











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



Brown fat is that magical tissue that you would want more of. Unlike white fat, which stores calories, brown fat burns energy and scientists hope it may hold the key to new obesity treatments.
But it has long been unclear whether people with ample brown fat truly enjoy better health. For one thing, it has been hard to even identify such individuals since brown fat is hidden deep inside the body.
Now, a new study in Nature Medicine offers strong evidence: among over 52,000 participants, those who had detectable brown fat were less likely than their peers to suffer cardiac and metabolic conditions ranging from type 2 diabetes to coronary artery disease, which is the leading cause of death in the United States.
The study, by far the largest of its kind in humans, confirms and expands the health benefits of brown fat suggested by previous studies. “For the first time, it reveals a link to lower risk of certain conditions,” says Paul Cohen, the Albert Resnick, M.D., Assistant Professor and senior attending physician at The Rockefeller University Hospital.
“These findings make us more confident about the potential of targeting brown fat for therapeutic benefit.”
A valuable resource
Although brown fat has been studied for decades in newborns and animals, it was only in 2009 that scientists appreciated it can also be found in some adults, typically around the neck and shoulders. From then on, researchers have scrambled to study the elusive fat cells, which possess the power to burn calories to produce heat in cold conditions.
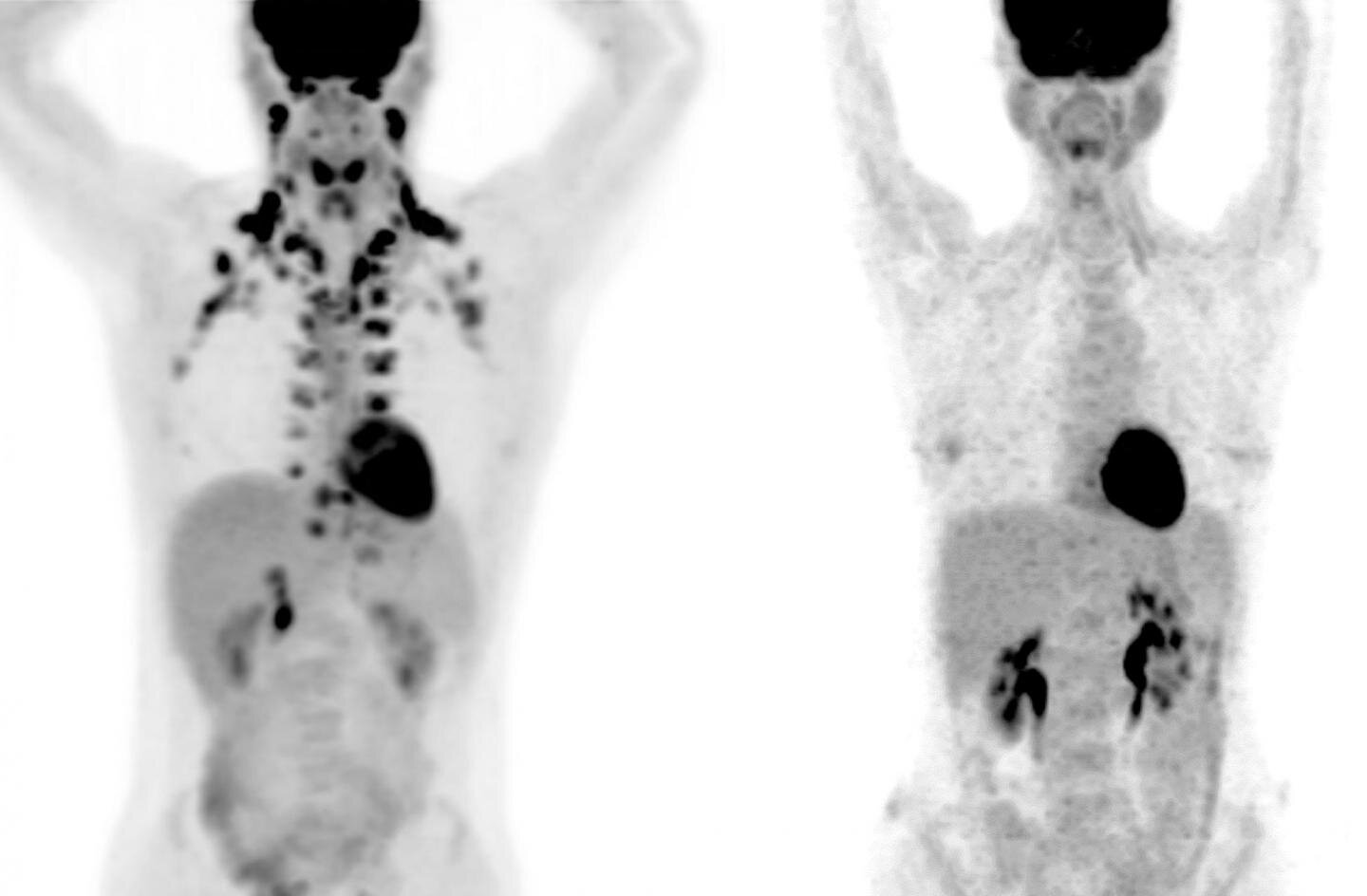
Large-scale studies of brown fat, however, have been practically impossible because this tissue shows up only on PET scans, a special type of medical imaging.
“These scans are expensive, but more importantly, they use radiation,” says Tobias Becher, the study’s first author and formerly a Clinical Scholar in Cohen’s lab. “We don’t want to subject many healthy people to that.”
A physician-scientist, Becher came up with an alternative. Right across the street from his lab, many thousands of people visit Memorial Sloan Kettering Cancer Center each year to undergo PET scans for cancer evaluation. Becher knew that when radiologists detect brown fat on these scans, they routinely make note of it to make sure it is not mistaken for a tumor. “We realized this could be a valuable resource to get us started with looking at brown fat at a population scale,” Becher says.
Protective fat
