










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



In a paper published today in the journal Cell, scientists describe unexpected, undesirable outcomes after editing genes in human embryos with CRISPR, a genomic editing system.
The study, the most detailed analysis to date of CRISPR in human embryos, shows that applying CRISPR to repair a blindness-causing gene early in the development of a human embryo often eliminates an entire chromosome or a large section of it.
“Our study shows that CRISPR/Cas9 is not yet ready for clinical use to correct mutations at this stage of human development,” says the study’s senior author Dieter Egli, assistant professor of developmental cell biology in the Department of Pediatrics at Columbia University Vagelos College of Physicians and Surgeons.
CRISPR in human embryos
The genome editing system called CRISPR/Cas9 has revolutionized molecular biology in recent years, and its discoverers were honored with this year’s Nobel Prize in chemistry. The system allows scientists to make precise changes in the genomes of cultured cells, living tissues, and animal embryos.
The first use of CRISPR in human embryos was reported in 2015. Just a few years later in 2018, He Jiankui claimed to have performed the procedure in a pair of twins, eliciting a firestorm of condemnation from scientists and government officials worldwide.
CRISPR can lead to chromosomal abnormalities
In the new paper, Egli and his colleagues tested CRISPR/Cas9 genome editing’s effects on early-stage human embryos carrying a mutation in a gene called EYS (eyes shut homolog), which causes hereditary blindness.
“We know from previous studies in differentiated human cells and in mice that a break in the DNA results in mostly two outcomes: precise repair or small local changes.
At the EYS gene, these changes can yield a functional gene, though it is not a perfect repair,” says Michael Zuccaro, a Ph.D. candidate at Columbia University Vagelos College of Physicians and Surgeons and co-lead author of the paper.
When Zuccaro, Egli and their colleagues looked at the entire genomes of the modified embryos in this study, they identified another outcome.
“We learned that in human embryonic cells, a single break in the DNA can result a third outcome, the loss of an entire chromosome or sometimes a large segment of that chromosome, and this loss of the chromosome is very frequent,” says Zuccaro.
In 2017, a study reported the successful correction of a heart disease-causing mutation in normal human embryos using CRISPR.
Comprehensive data from the new study offer a different interpretation of these results: instead of being corrected, the chromosome carrying the mutation may have been lost altogether.
“If our results had been known two years ago, I doubt that anyone would have gone ahead with an attempt to use CRISPR to edit a gene in a human embryo in the clinic,” Egli says.
“Our hope is that these cautionary findings should discourage premature clinical application of this important technology, but can also guide responsible research to achieve its ultimate safe and effective use.”
The study was published Oct. 29 in Cell, in a paper titled, “Allele-specific chromosome removal after Cas9 cleavage in human embryos.”
Genome editing (also called gene editing) is a group of technologies that give scientists the ability to change an organism’s DNA. These technologies allow genetic material to be added, removed, or altered at particular locations in the genome. Several approaches to genome editing have been developed.
A recent one is known as CRISPR-Cas9, which is short for clustered regularly interspaced short palindromic repeats and CRISPR-associated protein 9. The CRISPR-Cas9 system has generated a lot of excitement in the scientific community because it is faster, cheaper, more accurate, and more efficient than other existing genome editing methods.
CRISPR-Cas9 was adapted from a naturally occurring genome editing system in bacteria. The bacteria capture snippets of DNA from invading viruses and use them to create DNA segments known as CRISPR arrays.
The CRISPR arrays allow the bacteria to “remember” the viruses (or closely related ones). If the viruses attack again, the bacteria produce RNA segments from the CRISPR arrays to target the viruses’ DNA. The bacteria then use Cas9 or a similar enzyme to cut the DNA apart, which disables the virus.
The CRISPR-Cas9 system works similarly in the lab. Researchers create a small piece of RNA with a short “guide” sequence that attaches (binds) to a specific target sequence of DNA in a genome. The RNA also binds to the Cas9 enzyme.
As in bacteria, the modified RNA is used to recognize the DNA sequence, and the Cas9 enzyme cuts the DNA at the targeted location. Although Cas9 is the enzyme that is used most often, other enzymes (for example Cpf1) can also be used.
Once the DNA is cut, researchers use the cell’s own DNA repair machinery to add or delete pieces of genetic material, or to make changes to the DNA by replacing an existing segment with a customized DNA sequence.
Genome editing is of great interest in the prevention and treatment of human diseases. Currently, most research on genome editing is done to understand diseases using cells and animal models. Scientists are still working to determine whether this approach is safe and effective for use in people.
It is being explored in research on a wide variety of diseases, including single-gene disorders such as cystic fibrosis, hemophilia, and sickle cell disease. It also holds promise for the treatment and prevention of more complex diseases, such as cancer, heart disease, mental illness, and human immunodeficiency virus (HIV) infection.
Ethical concerns arise when genome editing, using technologies such as CRISPR-Cas9, is used to alter human genomes. Most of the changes introduced with genome editing are limited to somatic cells, which are cells other than egg and sperm cells.
