










 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



The vast majority of individuals infected with mild-to-moderate COVID 19 mount a robust antibody response that is relatively stable for at least five months, according to research conducted at the Icahn School of Medicine at Mount Sinai and published October 28, in the journal Science.
Additionally, the research team found that this antibody response correlates with the body’s ability to neutralize (kill) SARS-CoV-2, the virus that causes COVID-19.
“While some reports have come out saying antibodies to this virus go away quickly, we have found just the opposite – that more than 90 percent of people who were mildly or moderately ill produce an antibody response strong enough to neutralize the virus, and the response is maintained for many months,” said Florian Krammer, Ph.D., Professor of Vaccinology at the Icahn School of Medicine at Mount Sinai and a senior author of the paper.
“Uncovering the robustness of the antibody response to SARS-CoV-2, including its longevity and neutralizing effects, is critically important to enabling us to effectively monitor seroprevalence in communities and to determining the duration and levels of antibody that protect us from reinfection. This is essential for effective vaccine development.”
Study findings are based on a dataset of 30,082 individuals, who were screened within the Mount Sinai Health System between March and October, 2020.
The antibody test used in this research – an enzyme-linked immunosorbent assay (ELISA) – is based on the virus’s telltale spike protein that contains the machinery that enables it to attach and gain entry into our cells.
The ELISA assay was developed, validated, and launched at Mount Sinai by a team of internationally renowned researchers and clinicians. The Mount Sinai antibody test detects the presence or absence of antibodies to SARS-CoV-2 and, importantly, is capable of measuring the titer (level) of antibodies an individual has.
The high sensitivity and specificity of this test – meaning that a false negative or false positive is highly unlikely – allowed it to be among the first to receive emergency use authorization from New York State and the U.S. Food and Drug Administration.
In late March, Mount Sinai began to screen individuals for antibodies to SARS-CoV-2 in order to recruit volunteer donors for its convalescent plasma therapy program – one of the first such programs in the nation. The Clinical Laboratories of The Mount Sinai Hospital set up antibody test results using distinct dilutions set at 1:80, 1:160, 1:320, 1:960 or ≥ 1:2880.
The antibody titer score is generated by the number of times the scientist can dilute a patient’s serum and still be able to detect the presence of antibodies.
Titers of 1:80 and 1:160 were categorized as low titers; 1:320 moderate; and 1:960 or ≥ 1:2880 were high.
By early October, Mount Sinai had screened 72,401 individuals with a total of 30,082 being positive (defined as detectible antibodies to the spike protein at a titer of 1:80 or higher). Of the 30,082 positive samples, 690 (2.29 percent) had a titer of 1:80; 1453 (4.83 percent) of 1:160; 6765 (22.49 percent) of 1:320; 9564 (31.79 percent) of 1:960; and 11610 (38.60 percent) of 1:2880. Thus, the vast majority of positive individuals had moderate-to-high titers of anti-spike antibodies.
“Our microbiology colleagues generated great science and tools that were brought from the research lab into the clinical laboratory, where we were able to implement robust and compliant diagnostic tests at an unprecedented pace,” said Carlos Cordon-Cardo, MD, Ph.D., Irene Heinz Given and John LaPorte Given Professor and Chair of Pathology, Molecular and Cell-Based Medicine and last author of the paper.
“The tireless efforts of so many have enabled us to uncover knowledge that can help inform COVID-19 policy and aid in vaccine development.”
Determining the neutralizing effects of SARS-CoV-2 is critical to understanding the possible protective effects of the immune response. The research team performed a well-established, quantitative microneutralization assay based on authentic SARS-CoV-2 with 120 samples of known ELISA titers ranging from “negative” to ?1:2880.
They found that approximately 50 percent of sera in the 1:80-1:160 titer range had neutralizing activity; 90 percent in the 1:320 range had neutralizing activity; and all sera in the 1:960 to ?1:2880 range had neutralizing activity.
