et al. . Genetic control of the CD4/CD8 T-cell ratio in humans. Nat Med. 1995;1(12):1279-1283. [PubMed] [Google Scholar]
24. Klein SL, Flanagan KL. Sex differences in immune responses. Nat Rev Immunol. 2016;16(10):626-638. [PubMed] [Google Scholar]
25. Migeon BR. Females are Mosaics: X Inactivation and Sex Differences in Disease. 2nd ed. New York: Oxford University Press; November 12, 2013. [Google Scholar]
26. Carrel L, Brown CJ. When the Lyon(ized chromosome) roars: ongoing expression from an inactive X chromosome. Philos Trans R Soc Lond B Biol Sci. 2017;372(1733):20160355. [PMC free article] [PubMed] [Google Scholar]
27. Tukiainen T, Villani AC, Yen A, et al. ; GTEx Consortium; Laboratory, Data Analysis &Coordinating Center (LDACC)—Analysis Working Group; Statistical Methods groups—Analysis Working Group; Enhancing GTEx (eGTEx) groups; NIH Common Fund; NIH/NCI; NIH/NHGRI; NIH/NIMH; NIH/NIDA; Biospecimen Collection Source Site—NDRI; Biospecimen Collection Source Site—RPCI; Biospecimen Core Resource—VARI; Brain Bank Repository—University of Miami Brain Endowment Bank; Leidos Biomedical—Project Management; ELSI Study; Genome Browser Data Integration &Visualization—EBI; Genome Browser Data Integration &Visualization—UCSC Genomics Institute, University of California Santa Cruz Landscape of X chromosome inactivation across human tissues. Nature. 2017;550(7675):244-248. [PMC free article] [PubMed] [Google Scholar]
28. Souyris M, Cenac C, Azar P, et al. . TLR7 escapes X chromosome inactivation in immune cells. Sci Immunol. 2018;3(19):eaap8855. [PubMed] [Google Scholar]
29. Fink AL, Engle K, Ursin RL, Tang WY, Klein SL. Biological sex affects vaccine efficacy and protection against influenza in mice. Proc Natl Acad Sci U S A. 2018;115(49):12477-12482. [PMC free article] [PubMed] [Google Scholar]
30. Case LK, Wall EH, Dragon JA, et al. . The Y chromosome as a regulatory element shaping immune cell transcriptomes and susceptibility to autoimmune disease. Genome Res. 2013;23(9):1474-1485. [PMC free article] [PubMed] [Google Scholar]
31. Krementsov DN, Case LK, Dienz O, et al. . Genetic variation in chromosome Y regulates susceptibility to influenza A virus infection. Proc Natl Acad Sci U S A. 2017;114(13):3491-3496. [PMC free article] [PubMed] [Google Scholar]
32. Phiel KL, Henderson RA, Adelman SJ, Elloso MM. Differential estrogen receptor gene expression in human peripheral blood mononuclear cell populations. Immunol Lett. 2005;97(1):107-113. [PubMed] [Google Scholar]
33. Straub RH. The complex role of estrogens in inflammation. Endocr Rev. 2007;28(5):521-574. [PubMed] [Google Scholar]
34. Manolagas SC. Role of cytokines in bone resorption. Bone. 1995;17(2 Suppl 1):S63-S67. [PubMed] [Google Scholar]
35. Shi L, Feng Y, Lin H, Ma R, Cai X. Role of estrogen in hepatocellular carcinoma: is inflammation the key? J Transl Med. 2014;12:93. [PMC free article] [PubMed] [Google Scholar]
36. Di Martino V, Lebray P, Myers RP, et al. . Progression of liver fibrosis in women infected with hepatitis C: long-term benefit of estrogen exposure. Hepatology. 2004;40(6):1426-1433. [PubMed] [Google Scholar]
37. Naugler WE, Sakurai T, Kim S, et al. . Gender disparity in liver cancer due to sex differences in MyD88-dependent IL-6 production. Science. 2007;317(5834):121-124. [PubMed] [Google Scholar]
38. Speyer CL, Rancilio NJ, McClintock SD, et al. . Regulatory effects of estrogen on acute lung inflammation in mice. Am J Physiol Cell Physiol. 2005;288(4):C881-C890. [PubMed] [Google Scholar]
39. Robinson DP, Lorenzo ME, Jian W, Klein SL. Elevated 17β-estradiol protects females from influenza A virus pathogenesis by suppressing inflammatory responses. PloS Pathog. 2011;7(7):e1002149. [PMC free article] [PubMed] [Google Scholar]
40. Robinson DP, Hall OJ, Nilles TL, Bream JH, Klein SL. 17β-estradiol protects females against influenza by recruiting neutrophils and increasing virus-specific CD8 T cell responses in the lungs. J Virol. 2014;88(9):4711-4720. [PMC free article] [PubMed] [Google Scholar]