These changes affect only certain tissues and are not passed from one generation to the next. However, changes made to genes in egg or sperm cells (germline cells) or in the genes of an embryo could be passed to future generations.
Germline cell and embryo genome editing bring up a number of ethical challenges, including whether it would be permissible to use this technology to enhance normal human traits (such as height or intelligence). Based on concerns about ethics and safety, germline cell and embryo genome editing are currently illegal in many countries.
CRISPR/Cas9 Mediated Gene Editing
Pioneering Discoveries in CRISPR/Cas9 Technology
The bacterial CRISPR locus was first described by Francisco Mojica (23) and later identified as a key element in the adaptive immune system in prokaryotes (24). The locus consists of snippets of viral or plasmid DNA that previously infected the microbe (later termed “spacers”), which were found between an array of short palindromic repeat sequences.
Later, Alexander Bolotin discovered the Cas9 protein in Streptococcus thermophilus, which unlike other known Cas genes, Cas9 was a large gene that encoded for a single-effector protein with nuclease activity (25). They further noted a common sequence in the target DNA adjacent to the spacer, later known as the protospacer adjacent motif (PAM)—the sequence needed for Cas9 to recognize and bind its target DNA (25).
Later studies reported that spacers were transcribed to CRISPR RNAs (crRNAs) that guide the Cas proteins to the target site of DNA (26). Following studies discovered the trans-activating CRISPR RNA (tracrRNA), which forms a duplex with crRNA that together guide Cas9 to its target DNA (27).
The potential use of this system was simplified by introducing a synthetic combined crRNA and tracrRNA construct called a single-guide RNA (sgRNA) (28). This was followed by studies demonstrating successful genome editing by CRISPR/Cas9 in mammalian cells, thereby opening the possibility of implementing CRISPR/Cas9 in gene therapy (29) (Figure 1).

Hallmarks of CRISPR Gene Therapy. Timeline highlighting major events of traditional gene therapy, CRISPR development, and CRISPR gene therapy. The text in red denotes gene therapy events which have raised significant ethical concerns.
Mechanistic Overview of CRISPR/Cas9-Mediated Genome Editing
CRISPR/Cas9 is a simple two-component system used for effective targeted gene editing. The first component is the single-effector Cas9 protein, which contains the endonuclease domains RuvC and HNH.
RuvC cleaves the DNA strand non-complementary to the spacer sequence and HNH cleaves the complementary strand. Together, these domains generate double-stranded breaks (DSBs) in the target DNA.
The second component of effective targeted gene editing is a single guide RNA (sgRNA) carrying a scaffold sequence which enables its anchoring to Cas9 and a 20 base pair spacer sequence complementary to the target gene and adjacent to the PAM sequence.
This sgRNA guides the CRISPR/Cas9 complex to its intended genomic location. The editing system then relies on either of two endogenous DNA repair pathways: non-homologous end-joining (NHEJ) or homology-directed repair (HDR) (Figure 2).
NHEJ occurs much more frequently in most cell types and involves random insertion and deletion of base pairs, or indels, at the cut site.
This error-prone mechanism usually results in frameshift mutations, often creating a premature stop codon and/or a non-functional polypeptide. This pathway has been particularly useful in genetic knock-out experiments and functional genomic CRISPR screens, but it can also be useful in the clinic in the context where gene disruption provides a therapeutic opportunity.
The other pathway, which is especially appealing to exploit for clinical purposes, is the error-free HDR pathway. This pathway involves using the homologous region of the unedited DNA strand as a template to correct the damaged DNA, resulting in error-free repair.
Experimentally, this pathway can be exploited by providing an exogenous donor template with the CRISPR/Cas9 machinery to facilitate the desired edit into the genome (30).

CRISPR/Cas9 mediated gene editing. Cas9 in complex with the sgRNA targets the respective gene and creates DSBs near the PAM region. DNA damage repair proceeds either through the NHEJ pathway or HDR. In the NHEJ pathway, random insertions and deletions (indels) are introduced at the cut side and ligated resulting in error-prone repair. In the HDR pathway, the homologous chromosomal DNA serves as a template for the damaged DNA during repair, resulting in error-free repair.
sgRNA Template Construction for Cas9 Gene Editing
Along with Cas9 nuclease, CRISPR experiments require the introduction of an sgRNA containing an approximately 20-base sequence specific to the target DNA 5′ of a non-variable scaffold sequence. sgRNA can be delivered as RNA or by transforming with a plasmid with the sgRNA-coding sequence under a promoter. A number of strategies have been developed to quickly swap out the 20 base sequences allowing convenient sgRNA cloning using NEB products.