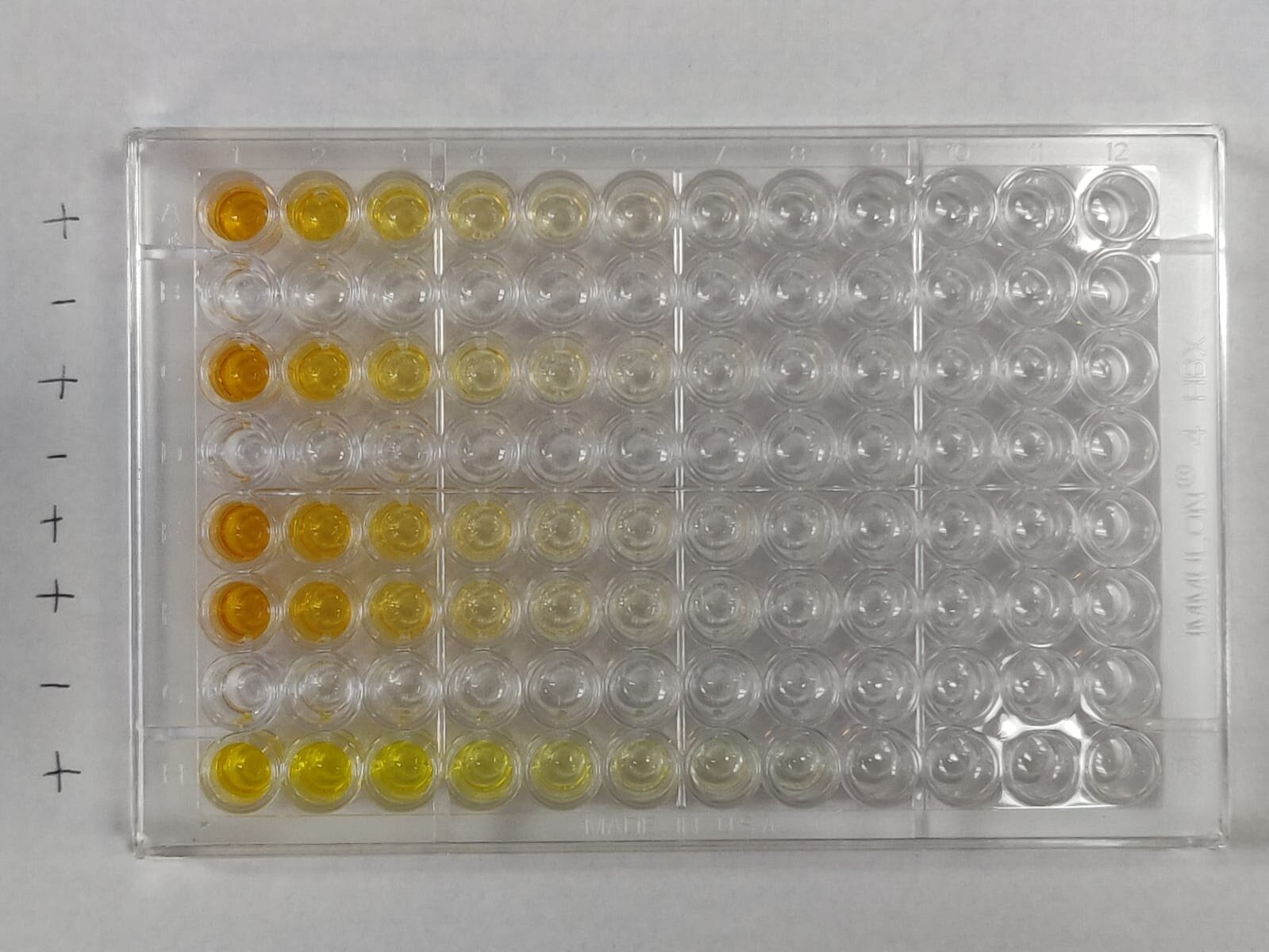
Another important and outstanding question in the scientific community is the longevity of the antibody response to the spike protein. To answer that question, the team recalled 121 plasma donors at a variety of titer levels for repeat antibody testing at approximately 3 months and 5 months post-symptom onset.
When comparing overall titers, they saw a slight drop from a geometric mean titer (GMT) of 764 to a GMT of 690 from the first to second testing time point and another drop to a GMT of 404 for the last testing time point, indicating that a moderate level of antibody is retained by most people 5 months after symptom-onset.
In the higher titer range, they observed a slow decline in titer over time. Interestingly, they saw an initial increase in titer for individuals who had originally tested as having low to moderate titer levels. This is in agreement with earlier observations from their study group that indicate seroconversion in mild COVID-19 cases might take a longer time to mount.
“The serum antibody titer we measured in individuals initially were likely produced by plasmablasts, cells that act as first responders to an invading virus and come together to produce initial bouts of antibodies whose strength soon wanes,” said Ania Wajnberg, MD, Director of Clinical Antibody Testing at the Mount Sinai Hospital and first author of the paper.
“The sustained antibody levels that we subsequently observed are likely produced by long-lived plasma cells in the bone marrow. This is similar to what we see in other viruses and likely means they are here to stay. We will continue to follow this group over time to see if these levels remain stable as we suspect and hope they will.”
The Mount Sinai data reveals antibody binding titers to the spike protein correlate significantly with neutralization of SARS-CoV-2 and that the vast majority of individuals with antibody titers of 320 or higher show neutralizing activity in their serum that are stable over a period of at least 3 months with only modest declines at the 5-month time point.
Correlates of protection have been established for many different viral infections including influenza, measles, hepatitis A, hepatitis B.
These correlates are usually based on a specific level of antibody acquired through vaccination or natural infection that significantly reduces the risk of re-infection. The team will continue following this study cohort over longer intervals of time.