41. Vermillion MS, Ursin RL, Attreed SE, Klein SL. Estriol reduces pulmonary immune cell recruitment and inflammation to protect female mice from severe influenza. Endocrinology. 2018;159(9):3306-3320. [PMC free article] [PubMed] [Google Scholar]
42. Peretz J, Pekosz A, Lane AP, Klein SL. Estrogenic compounds reduce influenza A virus replication in primary human nasal epithelial cells derived from female, but not male, donors. Am J Physiol Lung Cell Mol Physiol. 2016;310(5):L415-L425. [PMC free article] [PubMed] [Google Scholar]
43. Channappanavar R, Fett C, Mack M, Ten Eyck PP, Meyerholz DK, Perlman S. Sex-based differences in susceptibility to severe acute respiratory syndrome coronavirus infection. J Immunol. 2017;198(10):4046-4053. [PMC free article] [PubMed] [Google Scholar]
44. Dyall J, Coleman CM, Hart BJ, et al. . Repurposing of clinically developed drugs for treatment of Middle East respiratory syndrome coronavirus infection. Antimicrob Agents Chemother. 2014;58(8):4885-4893. [PMC free article] [PubMed] [Google Scholar]
45. Johansen LM, Brannan JM, Delos SE, et al. . FDA-approved selective estrogen receptor modulators inhibit Ebola virus infection. Sci Transl Med. 2013;5(190):190ra79. [PMC free article] [PubMed] [Google Scholar]
46. Piccinni MP, Giudizi MG, Biagiotti R, et al. . Progesterone favors the development of human T helper cells producing Th2-type cytokines and promotes both IL-4 production and membrane CD30 expression in established Th1 cell clones. J Immunol. 1995;155(1):128-133. [PubMed] [Google Scholar]
47. Szekeres-Bartho J, Wegmann TG. A progesterone-dependent immunomodulatory protein alters the Th1/Th2 balance. J Reprod Immunol. 1996;31(1-2):81-95. [PubMed] [Google Scholar]
48. Hall OJ, Limjunyawong N, Vermillion MS, et al. . Progesterone-based therapy protects against influenza by promoting lung repair and recovery in females. PloS Pathog. 2016;12(9):e1005840. [PMC free article] [PubMed] [Google Scholar]
49. Gordon DE, Jang GM, Bouhaddou M, et al. . A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature. 2020;583(7816):459-468. [PMC free article] [PubMed] [Google Scholar]
50. Robinson DP, Klein SL. Pregnancy and pregnancy-associated hormones alter immune responses and disease pathogenesis. Horm Behav. 2012;62(3):263-271. [PMC free article] [PubMed] [Google Scholar]
51. Doria A, Iaccarino L, Arienti S, et al. . Th2 immune deviation induced by pregnancy: the two faces of autoimmune rheumatic diseases. Reprod Toxicol. 2006;22(2):234-241. [PubMed] [Google Scholar]
52. Szekeres-Bartho J, Faust Z, Varga P, Szereday L, Kelemen K. The immunological pregnancy protective effect of progesterone is manifested via controlling cytokine production. Am J Reprod Immunol. 1996;35(4):348-351. [PubMed] [Google Scholar]
53. Pazos M, Sperling RS, Moran TM, Kraus TA. The influence of pregnancy on systemic immunity. Immunol Res. 2012;54(1-3):254-261. [PMC free article] [PubMed] [Google Scholar]
54. Qiancheng X, Jian S, Lingling P, et al. . Coronavirus disease 2019 in pregnancy. Int J Infect Dis. 2020;95:376-383. [PMC free article] [PubMed] [Google Scholar]
55. Chen L, Li Q, Zheng D, et al. . Clinical characteristics of pregnant women with Covid-19 in Wuhan, China. N Engl J Med. 2020;382(25):e100. [PMC free article] [PubMed] [Google Scholar]
56. Hantoushzadeh S, Shamshirsaz AA, Aleyasin A, et al. . Maternal death due to COVID-19 Disease. Am J Obstet Gynecol. 2020;223(1):109.e1-109.e16. [PMC free article] [PubMed] [Google Scholar]
57. Bernard-Poenaru O, Roux C, Blanqué R, Gardner C, de Vemejoul MC, Cohen-Solal ME. Bone-resorbing cytokines from peripheral blood mononuclear cells after hormone replacement therapy: a longitudinal study. Osteoporos Int. 2001;12(9):769-776. [PubMed] [Google Scholar]
58. Rachoń D, Myśliwska J, Suchecka-Rachoń K, Wieckiewicz J, Myśliwski A. Effects of oestrogen deprivation on interleukin-6 production by peripheral blood mononuclear cells of postmenopausal women. J Endocrinol. 2002;172(2):387-395. [PubMed] [Google Scholar]
59. Puder JJ, Freda PU, Goland RS, Wardlaw SL. Estrogen modulates the hypothalamic-pituitary-adrenal and inflammatory cytokine responses to endotoxin in women1. J Clin Endocrinol Metab. 2001;86(6):2403-2408. [PubMed] [Google Scholar]