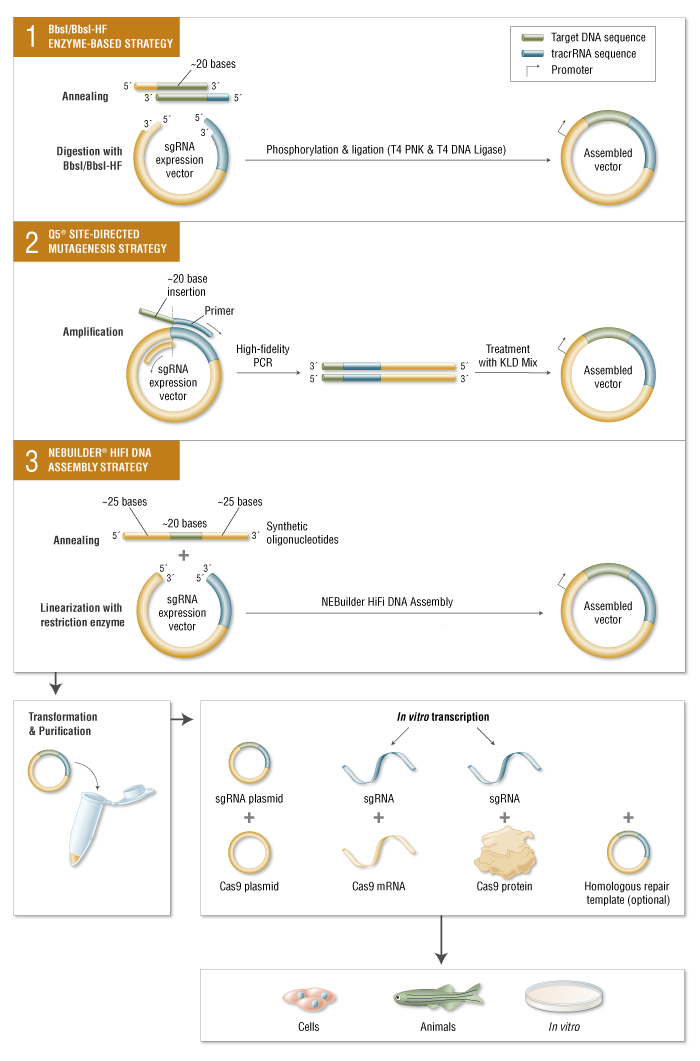
Limitations and Advancements of CRISPR/Cas9
Off-Target Effects
A major concern for implementing CRISPR/Cas9 for gene therapy is the relatively high frequency of off-target effects (OTEs), which have been observed at a frequency of ≥50% (31).
Current attempts at addressing this concern include engineered Cas9 variants that exhibit reduced OTE and optimizing guide designs.
One strategy that minimizes OTEs utilizes Cas9 nickase (Cas9n), a variant that induces single-stranded breaks (SSBs), in combination with an sgRNA pair targeting both strands of the DNA at the intended location to produce the DSB (32).
Researchers have also developed Cas9 variants that are specifically engineered to reduce OTEs while maintaining editing efficacy (Table 1).
SpCas9-HF1 is one of these high-fidelity variants that exploits the “excess-energy” model which proposes that there is an excess affinity between Cas9 and target DNA which may be enabling OTEs. By introducing mutations to 4 residues involved in direct hydrogen bonding between Cas9 and the phosphate backbone of the target DNA, SpCas9-HF1 has been shown to possess no detectable off-target activity in comparison to wildtype SpCas9 (35).
Other Cas9 variants that have been developed include evoCas9 and HiFiCas9, both of which contain altered amino acid residues in the Rec3 domain which is involved in nucleotide recognition. Desensitizing the Rec3 domain increases the dependence on specificity for the DNA:RNA heteroduplex to induce DSBs, thereby reducing OTEs while maintaining editing efficacy (38, 39).
One of the more recent developments is the Cas9_R63A/Q768A variant, in which the R63A mutation destabilizes R-loop formation in the presence of mismatches and Q768A mutation increases sensitivity to PAM-distal mismatches (49).
Despite the different strategies, the rational for generating many Cas9 variants with reduced OTEs has been to ultimately reduce general Cas9 and DNA interactions and give a stronger role for the DNA:RNA heteroduplex in facilitating the edits.
Table 1 – Cas9 variants.