Although this cannot provide conclusive evidence that these antibody responses protect from re-infection, the team believes it is very likely that the antibodies will decrease the odds of getting reinfected and may attenuate disease in the case of breakthrough infection.
To inform policy for the COVID-19 pandemic and for the benefit of vaccine development, it is imperative to swiftly perform studies to investigate and establish a correlate of protection to SARS-CoV-2. Such investigations are currently being carried out by researchers at the Icahn School of Medicine at Mount Sinai.
SARS-COV-2: VIRULENCE FACTORS AND PATHOLOGICAL EFFECTS
As already mentioned above, COVID-19 is an infection that has shortly become a health problem of global concern. Although COVID-19 is not the first outbreak of a coronaviral disease, neither SARS nor MERS led to such a high number of cases worldwide.
This suggests that SARS-CoV-2 is highly contagious and much more virulent than both SARS-CoV and MERS-CoV.
The continuous expansion of COVID-19 could be explained with the viral-specific characteristics and virulence factors[27]. CoV are enveloped viruses, named for the spikes on their surface that resemble a crown.
Their genome is organized in a positive single-stranded RNA. CoV are divided into four genera: α, β, γ, and δ. Along with SARS-CoV and MERS-CoV, the current coronavirus SARS-CoV-2 is classified as a β-coronavirus.
Its genome sequence is about 88% identical to that of two bat-derived SARS-like viruses, and about 80% and 50% identical to the genome sequences of SARS-CoV and MERS-CoV, respectively[28]. Mutations and recombination of the viral genome frequently occur due to error-prone RNA-dependent-RNA polymerases of the CoV.
These events are closely related to viral adaptation[27]. CoV proteins include structural proteins: Spike (S), envelope (E), nucleocapsid (N), and membrane (M) proteins and some proteins with unknown function.
S protein is a glycoprotein essential for viral entry by attachment and fusion to the cellular membrane (Figure 1). It is the main presented antigen on the viral surface and a target of neutralizing antibodies formed during the humoral immune response to the virus[28].

Virulence factors and immune mechanisms during severe acute respiratory syndrome coronavirus 2 infection. After the recognition of the virus by angiotensin-converting enzyme 2 and/or TMPRSS2 receptors, severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) internalizes in the host cell. Via several secondary messengers, the respiratory mucosal cell is stimulated to secrete cytokines and present viral antigens via major histocompatibility complex (human leukocyte antigen) class II molecules to cytotoxic CD8+ T cells, which further secrete cytokines, such as interferon gamma. Additionally, natural killer cells (not shown) also contribute to the killing of infected host cells along with T cytotoxic cells as a part of cellular immunity against every viral infection. Antigen-presenting cells, such as macrophages and dendritic cells, can present viral particles via human leukocyte antigen class II molecules and prime CD4+ T helper cells, which further differentiate to different Th cells, such as Th17 cells (showed), which secrete a vast majority of cytokines, leading to cytokines storms and acute respiratory distress syndrome. Th cells stimulate B cells to produce antibodies against some SARS-CoV-2 antigens, part of the humoral immunity against the virus. Viral replication and shedding are not shown. ARDS: Acute respiratory distress syndrome; SARS-CoV-2: Severe acute respiratory syndrome coronavirus 2; ACE2: Angiotensin-converting enzyme 2.
SARS-CoV and SARS-CoV-2 enter the host cell by binding to the angiotensin-converting enzyme 2 (ACE-2) receptor, while the dipeptidyl peptidase 4 receptor is required for MERS-CoV entry.
After the virus enters the cell, viral RNA is released and involved in consecutive processes of new viral particle formation, which are then released[29].
ACE-2 receptors are mainly expressed in the vascular epithelium, renal tubular epithelium, and Leydig cells in the testes.
For this reason, SARS-CoV might lead to hypogonadism and harm male fertility.
In the respiratory system, SARS-CoV-2 enters the alveoli by binding to the ACE-2 receptors, predominantly expanded on the type II pneumocytes[30].
Once infected, type II pneumocytes are destroyed, and surfactant production is reduced. Macrophages are then recruited to destroy the damaged tissue of the lungs. Macrophages secrete interleukin (IL)-1, IL-6, and tumor necrosis factor alpha (TNFα), which cause vasodilatation and high temperature and enhance neutrophils and lymphocytes migration to the affected area.
Alveoli edema causes respiratory failure due to blood-gas exchange disturbance. Nonstructural proteins might affect the innate immune response of the host and play a crucial role in the viral virulence and pathophysiology of SARS-CoV-2 infection[30].
In vitro and ex vivo examinations compared the viral tropism of SARS-CoV-2 with that of SARS-CoV, MERS-CoV, and 2009 pandemic influenza H1N1. It showed more extensive infection of bronchial epithelium, ciliated cells, and goblet cells with SARS-CoV-2 than with the other viruses[31].
A robust replication in human bronchus was observed, although ACE-2 receptor expression was relatively low in comparison with the lung parenchyma. However, the endothelium of the blood vessels was not found to be infected[32].