In collaboration with Heiko Schoder and Andreas Wibmer at Memorial Sloan Kettering, the researchers reviewed 130,000 PET scans from more than 52,000 patients, and found the presence of brown fat in nearly 10 percent of individuals. (Cohen notes that this figure is likely an underestimate because the patients had been instructed to avoid cold exposure, exercise, and caffeine, all of which are thought to increase brown fat activity).
Several common and chronic diseases were less prevalent among people with detectable brown fat. For example, only 4.6 percent had type 2 diabetes, compared with 9.5 percent of people who did not have detectable brown fat. Similarly, 18.9 percent had abnormal cholesterol, compared to 22.2 percent in those without brown fat.
Moreover, the study revealed three more conditions for which people with brown fat have lower risk: hypertension, congestive heart failure, and coronary artery disease – links that had not been observed in previous studies.
Another surprising finding was that brown fat may mitigate the negative health effects of obesity.
In general, obese people have increased risk of heart and metabolic conditions; but the researchers found that among obese people who have brown fat, the prevalence of these conditions was similar to that of non-obese people. “It almost seems like they are protected from the harmful effects of white fat,” Cohen says.
More than an energy burning powerhouse
The actual mechanisms by which brown fat may contribute to better health are still unclear, but there are some clues. For example, brown-fat cells consume glucose in order to burn calories, and it’s possible that this lowers blood glucose levels, a major risk factor for developing diabetes.
The role of brown fat is more mysterious in other conditions like hypertension, which is tightly connected to the hormonal system. “We are considering the possibility that brown fat tissue does more than consume glucose and burn calories, and perhaps actually participates in hormonal signaling to other organs,” Cohen says.
The team plans to further study the biology of brown fat, including by looking for genetic variants that may explain why some people have more of it than others—potential first steps toward developing pharmacological ways to stimulate brown fat activity to treat obesity and related conditions.
“The natural question that everybody has is, ‘What can I do to get more brown fat?'” Cohen says. “We don’t have a good answer to that yet, but it will be an exciting space for scientists to explore in the upcoming years.”