| Bacterial origin | Cas9 variant | Advantage | Variant mutation | PAM | References |
|---|---|---|---|---|---|
| Streptococcus pyogenes | Cas9-D1135E | Improved PAM recognition | D1135E | NGG | (33) |
| Cas9-VQR | Altered PAM | D1135V/R1335Q/T1337R | NGAN or NGNG | ||
| Cas9-EQR | Altered PAM | D1135E/R1335Q/T1337R | NGAG | ||
| Cas9-VRER | Altered PAM | D1135V/G1218R/R1335E/T1337R | NGCG | ||
| Cas9-VRQR | Altered PAM | M495V/Y515N/K526E/R661Q | NGA | ||
| Cas9-QQR1 | Altered PAM | G1218R/N1286Q/I1331F/D1332K/R1333Q/R1335Q/T1337R | NAAG | (34) | |
| SpCas9-HF1 | Reduced OTE | N497A/R661A/Q695A/Q926A | NGG | (35) | |
| eSpCas9 | Reduced OTE | K846A/K1003A/R1060A | NGG | (36) | |
| HeFSpCas9 | Reduced OTE | N497A/R661A/Q695A/K846A/Q926A/K1003A/R1060A | NGG | (37) | |
| evoCas9 | Reduced OTE | M495V/Y515N/K526E/R661Q | NGG | (38) | |
| HiFiCas9 | Reduced OTE | R691A | NGG | (39) | |
| Cas9n/Cas9D10A | SSB instead of DSB, Reduced OTE | D10A | NGG | (40, 41) | |
| Dimeric dCas9-FokI | Reduced OTE | dCas9 fused to FokI endonuclease domain | NGG | (42) | |
| xCas9-3.7 | Broad PAM specificity | A262T/R324L/S409I/E480K/E543D/M694I/E1219V | NG, GAA or GAT | (43) | |
| SpCas9-NG | Minimal PAM | R1335V/L1111R/D1135V/G1218R/E1219F/A1322R/T1337R | NGN | (44) | |
| HypaCas9 | Reduced OTE | N692A/M694A/Q695A/H698A | NGG | (45) | |
| Sniper-Cas9 | Reduced OTE | F539S/M763I/K890N | NGG | (46) | |
| SpG Cas9 | Minimal PAM | D1135L/S1136W/G1218K/E1219Q/R1335Q/T1337R | NGN | (47) | |
| SpRY Cas9 | Minimal PAM | D1135L/S1136W/G1218K/E1219Q/R1335Q/T1337R/L1111R/A1322R/A61R/N1317R/R1333P | NRN>NYN | ||
| SpCas9-HF1 plus | Reduced OTE | N497A/Q695A/Q926A; amino acids 1005-1013 replaced with two glycine | NGG | (48) | |
| eSpCas9 plus | Reduced OTE | K848A/R1060A; amino acids 1005-1013 replaced with two glycine | NGG | ||
| Cas9_R63A/Q768A | Reduced OTE | R63A/Q768A | NGG | (49) | |
| Staphylococcus aureus | KKH SaCas9 | Relaxed PAM | E782K/N968K/R1015H | NNNRRT | (33) |
| SaCas9-HF | Reduced OTE | R245A/N413A/N419A/R654A | NNGRRT | (50) | |
| SaCas9-NR | Relaxed PAM | N986R | NNGRR | (51) | |
| SaCas9-RL | Relaxed PAM | N986R/R991L | NNGRR | ||
| Streptococcus canis | ScCas9 | Minimal PAM | N/A (wildtype) |
Optimizing guide designs can also reduce the frequency of OTEs (31). Many features in an sgRNA determine specificity including the seed sequence (a 10–12 bp region proximal to PAM on 3′ of spacer sequence) (29, 53), GC content (54, 55), and modifications such as 5′ truncation of the sgRNA (56).
Several platforms have also been designed to provide optimized guide sequences against target genes, including E-Crisp (31, 57), CRISPR-design, CasOFFinder, and others (31). However, many of these tools are designed based on computational algorithms with varying parameters or rely on phenotypic screens that may be specific to cell types and genomes, generating appreciable noise and lack of generalizability across different experimental setups (58, 59).
Recently, an additional guide design tool named sgDesigner was developed that addressed these limitations by employing a novel plasmid library in silico that contained both the sgRNA and the target site within the same construct.
This allowed collecting Cas9 editing efficiency data in an intrinsic manner and establish a new training dataset that avoids the biases introduced through other models. Furthermore, a comparative performance evaluation to predict sgRNA efficiency of sgDesigner with 3 other commonly used tools (Doench Rule Set 2, Sequence Scan for CRISPR and DeepCRISPR) revealed that sgDesigner outperformed all 3 designer tools in 6 independent datasets, suggesting that sgDesigner may be a more robust and generalizable platform (60).
Protospacer Adjacent Motif Requirement
An additional limitation of the technology is the requirement for a PAM near the target site. Cas9 from the bacteria Streptococcus pyogenes (SpCas9) is one of the most extensively used Cas9s with a relatively short canonical PAM recognition site: 5′NGG3′, where N is any nucleotide.
However, SpCas9 is relatively large and difficult to package into AAV vectors (61, 62), the most common delivery vehicle for gene therapy.
Staphylococcus aureus Cas9 (SaCas9) is a smaller ortholog that can be packaged more easily in AAV vectors but has a longer PAM sequence: 5′NNGRRT3′ or 5′NNGRR(N)3′, where R is any purine, which further narrows the window of therapeutic targeting sites.
Engineered SaCas9 variants have been made, such as KKH SaCas9, which recognizes a 5′NNNRRT3′ PAM, broadening the human targeting sites by 2- to 4-fold. OTEs, however, are observed with frequencies similar to wildtype SaCas9 and need to be considered in designing any therapeutic application (33).
Several other variants of SpCas9 have also been engineered for broadening the gene target window including SpCas9-NG, which recognizes a minimal NG PAM (44) and xCas9, which recognizes a broad range of PAM including NG, GAA, and GAT (43). A side by side comparison of both variants revealed that while SpCas9-NG had a broader PAM recognition, xCas9 had the lowest OTE in human cells (63).
Another Cas9 ortholog from the bacteria Streptococcus canis, ScCas9, has been recently characterized with a minimal PAM specificity of 5′NNG3′ and an 89.2% sequence homology to SpCas9 and comparable editing efficiency to SpCas9 in both bacterial and human cells (52).