SARS-CoV-2 could also be detected in tears, anal swabs, and stool specimens. Moreover, infection and productive replication of the virus were observed in the conjunctiva and colorectal carcinoma cell lines.
These findings suggest that conjunctiva and the bronchus epithelium might also be portals of infection and also poses the possibility of fecal-oral transmission. Various routes of viral transmission would explain the extensive spread of the SARS-CoV-2 throughout the whole world, causing a pandemic[32].
CoV have developed many mechanisms to avoid the immune system, which allow them to better survive in host cells. One of the multiple strategies is the forming of double-membrane vesicles that lack pattern recognition receptors when shedding, thus avoiding the recognition of their evolutionary old and conservative pathogen-associated molecular patterns, such as double-stranded RNA[33].
Moreover, studies in mice showed that SARS-CoV and MERS-CoV infection might inhibit the most potent anti-viral molecules, such as interferons (IFN)-I (IFN-α and IFN-β)[34,35]. The described mechanisms behind this inhibition in MERS-CoV include blocking of melanoma differentiation-associated protein 5, inhibiting the nuclear transport of IFN regulatory factor 3, suppressing the antigen presentation[29], etc.
Therefore, the ability of SARS-CoV-2 to avoid the action of the immune system is a critical factor in the treatment of current infection and the development of specific drugs.
HUMORAL AND CELLULAR IMMUNE RESPONSE – DANCING ON THE FIRE
The presentation of virus antigens stimulates the humoral and cellular immune response, which are exerted by virus-specific B and T lymphocytes, respectively. Like common acute viral infections, the antibody production against SARS-CoV viruses has a typical immunoglobulin (Ig)M and IgG pattern.
SARS-specific IgM antibodies may disappear at the end of the 3rd mo.
In contrast, the IgG antibody may persist longer, indicating that IgG antibodies are likely to be protective[29]. SARS-specific IgG antibodies are predominantly against S- and N-proteins.
Immunological follow-up revealed a progressive increase in plasma SARS-CoV-2-specific IgM and IgG antibodies from 1st to 3rd wk[36].
Patients with COVID-19 may worsen clinically after some days of illness (especially around day 9), which is accompanied by an increase in the pro-inflammatory response, which often leads to admission to intensive care units and the need for supportive mechanical ventilation.
This secondary deterioration is reminiscent of SARS-CoV, in which 80% of patients with SARS-CoV develop acute respiratory disease at the time of anti-viral IgG seroconversion[37]. IgG seroconversion is associated with two points in the immune response – the disappearance of the virus and the appearance of IgG antibodies. IgM antibodies may persist for some time together with IgG.
Moreover, patients who produce anti-S-neutralizing antibodies at the onset of the disease have a higher risk of death.
The mechanisms behind these observations are not clear, but it is assumed that excessive complement activation may play a role. In addition, a phenomenon called antibody-mediated enhancement of viral infection may be responsible for persistent viral load and subsequently cause a direct or indirect effect on ACE-2 activity in the lung[37] and eventually death.
Compared to humoral responses, there are more studies on cellular immunity in coronavirus infection, which is not surprising, taking into account that the principal and effective immune response towards viral infection is cell-mediated immunity.
A clinical case of mild SARS-2-CoV infection and favorable outcome for the patient revealed that the antibody-producing immune cells, defined by the expression of CD3-CD19+CD27hiCD38hi and CD4+CXCR5+ICOS+PD-1+ Th follicular cells, appear in the blood during viral clearance (day 7; 1.48%) and peak on day 8 (6.91%); and on day 7 (1.98%), increasing on day 8 (3.25%) and day 9 (4.46%), respectively. The peak of both cell subpopulations was significantly higher in the COVID-19 patient than in healthy controls and persisted during convalescence[38].
Cytotoxic T cells also increased rapidly from day 7 (3.57%) to day 8 (5.32%) and day 9 (11.8%), followed by a decrease on day 20, respectively.
Besides, the incidence of CD38+human leukocyte antigen (HLA)-DR+CD8+ T cells was significantly higher in this patient than in healthy subjects (1.47% ± 0.50%)[37]. We have to emphasize once again that activated cytotoxic T cells, representatives of adaptive or so-called specific immunity, along with natural killer (NK) cells, part of innate immunity, are critical players in the cell-mediated immune response that accompanies any viral infection.
Helper CD4+ T cells simultaneously expressing CD38 and HLA-DR increased between day 7 (0.55%) and day 9 (3.33%) in this patient compared to healthy donors (0,63% ± 0.28%), although being in lower percentages than for CD8+ T cells. CD38+HLA-DR+CD8+ T cells produce about 34%-54% more granzymes A and B and perforin.
The appearance and rapid increase of activated CD38+HLA-DR+ T cells, especially CD8+ T cells, on day 7-9 precede the disappearance of symptoms[37].