Brown adipocytes, which reside in specific depots called brown adipose tissues (BAT), produce heat in a process called non-shivering thermogenesis. Thermogenesis in BAT is stimulated mainly by the sympathetic nervous system in response to cold exposure, and it helps maintain body temperature (euthermia) in placental mammals.
The acquisition of BAT in early mammalian evolution is considered one key evolutionary advantage that allowed for the successful expansion of mammals, and its functional importance in newborn humans and small rodents has long been appreciated.
More recently, it has become apparent that adult humans also have functionally relevant BAT, and possibly the additional capacity to induce the formation of brown-like adipocytes within white adipose tissues (WAT) (called brite or beige adipocytes) under certain conditions. Because these thermogenic cells, when active, have a high rate of nutrient consumption and energy expenditure, their existence in adult humans not only correlates with improved metabolic profiles (Betz and Enerback, 2018), but has stimulated interest in targeting them therapeutically to fight obesity and improve glycemic control (Hanssen et al., 2015, Ouellet et al., 2012, Yoneshiro et al., 2011b, Yoneshiro et al., 2011c).
This has gone hand-in-hand with renewed interest in understanding the basic biological mechanisms of brown fat development and metabolic regulation, which includes understanding the cellular lineages and precursor cell pools that give rise to brown and brite/beige adipocytes, and the signals that govern their fuel selection and unique metabolism.
Identifying brown adipocyte stem and progenitor cells, and elucidating the mechanisms that stimulate their differentiation into mature thermogenic adipocytes, could have important implications in developing brown fat based therapeutics. Here, we will discuss our present understanding of brown adipocyte development and function, the related topic of brite/beige adipocytes, and key future goals and unanswered questions especially as they relate to potential therapies.
Basics of Non-shivering thermogenesis
Cold-stimulated non-shivering thermogenesis (NST) in the brown adipocyte is dependent upon the intrinsic expression and function of uncoupling protein 1 (UCP1), an inner mitochondrial membrane transporter that dissipates the energy stored in the mitochondrial electrochemical gradient as heat, “uncoupled” from ATP synthesis (Betz and Enerback, 2018).
In the absence of thermal stress, brown adipocyte UCP1 is thought to be inhibited by purine nucleotides (Nicholls, 2006, Sluse et al., 2006). During cold stress, brown fat thermogenesis is classically stimulated by norepinephrine released from the sympathetic nervous system (SNS), which activates β3-adrenergic receptors on brown adipocytes to stimulate intracellular synthesis of the second messenger cyclic AMP (cAMP), leading to cAMP-driven protein kinase A (PKA) signaling activation.
This stimulates lipid catabolism processes such as lipolysis which liberates free fatty acids from triacylglycerol lipid storage droplets, and increases expression of a thermogenic gene expression program that includes UCP1 mRNA (Nicholls, 2006, Sluse et al., 2006, Fedorenko et al., 2012, Lehr et al., 2006).
Exactly how brown adipocytes choose and utilize fuel remains an important and open question. Recent studies suggest that active lipolysis in brown adipocytes may not be required for sustaining thermogenesis so long as exogenous lipids are available; nevertheless, cellular free fatty acids reportedly directly activate UCP1 (Fedorenko et al., 2012, Shin et al., 2017, Schreiber et al., 2017).
Active brown adipocytes also take up glucose from circulation, and synthesize free fatty acids de novo from glucose and possibly other lipogenic precursors (i.e. the process of de novo lipogenesis) to continuously fuel NST or to provide other yet to be appreciated metabolic advantages (Sanchez-Gurmaches et al., 2018, McCormack and Denton, 1977, Mottillo et al., 2014, Shimazu and Takahashi, 1980, Trayhurn, 1979, Yu et al., 2002).
In addition, BAT thermogenesis is fueled by liver-derived plasma lipid metabolites (acyl-carnitines), the release of which is stimulated by cold-induced lipolysis in the WAT (Simcox et al., 2017). It has also been suggested recently that UCP1-independent mechanisms of thermogenesis might exist under certain circumstances (Bertholet et al., 2017, Ikeda et al., 2018, Kazak et al., 2015).