The most recent development is a variant of SpCas9 named SpRY that has been engineered to be nearly PAMless, recognizing minimal NRN > NYN PAMs. This new variant can potentially edit any gene independent of a PAM requirement, and hence can be used therapeutically against several genetic diseases (47).
Alternatively, RNA-targeting Cas9 variants have been developed which also broaden the gene targeting spectrum by mitigating PAM requirement restrictions. S. pyogenese Cas9 (SpyCas9) can be manipulated to target RNA by providing a short oligonucleotide with a PAM sequence, known as a PAMmer (64, 65), and thus eliminates the need for a PAM site within the target region.
Other subsets of Cas enzymes have also been discovered that naturally target RNA independent of a PAM, such as Cas13d. Upon further engineering of this effector, CasRx was developed for efficient RNA-guided RNA targeting in human cells (66, 67). Although RNA-targeting CRISPR advances provide a therapeutic opportunity without the risk of DNA-damage toxicity, they exclude the potential for editing a permanent correction into the genome.
DNA-Damage Toxicity
CRISPR-induced DSBs often trigger apoptosis rather than the intended gene edit (68). Further safety concerns were revealed when using this tool in human pluripotent stem cells (hPSCs) which demonstrated that p53 activation in response to the toxic DSBs introduced by CRISPR often triggers subsequent apoptosis (69).
Thus, successful CRISPR edits are more likely to occur in p53 suppressed cells, resulting in a bias toward selection for oncogenic cell survival (70). In addition, large deletions spanning kilobases and complex rearrangements as unintended consequences of on-target activity have been reported in several instances (71, 72), highlighting a major safety issue for clinical applications of DSB-inducing CRISPR therapy.
Other variations of Cas9, such as catalytically inactive endonuclease dead Cas9 (dCas9) in which the nuclease domains are deactivated, may provide therapeutic utility while mitigating the risks of DSBs (73). dCas9 can transiently manipulate expression of specific genes without introducing DSBs through fusion of transcriptional activating or repressing domains or proteins to the DNA-binding effector (74).
Other variants such as Cas9n can also be considered, which induces SSBs rather than DSBs. Further modifications of these Cas9 variants has led to the development of base editors and prime editors, a key innovation for safe therapeutic application of CRISPR technology (see Precision Gene Editing With CRISPR section).
Immunotoxicity
In addition to technical limitations, CRISPR/Cas9, like traditional gene therapy, still raises concerns for immunogenic toxicity.
Charlesworth et al. showed that more than half of the human subjects in their study possessed preexisting anti-Cas9 antibodies against the most commonly used bacterial orthologs, SaCas9 and SpCas9 (75).
Furthermore, AAV vectors are also widely used to deliver CRISPR components for gene therapy. To this end, several Cas9 orthologs and AAV serotypes were tested based on sequence similarities and predicted binding strength to MHC class I and class II to screen for immune orthologs that can be used for safe repeated administration of AAV-CRISPR gene therapy.
Although no two AAV serotypes were found to completely circumvent immune recognition, the study verified 3 Cas9 orthologs [SpCas9, SaCas9, and Campylobacter jejuni Cas9 (CjCas9)] which showed robust editing efficiency and tolerated repeated administration due to reduced immunogenic toxicity in mice immunized against AAV and Cas9 (76).
A major caveat is pre-existing immunity in humans against 2 of these orthologs—SpCas9 and SaCas9, leaving CjCas9 as the only current option for this cohort of patients. However, this ortholog has not been well-studied in comparison to the other 2 orthologs and will need further investigation to provide evidence for its safety and efficacy for clinical use. Future studies may also identify other Cas9 immune-orthogonal orthologs for safe repeated gene therapy.
Precision Gene Editing With CRISPR
Precise-genome editing is essential for prospects of CRISPR gene therapy. Although HDR pathways can facilitate a desired edit, its low efficiency renders its utility for precise gene editing for clinical intervention highly limiting, with NHEJ as the default pathway human cells take for repair.
Enhancement of HDR efficiency has been achieved via suppression of the NHEJ pathway through chemical inhibition of key NHEJ modulating enzymes such as Ku (77), DNA ligase IV (78), and DNA-dependent protein kinases (DNA-PKcs) (79). Other strategies that improve HDR efficiency include using single-stranded oligodeoxynucleotide (ssODN) template, which contains the homology arms to facilitate recombination and the desired edit sequence, instead of double-stranded DNA (dsDNA).
Rationally designed ssODN templates with optimized length complementarity have been shown to increase HDR rates up to 60% in human cells for single nucleotide substitution (80). Furthermore, cell cycle stage plays a key role in determining the DNA-damage repair pathway a cell may take. HDR events are generally restricted to late S and G2 phases of the cell cycle, given the availability of the sister chromatid to serve as a template at these stages, whereas NHEJ predominates the G1, S, and G2 phases (81).
Pharmacological arrest at the S phase with aphidicolin increased HDR frequency in HEK293T with Cas9-guide ribonucleoprotein (RNP) delivery. Interestingly, cell arrest in the M phase using nocodazole with low concentrations of the Cas9-guide RNP complex yielded higher frequencies of HDR events in these cells, reaching a maximum frequency of up to 31% (82).