Analysis of CD16+CD14+ mononuclear cells, which are associated with the immunopathology of COVID-19, showed a lower frequency of these cells in the blood of this patient on days 7, 8, and 9 (1.29%, 0.43%, and 1.47%, respectively), compared to healthy control donors (9.03% ± 4.39%), probably indicative of their entry from the blood to the site of infection. No differences were found in activated HLA-DR+CD3-CD56+ NK cells during infection and compared to healthy levels[37].
However, recent data conclude that even reduced in numbers, Th and Tc cells are overactivated in patients infected with SARS-CoV-2[38]. This significant reduction is also observed in the acute-phase response in COVID-19 patients. However, once presented in the organism, CD4+ and CD8 + anti-virus memory T cells persist in the bloodstream of recovered patients for up to 4 years, even in the absence of viral antigens[39]. Moreover, other studies detected SARS-CoV-S protein-specific memory T memory cells 4 years after the patients` infection[40] and MERS-CoV-specific CD8+ T cells in mice, mainly involved in viral clearance[41]. These findings are a reasonable basis for designing effective vaccines against SARS-CoV-2.
Th17 cell responses are also involved in the immune pathogenesis of COVID-19, mainly with the secretion of various cytokines, such as IL-17, granulocyte-macrophage colony-stimulating factor, IL-21, and IL-22[42]. IL-1b and TNFα, as promoters of human Th17 cell differentiation, are also involved in stimulating vascular permeability and leakage.
Nevertheless, it was shown that patients with a severe form of COVID-19 had increased levels of CCR+ Th17 cells[39]. This may suggest that Th17 cells and their cytokines are also involved in the cytokine storm. Besides, elevated Th17 and Th1 cell responses were also described in MERS-CoV and SARS-CoV patients[43,44].
These results support the hypothesis that enhanced IL-17-related pathways, including higher IL-17 but lower IFNγ and IFNα, are associated with worse outcomes for the patients[39].
As we have some proof of the contributing role of Th17 cells to the cytokine storm, Th17 cells may likely promote pulmonary edema, tissue damage, and lung failure. In line with this, targeting Th17 cells may be beneficial for some patients with COVID, especially in those with a dominant Th17 immune profile[43,45].
Some of the immunological responses that are accompanying the SARS-CoV-2 infection are presented in Figure 1.
CLINICAL MANIFESTATIONS OF COVID–19
A symptomatic COVID-19 case is defined as an infected person with a clinical picture suggestive of COVID-19. On the other hand, an asymptomatic case is an infected person who has not developed any signs or symptoms of COVID-19[46].
The incubation period of the disease is 5-6 d on average but can be up to 14 d.
Critical epidemiological and immunological aspects of the disease are that an infected person may be contagious 1-3 d before the onset of the symptoms[47]. About 40% of COVID-19 patients experience mild clinical course, and another 40% present with moderate disease.
Severe disease that requires oxygen support is observed in about 15% of the patients.
Five percent of the infected ones develop a severe disease that progresses to respiratory failure (ARDS) but also sepsis and septic shock, thromboembolism, multiorgan failure, including acute kidney, and cardiac injury[48].
Advanced age, smoking, as well as comorbidities such as diabetes, hypertension, cardiac and chronic lung disease, cerebrovascular diseases, immunosuppression, and cancer have been reported to predispose to a severe course of COVID-19. Children and infants usually experience mild disease or asymptomatic infection[49].
There are no specific clinical symptoms of COVID-19 that can be taken as a reliable pathognomonic sign. Patients most commonly present with fever, cough (usually dry), fatigue, and anorexia.
Dyspnea is usually seen in severe cases. Other nonspecific symptoms include sore throat, nasal congestion, headache, nausea, as well as diarrhea and vomiting. Loss of smell and taste have also been reported. In older people, some atypical symptoms such as reduced alertness and/or mobility and delirium might be seen[50,51].
COVID-19 may also present with neurological and mental manifestations, including delirium or encephalopathy, agitation, stroke, meningoencephalitis, anxiety, depression, and sleep problems. In many cases, some of these neurological manifestations have been documented without respiratory symptoms[52,53].
A retrospective study of SARS-CoV-2 infected patients showed mild leucopenia in mild cases. Mild to moderate leukocytosis was observed in severe cases with significantly high neutrophil and significantly low lymphocyte count.
In patients with a severe course of the disease, significantly high alanine aminotransferase and aspartate aminotransferase were observed, as well as hypoalbuminemia, elevated concentration of C-reactive protein, lactate dehydrogenase, ferritin, and D-dimer. These laboratory changes are a result of an acute and severe inflammatory response[50].
Research on cellular immunity in SARS-CoV-2 positive patients showed a significant reduction in CD4+ and CD8+ T lymphocytes[29]. A more significant decrease in T cells was observed in severe cases as well as an elevation of the concentration of serum cytokines (IL-2, IL-6, IL-10, and TNFα). Elevation of IL-6 concentrations was also found in moderate cases[50]. On that basis, the cytokine storm is a crucial factor for the clinical course of COVID-19 and the disease severity.