60. Aune B, Øian P, Omsjø I, Østerud B. Hormone replacement therapy reduces the reactivity of monocytes and platelets in whole blood—a beneficial effect on atherogenesis and thrombus formation? Am J Obstet Gynecol. 1995;173(6):1816-1820. [PubMed] [Google Scholar]
61. Kim JH, Meyers MS, Khuder SS, et al. . Tissue-selective estrogen complexes with bazedoxifene prevent metabolic dysfunction in female mice. Mol Metab. 2014;3(2):177-190. [PMC free article] [PubMed] [Google Scholar]
62. Lovre D, Peacock E, Katalenich B, et al. . Conjugated estrogens and bazedoxifene improve β cell function in obese menopausal women. J Endocr Soc. 2019;3(8):1583-1594. [PMC free article] [PubMed] [Google Scholar]
63. Mauvais-Jarvis F, Manson JE, Stevenson JC, Fonseca VA. Menopausal hormone therapy and type 2 diabetes prevention: evidence, mechanisms, and clinical implications. Endocr Rev. 2017;38(3):173-188. [PMC free article] [PubMed] [Google Scholar]
64. Porter VR, Greendale GA, Schocken M, Zhu X, Effros RB. Immune effects of hormone replacement therapy in post-menopausal women. Exp Gerontol. 2001;36(2):311-326. [PubMed] [Google Scholar]
65. Levin ER, Hammes SR. Nuclear receptors outside the nucleus: extranuclear signalling by steroid receptors. Nat Rev Mol Cell Biol. 2016;17(12):783-797. [PMC free article] [PubMed] [Google Scholar]
*-*-*-
26a.Shastri A, Wheat J, Agrawal S, Chaterjee N, Pradhan K, Goldfinger M, et al. Delayed clearance of sars-cov2 in male compared to female patients: high ace2 expression in testes suggests possible existence of gender-specific viral reservoirs. MedRxiv. 2020; 2020.2004.2016.20060566.
51a.Klein SL, Flanagan KL. Sex differences in immune responses. Nat Rev Immunol. 2016;16:626–38.CAS PubMed Article Google Scholar
52a.Schurz H, Salie M, Tromp G, Hoal EG, Kinnear CJ, Moller M. The x chromosome and sex-specific effects in infectious disease susceptibility. Hum Genomics. 2019;13.CASPubMedArticleGoogle Scholar
55a. Olsen NJ, Kovacs WJ. Gonadal steroids and immunity. J Endocrin Rev. 1996;17:369–84.
63a.Channappanavar R, Fett C, Mack M, Ten Eyck PP, Meyerholz DK, Perlman S. Sex-based differences in susceptibility to severe acute respiratory syndrome coronavirus infection. J Immunol. 2017;198:4046–53.CAS PubMed PubMed Central Article Google Scholar
72a.Ghosh S, Klein RS. Sex drives dimorphic immune responses to viral infections. J Immunol. 2017;198:1782–90.CAS PubMed PubMed Central Article Google Scholar
73a.Phoenix CH, Goy RW, Gerall AA, Young WC. Organizing action of prenatally administered testosterone propionate on the tissues mediating mating behavior in the female guinea pig. Endocrinology. 1959;65:369–82.CAS PubMed Article Google Scholar
74a.Koopman P. Sex determination: a tale of two sox genes. Trends Genet. 2005;21:367–70.CAS PubMed Article Google Scholar
75a.Arnold AP, Burgoyne PS. Are xx and xy brain cells intrinsically different? Trends Endocrinol Metab. 2004;15:6–11.CAS PubMed Article Google Scholar
76a.Nilsson S, Makela S, Treuter E, Tujague M, Thomsen J, Andersson G, et al. Mechanisms of estrogen action. Physiol Rev. 2001;81:1535–65.CAS PubMed Article Google Scholar
77a.Kovats S. Estrogen receptors regulate innate immune cells and signaling pathways. Cell Immunol. 2015;294:63–9.CAS PubMed PubMed Central Article Google Scholar
78a.Phiel KL, Henderson RA, Adelman SJ, Elloso MM. Differential estrogen receptor gene expression in human peripheral blood mononuclear cell populations. Immunol Lett. 2005;97:107–13.CAS PubMed Article Google Scholar
79a.Pierdominici M, Maselli A, Colasanti T, Giammarioli AM, Delunardo F, Vacirca D, et al. Estrogen receptor profiles in human peripheral blood lymphocytes. Immunol Lett. 2010;132:79–85.CAS PubMed Article Google Scholar
80a.Nakada D, Oguro H, Levi BP, Ryan N, Kitano A, Saitoh Y, et al. Oestrogen increases haematopoietic stem-cell self-renewal in females and during pregnancy. Nature. 2014;505:555–8.CAS PubMed PubMed Central Article Google Scholar
81a.Kissick HT, Sanda MG, Dunn LK, Pellegrini KL, On ST, Noel JK, et al. Androgens alter t-cell immunity by inhibiting t-helper 1 differentiation. 2014;111:9887–92.