Although HDR is considered to be restricted to mitotic cells, a recent study revealed that the CRISPR/Cas9 editing can achieve HDR in mature postmitotic neurons. Nishiyama et al. successfully edited the CaMKIIα locus through HDR in postmitotic hippocampal neurons of adult mice in vitro using an AAV delivered Cas9, guide RNA, and donor template in the CaMKIIα locus, which achieved successful HDR-mediated edits in ~30% of infected cells.
Although HDR efficiency was dose-dependent on AAV delivered HDR machinery and off-target activity was not monitored, this study demonstrated CRISPR’s potential utility for translational neuroscience after further developments (83). To further exploit cell-cycle stage control as a means to favor templated repair, Cas9 conjugation to a part of Geminin, a substrate for G1 proteosome degradation, can limit Cas9 expression to S, G2, and M stages.
This strategy was shown to facilitate HDR events while mitigating undesired NHEJ edits in human immortalized and stem cells (84, 85). A more recent strategy combined a chemically modified Cas9 to the ssODN donor or a DNA adaptor that recruits the donor template, either of which improved HDR efficiency by localizing the donor template near the cleavage site (86).
Despite these advancements, HDR is still achieved at a relatively low efficiency in eukaryotic cells and use of relatively harmful agents in cells such as NHEJ chemical inhibitors may not be ideal in a clinical setting.
A recent advancement that allows precision gene editing independent of exploiting DNA damage response mechanisms is the CRISPR base editing (BE) system. In this system, a catalytically inactive dead Cas9 (dCas9) is conjugated to deaminase, which can catalyze the conversion of nucleotides via deamination.
For increased editing efficiency, Cas9 nickase (Cas9n) fused with deaminase is recently being more utilized over dCas9 for base editing, as the nicks created in a single strand of DNA induce higher editing efficiency. Currently, the two types of CRISPR base editors are cytidine base editors (CBEs) and adenosine base editors (ABEs).
CBEs catalyze the conversion of cytidine to uridine, which becomes thymine after DNA replication. ABEs catalyze the conversion of adenosine to inosine which becomes guanine after DNA replication (87). Base editors provide a means to edit single nucleotides without running the risk of causing DSB-induced toxicity.
However, base editors are limited to “A to T” and “C to G” conversions, narrowing its scope for single-base gene edition to only these bases. In addition, base editors still face some of the same challenges as the previously described CRISPR systems, including OTEs, more so with CBEs than ABEs (88, 89) and packaging constraints, namely in AAV vectors due to the large size of base editors (90).
Furthermore, the editing window for base editors are limited to a narrow range of a few bases upstream of the PAM (90). More recently, prime editing has been developed as a strategy to edit the genome to insert a desired stretch of edits without inducing DSBs (91).
This technology combines fusion of Cas9n with a reverse transcriptase and a prime editing guide RNA (pegRNA), which contains sgRNA sequence, primer binding site (PBS), and an RNA template encoding the desired edit on the 3′ end. Prime editors use Cas9n to nick one strand of the DNA and insert the desired edit via reverse transcription of the RNA template.
The synthesized edit is incorporated into the genome and the unedited strand is cleaved and repaired to match the inserted edit. With an optimized delivery system in place, base editors and primer editors can open the door for precision gene editing to correct and potentially cure a multitude of genetic diseases (Figure 3).

Precise Gene Editing. (A) CRISPR/Cas9-HDR. Cas9 induces a DSB. The exogenous ssODN carrying the sequence for the desired edit and homology arms is used as a template for HDR-mediated gene modification. (B) Base Editor. dCas9 or Cas9n is tethered to the catalytic portion of a deaminase. Cytosine deaminase catalyzes the formation of uridine from cytosine. DNA mismatch repair mechanisms or DNA replication yield an C:G to T:A single nucleotide base edit. Adenosine deaminase catalyzes the formation of inosine from adenosine. DNA mismatch repair mechanisms or DNA replication yield an A:T to G:C single nucleotide base edit. (C) Prime Editor. Cas9n is tethered to the catalytic portion of reverse transcriptase. The prime editor system uses pegRNA, which contains the guide spacer sequence, reverse transcriptase primer, which includes the sequence for the desired edit and a primer binding site (PBS). PBS hybridizes with the complementary region of the DNA and reverse transcriptase transcribes new DNA carrying the desired edit. After cleavage of the resultant 5′ flap and ligation, DNA repair mechanisms correct the unedited strand to match the edited strand. HDR, homology directed repair. DSB, double stranded break; SSB, single-stranded break; ssODN, single-stranded oligodeoxynucleotide.
Delivery of CRISPR Gene Therapy
The delivery modality of CRISPR tools greatly influences its safety and therapeutic efficacy. While traditional gene therapy utilizing viruses have been scrutinized for the risk of immunotoxicity and insertional oncogenesis, AAV vectors remain a key delivery vehicle for CRISPR gene therapy and continues to be extensively used for its high efficiency of delivery (92).