A common complication of SARS-CoV-2 infection is the development of ARDS. The latter is assumed as the leading cause of death in patients with COVID-19, especially among those with underlying diseases and conditions, evaluated as risk factors, smokers, and older age.
ARDS is a result of the cytokine storm – an immunopa-thological event that leads to an uncontrolled systemic inflammatory response from the release of pro-inflammatory cytokines IFNα, IFNγ, IL-1β IL-3, IL-6, IL-12, IL-18, TNFα, and chemokines by the immune cells during SARS-CoV infection[29].
These biologically active substances seriously damage the lung parenchyma and lead to respiratory failure. This severe acute hypoxic status is accompanied by increased pulmonary capillary permeability and alveolar epithelial cell damage[51].
TNFα and IL-6 production in COVID-19 follows a different pattern than the pattern in bacterial sepsis or influenza. Furthermore, it was shown that blocking IL-6 by Tocilizumab restored partially HLA-DR expression and increased the number of circulating lymphocytes[51].
The immune mechanisms in COVID-19 are characterized by two main pathways, IL-6 or IL-1b-driven immune hyperactivation, leading to macrophages activations syndrome (MAS) or immune dysregulation (Figure 2).

The immune dysregulation and macrophage activation syndrome, caused by IL-1b- and IL-6-driven hyperactivation, accompanied by a decrease in many cell types, both leading to acute respiratory distress syndrome. CRP: C-reactive protein; HLA: Human leukocyte antigen; Ig: Immunoglobulin; IL: Interleukin; Th: T helper; TNFα: Tumor necrosis factor alpha.
The initiation of pneumonia in COVID-19 includes intense antigen presentation, accompanied by elevated C-reactive protein (CRP), D-dimer, and liver amino-transferases plus infiltrates in the lungs, whereas severe respiratory failure displays either MAS or deficient HLA-DR expression and profound depletion of Th, B cells, and NK cells[51]. In such a way, during ARDS, CRP, D-Dimer, and liver transaminases are further increased, leading to permanent pathological changes.
The described cytokine storm violently attacks not only the lungs but the organs of every system in the organism, causing multiple organ failure that leads to death in severe cases of SARS-CoV-2, SARS-CoV, and MERS-CoV infection[29].
Affecting the whole organism, the cytokine storm is the primary mechanism that induces disseminated intravascular coagulation (DIC). Pro-inflammatory cytokines TNFα and IL-1 suppress the endogenous anticoagulation.
Inflammation damages the endothelium and leads to the release of tissue plasminogen activator, which could explain the elevation of the D-dimer and fibrin degradation products[54]. In summary, the accumulated data showed that COVID-19 is associated with a hypercoagulable state with increased risk of thromboembolic complications.
However, ARDS is assumed as the leading cause of death in COVID-19. During the early stages of the outbreak, it was reported that of the 41 SARS-CoV-2 infected patients admitted, six died of ARDS[48]. ARDS remains the most common immunopathological event for SARS-CoV-2, SARS-CoV, and MERS-CoV infections.
One of the primary mechanisms for ARDS is the cytokine storm. The latter is a uncontrolled systemic inflammatory reaction resulting from the release of large amounts of pro-inflammatory cytokines (IFN-α, IFN-γ, IL-1β, IL-6, IL-12, IL-18, IL-33, TNF-α, TGFβ, etc.) and chemokines (CCL2, CCL3, CCL5, CXCL8, CXCL9, CXCL10, etc.) from immune effector cells during many viral infection[48,55-57].
Similar to SARS-CoV, individuals with severe MERS-CoV infection show elevated serum levels of IL-6, IFN-α, CCL5, CXCL8, and CXCL-10 compared to those with mild or moderate disease[37]. The cytokine storm causes ARDS and multi-/multiorgan failure, leading to death in severe cases of coronaviral infections[58].
Another hypothesis on the severity of COVID-19 disease includes the problem associated with the activation of bradykinin B1 receptors on the lung endothelial cells. It is assumed that the enzymatic activity of the ACE-2 receptor inactivates des-Arg9 bradykinin, which is a ligand for B1.
Furthermore, unlike B2, B1 is regulated by pro-inflammatory cytokines[59]. Interestingly, without the inactivation of B1 ligands, increased local vascular permeability is observed in lung mucosa, leading to angioedema. Probably, angioedema is a typical manifestation of the severe disease onset and the cause of typical changes visible on the computed tomography scan.
The feeling of choking is observed mainly around day 9, denoted the worsening of the patients[60].
As any common viral infection, here, we also observed a progressive inflammatory condition with elevated IL-6, CRP, and ferritin but not procalcitonin or erythrocyte sedimentation rate, which indicates the second stage of the disease[60].