Brown adipocytes might also have key metabolic functions in addition to thermogenesis, such as secreting special adipokines (called BATokines) and exosome containing miRNAs that might have both autocrine function and paracrine functions on nearby immune cells, as well as endocrine functions related to glucose homeostasis and cardiovascular health (Thomou et al., 2017, Villarroya et al., 2013, Hansen et al., 2014, Svensson et al., 2016, Long et al., 2016, Wang et al., 2014a, Villarroya and Giralt, 2015).
Brown Fat Anatomy and Morphology
The color distinction between a “brown” and a “white” adipocyte largely reflects the many more mitochondria (which are high in iron) in brown adipocytes compared to white adipocytes (Figure 1). A stimulated brown adipocyte actively generating heat also contains many small lipid droplets and is referred to as being multi-locular, while white adipocytes, such as those in subcutaneous and visceral depots (sWAT and vWAT, respectively) typically have a single large unilocular lipid droplet (Figure 1). Having many small lipid droplets increases lipid droplet surface area and presumably promotes metabolite exchange with mitochondria (Blanchette-Mackie and Scow, 1983, Benador et al., 2018).
A less active brown adipocyte that is not engaged in thermogenesis (e.g. after acclimation to thermoneutrality) adopts a morphology more similar to a white adipocyte although it retains an epigenetic cellular identity that differentiates it from a white adipocyte (Roh et al., 2018).

General characteristics of brown, white and brite/beige adipocytes.
A stimulated brown adipocyte (left) contains numerous small lipid droplets, many mitochondria, and expresses high levels of uncoupling protein 1 (UCP1), which is embedded in the inner mitochondrial membrane and required for thermogenesis. The color of brown fat reflects the high iron content of mitochondria. A white adipocyte (middle) in contrast contains a single large lipid droplet, fewer mitochondria, and does not express UCP1. A Brite/beige adipocyte (right) is characteristically intermediate between brown and white adipocyte, having multiple lipid droplets (though often larger than those seen in a brown adipocyte), more mitochondria than a white adipocyte, and it expresses UCP1.
As indicated above, brown adipocytes exist in defined BAT depots in the mouse, which is the main model organism used to study brown fat. Notably, the size and composition of each BAT depot differs with age, gender and mouse strain background (Frontini and Cinti, 2010, Murano et al., 2009).
The largest BAT depots are clustered in the dorsal anterior regions of the mouse body and include the inter-scapular (iBAT), sub-scapular (sBAT) and cervical depots (cBAT) (Frontini and Cinti, 2010, de Jong et al., 2015, Walden et al., 2012, Cinti, 2005) (Figure 2). In addition, there are several small BAT depots proximal to major blood vessels and specific organs, such as the peri-aortic BAT depot (paBAT) that aligns to aortic vessels, and the peri-renal BAT depot (prBAT) that localizes in a fibrous capsule of the kidney (Frontini and Cinti, 2010) (Figure 2).
Recent studies using 18F-FDG PET/CT or (123/125I)-β-methyl-p-iodophenyl-pentadecanoic acid with SPECT/CT imaging, which traces glucose and lipid uptake respectively, suggests additional small pockets of cold responsive fat depots exist in suprascapular, supraspinal, infrascapular, and ventral spinal regions (Zhang et al., 2018, Mo et al., 2017).