The CRISPR toolkit can be packaged as plasmid DNA encoding its components, including Cas9 and gRNA, or can be delivered as mRNA of Cas9 and gRNA. Nucleic acids of CRISPR can be packaged in AAV vectors for delivery or introduced to target cells via electroporation/nucleofection or microinjection, with the latter methods averting virus-associated risks.
However, microinjection can be technically challenging and is only suited for ex vivo delivery. Electroporation is also largely used for ex vivo but can be used in vivo for certain target tissues (93). However, high-voltage shock needed to permeabilize cell membranes via electroporation can be toxic and can lead to permanent permeabilization of treated cells (94).
In addition to viral toxicity, AAV delivery of CRISPR components yields longevity of expression, leading to greater incidence of OTEs. Alternatively, delivery of the Cas9 protein and gRNA as RNP complexes has reduced OTEs while maintained editing efficacy, owing to its transient expression and rapid clearance in the cell (95).
Once the delivery modality is selected, CRISPR/Cas9 edits can be facilitated either ex vivo where cells are genetically modified outside of the patient and reintroduced back, or in vivo with delivery of the CRISPR components directly into the patient where cells are edited (Figure 4). Both systems pose their own set of advantages and challenges.
Advantages for ex vivo delivery include greater safety since patients are not exposed to the gene altering tool, technical feasibility, and tighter quality control of the edited cells. However, challenges to this method include survival and retention of in vivo function of cells outside the patient after genetic manipulation and extensive culture in vitro. Also, an adequate supply of cells is needed for efficient re-engraftment. These conditions limit this method to certain cell types that can survive and be expanded in culture, such as hematopoietic stem and progenitor cells (HSPCs) (96) and T cells (97).

Delivery of CRISPR Therapy. Nucleic acids encoding CRISPR/Cas9 or its RNP complex can be packaged into delivery vehicles. Once packaged, edits can be facilitated either ex vivo or in vivo. Ex vivo editing involves extraction of target cells from the patient, cell culture, and expansion in vitro, delivery of the CRISPR components to yield the desired edits, selection, and expansion of edited cells, and finally reintroduction of therapeutic edited cells into the patient. In vivo editing can be systemically delivered via intravenous infusions to the patient, where the CRISPR cargo travels through the bloodstream via arteries leading to the target tissue, or locally delivered with injections directly to target tissue. Once delivered, the edits are facilitated in vivo to provide therapeutic benefit.
While ex vivo gene therapy has provided therapeutic benefit for hematological disorders and cancer immunotherapy, many tissue types are not suited for this method, severely limiting its therapeutic utility for other genetic diseases. in vivo manipulation is thus needed to expand CRISPR’s utility to treat a broader range of genetic diseases, such as Duchenne muscular dystrophy (DMD) (98) and hereditary tyrosinemia (99).
CRISPR components can be delivered in vivo systemically through intravenous injections or can be locally injected to specific tissues (Figure 4). With systemic delivery, the CRISPR components and its vehicle are introduced into the circulatory system where expression of the gene editing toolkit can be controlled to target specific organs via tissue-specific promoters (100). However, challenges of in vivo delivery include degradation by circulating proteases or nucleases, opsonization by opsonins, or clearance by the mononuclear phagocyte system (MPS).
Furthermore, the cargo must reach the target tissue and bypass the vascular endothelium, which are often tightly connected by cell-cell junctions (101), preventing accessibility to larger delivery vehicles (>1 nm diameter). Additionally, once the cargo has reached the target cells, they must be internalized, which is generally facilitated through endocytosis where they can be transported and degraded by lysosomal enzymes (102).
In addition, localization of the editing machinery near the point of injection can result in uneven distribution of the edited cell repertoire within the tissue, which may result in suboptimal therapeutic outcomes (102). While advancements are continuing to refine delivery techniques, the current systems have allowed CRISPR gene therapy to be used in the clinic.
CRISPR Editing in Human Embryos and Ethical Considerations
While somatic editing for CRISPR therapy has been permitted after careful consideration, human germline editing for therapeutic intent remains highly controversial. With somatic edition, any potential risk would be contained within the individual after informed consent to partake in the therapy. Embryonic editing not only removes autonomy in the decision-making process of the later born individuals, but also allows unforeseen and permanent side effects to pass down through generations.
This very power warrants proceeding with caution to prevent major setbacks as witnessed by traditional gene therapy. However, a controversial CRISPR trial in human embryos led by Jiankui He may have already breached the ethical standards set in place for such trials.
This pilot study involved genetic engineering of the C-C chemokine receptor type 5 (CCR5) gene in human embryos, with the intention of conferring HIV-resistance, as seen by a naturally occurring CCR5Δ32 mutation in a few individuals (108). However, based on the limited evidence, CRISPR/Cas9 was likely used to target this gene, but rather than replicate the naturally observed and beneficial 32-base deletion, the edits merely induced DSBs at one end of the deletion, allowing NHEJ to repair the damaged DNA while introducing random, uncharacterized mutations.
Thus, it is unknown whether the resultant protein will function similarly to the naturally occurring CCR5Δ32 protein and confer HIV resistance. In addition, only one of the two embryos, termed with the pseudonym Nana, had successful edits in both copies of the CCR5 gene, whereas the other embryo, with pseudonym Lulu, had successful editing in only one copy. Despite these findings, both embryos were implanted back into their mother, knowing that the HIV-resistance will be questionable in Nana and non-existent in Lulu (109, 110).