In line with this, the renin-angiotensin system could also contribute to the lung injuries along with sepsis-associated DIC. However, the levels of platelets, prothrombin time, and fibrinogen may remain normal[61].
The pathophysiology of the disease may be a combination of injury of pulmonary type II pneumocyte, viral pneumonia, ARDS, DIC, sepsis, cytokine storm, MAS, and overall immune dysregulation[62,63]. Some of these aspects are shown in Figure 2.
THERAPY OPTIONS RELATED TO THE IMMUNE MECHANISMS
COVID-19 infection revealed some treatment options related to the immune responses. If we accept the division of SARS-CoV-2 infection into three stages, at the beginning of the infection (stage I, an asymptomatic incubation period with or without detectable virus), some of the mechanisms of innate immunity play a role, including NK cells, interferon production, and some cytokines.
Therefore, strategies to boost immunity, such as administration of anti-serums (ready-made antibodies from survivors) or pegylated IFNα, are undoubtedly crucial at this stage.
During the incubation period, as well as the non-severe stage (stage II, a mild symptomatic period with the presence of a virus in the body), a specific adaptive immune response is required to neutralize and eliminate the virus, which will eventually prevent disease progression to severe stages.
The adaptive response, however, occurs more slowly and is activated at a later stage. When the protective immune response is compromised, the virus will spread, and massive damage of the affected tissues and organs will occur, especially in those that highly express ACE-2 receptors, such as the gut and kidneys.
This leads to lung inflammation, mostly mediated by pro-inflammatory immune cells. Inflammation of the lungs is the leading cause of life-threatening respiratory failure in the severe stage of the disease (stage III, severe respiratory symptomatic stage with high viral load).
Therefore, once severe lung damage occurs, attention should be moved to the inflammation and the efforts to suppress the immune responses and control the symptoms[72]. Some of the treatment options related to the immunological mechanisms could be seen in Figure 3.

Treatment modalities related to the immunological mechanisms observed during coronavirus disease 2019. ACE2: Angiotensin-converting enzyme 2; ARDS: Acute respiratory distress syndrome; MHC: Major histocompatibility complex; MSCs: Mesenchymal stromal/stem cells; SARS-CoV-2: Severe acute respiratory syndrome coronavirus 2.
Antibody and plasma therapy are the next therapy option, closely related to the immune mechanisms. It has been reported that many cured patients donate plasma against SARS-CoV-2, as there were clinical trials for SARS-CoV[73] and MERS-CoV[74].
Preliminary data indicate favorable results for use in patients with acute and severe SARS-CoV-2 infection. In addition, the development of a recombinant human monoclonal antibody, such as CR3022, is a reasonably easy way to neutralize the virus by attachment to the receptor-binding domain of SARS-CoV-2.
This SARS-specific human monoclonal antibody has the potential to be developed as a drug for SARS-CoV-2 infection[75]. Other monoclonal antibodies that neutralize SARS-CoV, such as m396 and CR3014, may be alternatives for the treatment of SARS-CoV-2[76].
Passive antibody therapy is the administration of ready-made antibodies against an infectious agent to a susceptible individual to prevent or treat an infectious disease caused by that microorganism. Thus, the passive application of antibodies is the only means of ensuring the immediate and ready immunity of endangered persons. Experience with previous epidemics with other CoV, such as SARS-CoV-1, has shown that survivors’ sera may contain neutralizing antibodies to the virus, thus the expected mechanism of action of passive antibody therapy is viral neutralization and elimination.
However, other mechanisms, such as antibody-dependent cellular cytotoxicity and/or phagocytosis, are also possible[72].
Currently, the only ready-to-use antibodies are those obtained from survived patients. As more people become ill with COVID-19 and recover, the number of potential donors will continue to increase. The serum of recovered individuals can be used prophylactically to prevent infection in high-risk individuals, such as those with chronic diseases, medical personnel, and those who have been in contact with patients.
The efficacy of this approach is unknown, but historical experience has shown that products containing passive antibodies are more effective in preventing than in treating existing disease.
If used for therapy, antibodies are most effective when administered soon after the onset of symptoms. The antibody acts by altering the inflammatory responses, which is also more easily achieved during the initial immune response, or the asymptomatic stage.
The reason for the differences in efficacy is not well understood. Still, it may reflect the fact that the antibody works by neutralizing the initial inoculum from the infectious agent, which is probably much less than that in an already developed infection and a large number of viral copy. In line with this, to be effective, a sufficient amount of antibodies must be administered.
When given to a person at risk of infection, this antibody will reach the tissues via blood and can provide protection against the infection. Depending on the amount and composition of immunoglobulins, passive protection can last from weeks to months (for IgG).
The challenges, however, are related to the difficulties in that, as we have mentioned above, some patients do not possess high antibody titers after the illness. Therefore, due to the individual variation of the immune responses, the insufficiently active immune response in some people will be a reason for them to be prone to reinfection.