82a.Muehlenbein MP, Cogswell FB, James MA, Koterski J, Ludwig GV. Testosterone correlates with venezuelan equine encephalitis virus infection in macaques. Virol J. 2006;3:19.PubMed PubMed Central Article CAS Google Scholar
83a.Furman D, Hejblum BP, Simon N, Jojic V, Dekker CL, Thiébaut R, et al. Systems analysis of sex differences reveals an immunosuppressive role for testosterone in the response to influenza vaccination. Proc Natl Acad Sci U S A. 2014;111:869–74.CAS PubMed Article Google Scholar
84a.Drake CG, Doody AD, Mihalyo MA, Huang CT, Kelleher E, Ravi S, et al. Androgen ablation mitigates tolerance to a prostate/prostate cancer-restricted antigen. Cancer Cell. 2005;7:239–49.CAS PubMed PubMed Central Article Google Scholar
85a.Viselli SM, Stanziale S, Shults K, Kovacs WJ, Olsen NJ. Castration alters peripheral immune function in normal male mice. Immunology. 1995;84:337–42.CAS PubMed PubMed Central Google Scholar
86a.Montopoli M, Zumerle S, Vettor R, Rugge M, Zorzi M, Catapano CV, et al. Androgen-deprivation therapies for prostate cancer and risk of infection by sars-cov-2: a population-based study (n = 4532). Ann Oncol. 2020.
87a.Lucas JM, Heinlein C, Kim T, Hernandez SA, Malik MS, True LD, et al. The androgen-regulated protease tmprss2 activates a proteolytic cascade involving components of the tumor microenvironment and promotes prostate cancer metastasis. Cancer Discov. 2014;4:1310–25.CAS PubMed PubMed Central Article Google Scholar
88a.Sharifi N, Ryan CJ. Androgen hazards with covid-19. Endocr Relat Cancer. 2020;27:E1–3.PubMed Article Google Scholar
89a.Baratchian M, McManus J, Berk M, Nakamura F, Erzurum S, Mukhopadhyay S, et al. 2020. No evidence that androgen regulation of pulmonary tmprss2 explains sex-discordant covid-19 outcomes.2020.2004.2021.051201.
90a.Stopsack KH, Mucci LA, Antonarakis ES, Nelson PS, Kantoff PW. Tmprss2 and covid-19: serendipity or opportunity for intervention? Cancer Discov. 2020;10:779–82.PubMed Article Google Scholar
91a.Peretz J, Pekosz A, Lane AP, Klein SL. Estrogenic compounds reduce influenza A virus replication in primary human nasal epithelial cells derived from female, but not male, donors. Am J Phys Lung Cell Mol Phys. 2016;310:L415–25.Google Scholar
92a.Zhao ST, Zhu WW, Xue SP, Han DS. Testicular defense systems: immune privilege and innate immunity. Cell Mol Immunol. 2014;11:428–37.PubMed PubMed Central Article Google Scholar
93a.Dejucq N, Jegou B. Viruses in the mammalian male genital tract and their effects on the reproductive system. Microbiol Mol Biol R. 2001;65:208–31.CAS Article Google Scholar
94a.Salam AP, Horby PW. The breadth of viruses in human semen. Emerg Infect Dis. 2017;23:1922–4.PubMed PubMed Central Article Google Scholar
95a.Li D, Jin M, Bao P, Zhao W, Zhang S. Clinical characteristics and results of semen tests among men with coronavirus disease 2019. JAMA Netw Open. 2020;3:e208292.PubMed PubMed Central Article Google Scholar
96a.Fagerberg L, Hallstrom BM, Oksvold P, Kampf C, Djureinovic D, Odeberg J, et al. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol Cell Proteomics. 2014;13:397–406.CAS PubMed Article Google Scholar
97a.Ma L, Xie W, Li D, Shi L, Mao Y, Xiong Y, et al. Effect of sars-cov-2 infection upon male gonadal function: a single center-based study. MedRxiv. 2020; 2020.2003.2021.20037267.



{kind=link}