Furthermore, recent studies have shown that the mechanism for infection of some variants of the highly mutable HIV virus may heavily rely on the C-X-C chemokine receptor type 4 (CXCR4) co-receptor (108, 111). With no attempts at editing CXCR4, this adds yet another layer of skepticism toward achieving HIV resistance by this strategy. In addition, OTEs, particularly over the lifetime of an individual, remain a major concern for applying this technology in humans.
The recent advances in the editing tool to limit OTEs, such as using high fidelity Cas9 variants, has not been exploited. Furthermore, the rationale for selecting HIV prevention for the first use of CRISPR in implanted human embryos contributes to the poor risk to benefit ratio of this study, considering HIV patients can live long, healthy lives on a drug regimen.
A more appropriate first attempt would have been to employ this technology for a more severe disease. For example, correction of the MYBPC3 gene is arguably a better target for embryonic gene editing, as mutations in MYBPC3 can cause hypertrophic cardiomyopathy (HCM), a heart condition responsible for most sudden cardiac deaths in people under the age of 30.
Gene correction for this pathological mutation was achieved recently for the first time in the US in viable human embryos using the HDR-mediated CRISPR/Cas9 system. However, these embryos were edited for basic research purposes and not intended for implantation. In this study, sperm carrying the pathogenic MYBPC3 mutation and the CRISPR/Cas9 machinery as an RNP complex were microinjected into healthy donor oocytes arrested at MII, achieving 72.4% homozygous wildtype embryos as opposed to 47.4% in untreated embryos.
The HDR-mediated gene correction was observed at considerably high frequencies with no detectable OTEs in selected blastomeres, likely owing to the direct microinjection delivery of the RNP complex in the early zygote. Interestingly, the maternal wildtype DNA was used preferentially for templated repair over the provided exogenous ssODN template (112).
While evidence for gene correction was promising, NHEJ mediated DNA repair was still observed in many embryos, highlighting the need to improve HDR efficiency before clinical application can be considered. Although strategies have been developed to improve HDR, such as chemical inhibitors of NHEJ (77–79), such techniques may have varying outcomes in embryonic cells and side effects that may arise from treatment needs to be investigated. Germline gene editing will remain to be ethically unfavorable at its current state and its discussions may not be considered until sufficient long-term studies of the ongoing somatic CRISPR therapy clinical trials are evaluated.
Potential for CRISPR Therapeutics During COVID-19 Pandemic
The rapidly advancing CRISPR technology may provide aid during our rapidly evolving times. The recent outbreak of a novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has led to a global pandemic (113). These pressing times call for an urgent response to develop quick and efficient testing tools and treatment options for coronavirus disease 2019 (COVID-19) patients.
Currently available methods for testing are relatively time consuming with suboptimal accuracy and sensitivity (114). The two predominant testing methods are molecular testing or serological testing. The US Centers for Disease Control and Prevention (CDC) has developed a real-time RT-PCR assay for molecular testing for the presence of viral RNA to detect COVID-19 (115). However, this assay has a roughly ~30% false negative rate (116, 117) with the turnaround time of several hours to >24 h. Serological testing methods are much more rapid but lack the ability to detect acute respiratory infection since antibodies used to detect infection can take several days or weeks to develop.
Recently, a CRISPR Cas12-based assay named SARS-CoV-2 DETECTR has been developed for detection of COVID-19 with a short turnaround time of about 40 min and a 95% reported accuracy. The assay involves RNA extraction followed by reverse transcription and simultaneous isothermal amplification using the RT-LAMP method. Cas12 and a guide RNA against regions of the N (nucleoprotein) gene and E (envelope) gene of SARS-CoV-2 are then targeted, which can be visualized by cleavage of a fluorescent reporter molecule.
The assay also includes a laminar flow strip for a visual readout, where a single band close to where the sample was applied indicates a negative test and 2 higher bands or a single higher band would indicate cleavage of the fluorescent probe and hence positive for SARS-CoV-2 (118).
In addition to CRISPR’s diagnostic utility, CRISPR may provide therapeutic options for COVID-19 patients. The recently discovered Cas13 is an RNA-guided RNA-targeting endonuclease may serve as a potential therapeutic tool against COVID-19. PAC-MAN (Prophylactic Antiviral CRISPR in huMAN cells) has been developed, which utilizes the Ruminococcus flavefaciens derived VI-D CRISPR-Cas13d variant, selected for its small size facilitating easier packaging in viral vehicles, high specificity, and strong catalytic activity in human cells.
This technique was developed to simultaneously target multiple regions for RNA degradation, opening the door for a much-needed pan-coronavirus targeting strategy, given the evidence suggesting relatively high mutation and recombination rates of SARS-CoV-2 (119). With these advances, the CRISPR/Cas machinery may again be implemented to serve its original purpose as a virus-battling system to provide aid during this pandemic.
reference link: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7427626/
Journal information: Cell



{kind=link}