In a prophylactic regimen of the use of passive immunotherapy in persons at risk, the aim is to prevent disease. Used therapeutically, the passive serum is administered to patients with clinical manifestations of the disease to reduce the severity of symptoms and mortality. The efficacy of these approaches cannot be measured without conducting a controlled clinical trial[72].
The risks of passive application of sera products fall into two categories – known and theoretical. The known risks are those associated with the use of blood products – infectious diseases and allergic reactions to the components of the serum, including serum sickness.
With modern blood banking techniques, these risks are low. For sera used for therapy, there are also theoretical risks for transfusion-related acute lung injury and antibody-dependent enhancement (ADE) of infection.
Several mechanisms of ADE have been described for CoV, and there are concerns that antibodies against one type of coronavirus may exacerbate infection to another virus strain. It is possible to experimentally predict the risk of ADE in SARS-CoV-2, as suggested for MERS.
It is assumed that when using sera rich in virus-neutralizing SARS-CoV-2 antibodies, the risk of developing ADE is minimal[72]. However, it is good to test these hypotheses in clinical trials. Another theoretical risk is that the use of antibodies to individuals exposed to SARS-CoV-2 may weaken the immune response so that not enough of their own antibodies would form. This puts these people at risk for subsequent reinfection. However, if the risk proves to be real, these individuals can be vaccinated against COVID-19 when the vaccine becomes available.
The preparation of highly purified and enriched neutralizing antibodies against SARS2-CoV-2 is preferable because they are safer and have higher activity. However, such preparations will not be available in the coming months, while locally derived serums may be available much sooner[77].
Other therapy strategies are oriented against the cytokine storm. Because lymphocytopenia is commonly seen in severe cases of COVID-19, the cytokine storm caused by the SARS-CoV-2 virus may be mediated by leukocytes other than T cells.
High white blood cell counts are common, which, along with lymphocytopenia, is used as a differential diagnostic criterion for COVID-19. In any case, the blocking of IL-6 can be effective, as well as the blocking of IL-1 and TNFα.
The approach to blocking these pro-inflammatory cytokines has been adopted in various autoimmune diseases. We also proposed anti-IL-6 therapy in IBD patients as a way of limiting the inflammation and development of colorectal carcinoma[78].
Anti-inflammatory strategies, including the blockade of specific cytokines that increase B1 expression on endothelial cells locally at the site of inflammation in combination with B1 and or B2 receptor blockade, should be considered.
IL-1 (consisting of IL-1α and IL-1β) and TNF are potent inducers of the B1 receptor. Blocking the translocation of nuclear factor kappa B, TNF-α, or IL-1 prevents the regulation of B1 receptors both functional and molecular by lipopolysaccharide.
Therefore, one strategy could be the treatment with anakinra, a monoclonal antibody that blocks not only IL-1α but also IL-1β, and has an excellent safety profile.
IL-1a is probably in extremely high concentrations locally due to its release from damaged cells. TNF blockade is an option but is associated with many more infectious complications. Also, complement activity has been described and may play a role in this stage of the disease.
Moreover, it may be affected by blockade of C5 component with eculizumab, a monoclonal antibody that was randomized to COVID-19 (NCT04288713)[60]. The use of antibodies against cytokines is a general approach in several autoimmune and autoinflammatory diseases in which one or more cytokines play a direct role in the pathogenesis. However, this approach is not without any side effects, so the benefit should always be considered.
On the other hand, a significant drawback is the fact that blocking of a cytokine rarely has the desired effect because the cytokines are connected in a network. Corticosteroids are also an option for therapy in these cases. Anti-inflammatory strategies may provide time, but will not cure the disease on their own if the virus is present or bradykinin-associated angioedema is not controlled[60].
Although various clinics in China have proclaimed the use of mesenchymal stromal/stem cells (MSCs) in severe cases of COVID-19 infection, no reliable results have yet been published. One of the advantages of this therapy is that MSCs must be activated by IFNγ. After that, MSCs are capable of exerting their anti-inflammatory effects.
By producing various growth factors, MSCs can help repair damaged lung tissue. However, severely affected patients may lack these MSCs properties because T cells do not activate well in SARS-CoV-2 infection. The use of a “licensing approach” is being considered: Pre-treatment of MSCs with IFNγ with or without TNFα or IL-1.
Thus, cytokine-treated MSCs may be more effective in suppressing the hyperactive immune response and promoting tissue repair, as such MSCs are useful in acute lung damage caused by lipopolysaccharide[72]. In any case, the use of MSCs in clinical practice should be limited to strict indications, taking into account the benefit-risk for each patient. Their oncogenic potential should not be neglected.
reference link : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7520644/
More information: Robust neutralizing antibodies to SARS-CoV-2 infection persist for months, Science 28 Oct 2020: eabd7728. DOI: 10.1126/science.abd7728 , science.sciencemag.org/content … 0/27/science.abd7728



{kind=link}