Adipose tissue anatomy and plasticity
(A) Cartoons showing brown and white fat depots in mice that are acclimated to thermoneutrality (30°C ~ 32°C), mild cold (20°C ~ 22°C), and severe cold (6°C ~ 10°C). The color and size of each depot is modeled such that it reflects the observed differences in mice acclimated to each temperature. A key showing the gradient of “browning” or “britening/beiging” is provided below each model. (B) Hematoxylin and Eosin staining of the indicated brown and white fat depots at each temperature. Note that at thermoneutrality, brown adipocytes contain larger single lipid droplets. At 20–22°C, the standard mouse facility temperature, brown adipocytes exhibit their stimulated morphology of being multi-locular (see Figure 1) while white adipocytes remain unilocular though SWAT adipocyte size is reduced likely reflecting in part a higher level of lipolysis that is necessary to fuel the active brown fat depots. At severe cold temperatures, (6–10°C), additional morphological changes can been see in BAT (i.e. lipid droplets become more uniform), and under these conditions, brite/beige adipocytes also from in the subcutaneous WAT. Of note, the browning capacity of WAT depots is not dependent on a depot being subcutaneous or visceral because, for example, the retroperitoneal visceral WAT depot has high britening/beiging capacity (not shown) while the perigonadal visceral WAT (shown) does not. [Abbreviations] iBAT, interscapular BAT; sBAT, subscapular BAT; cBAT, cervical BAT; paBAT, peri-aortic BAT; prBAT, peri-renal BAT; asWAT, anterior subcutaneous WAT; psWAT, posterior subcutaneous WAT; mWAT, mesenteric WAT; rWAT, retroperitoneal WAT; pgWAT, perigonadal WAT. The images in this figure are based primarily on experiments with C57Bl/6 mice.
Similar to the mouse, newborn humans have active brown adipocytes present at birth in large interscapular BAT depots and peri-renal depots (Figure 3A), which presumably helps maintain core body temperature though could also have other neonatal functions not yet appreciated. Until recently, it was widely believed that after neonatal BAT recedes, adult humans lacked brown fat.
However, about a decade ago the widespread existence of active BAT in adults was revealed by retrospective analyses of 18F-fluodeoxyglucose (FDG) uptake assays, which uses positron emission tomography/computed tomography (PET-CT) to measure glucose uptake into organs (Yoneshiro et al., 2011a, Yoneshiro et al., 2013, van der Lans et al., 2013, Ouellet et al., 2012, Hanssen et al., 2015, Nedergaard et al., 2007, Cypess et al., 2009, van Marken Lichtenbelt et al., 2009, Saito et al., 2009, Virtanen et al., 2009, Kortelainen et al., 1993).
These studies also revealed a correlation between BAT activity/amount and metabolic fitness. More recent studies show that BAT depots in adult humans exist in the supraclavicular, axillar and paravertebral regions, though the variability across individuals and populations is still being worked out (Zhang et al., 2018, Nedergaard et al., 2007, Cypess et al., 2009, van Marken Lichtenbelt et al., 2009, Virtanen et al., 2009, Ouellet et al., 2012) (Figure 3B).
There are also small BAT depots in perivascular regions (aorta, common carotid artery), and near the heart wall (epicardium), lung bronchia, and some solid organs (hilum of kidney and spleen, adrenal, pancreas, liver) (Sacks and Symonds, 2013) (Figure 3B).

Brown fat locations in humans(A) Newborn infants have large interscapular and peri-renal BAT depots. (B) In adults, smaller BAT depots are located in the cervical, supraclavicular, axillary, peri-aortic, paravertebral and suprarenal regions. The mapping of these depots in adults is largely based on glucose uptake measurements by 18F-FDG-PET/CT imaging, which shows increased glucose flux at colder temperatures (shown in figure) and on post-mortem resections. The molecular and functional nature of individual (putative) BAT depots remains unclear in humans. Also note that the amount of BAT is highly variable between individuals, but when active BAT is present, it has been shown to correlate with improved metabolism (not shown, discussed in text). Emerging advances in BAT imaging will likely confirm additional depots.
BAT vascularization and innervation
Brown fat depots are also highly vascularized, which facilitates the exchange of oxygen and nutrients and the dissipation of heat and release of BATokines into circulation (Bartelt et al., 2011, Labbe et al., 2015, Sacks and Symonds, 2013). In fact, BAT requires increased blood infusion rate during BAT recruitment (i.e. cold stimulation) to obtain sufficient metabolic substrates and oxygen.
Brown adipocytes also generate vascular endothelial growth factor-A (VEGF-A) and nitric oxide (NO), which facilitates BAT angiogenesis and vascularization (Xue et al., 2009, Sun et al., 2014, Nisoli et al., 1998, Mahdaviani et al., 2016), a process that is reduced in obese mice resulting in loss of thermogenic activity (Shimizu et al., 2014). Other recent work suggests that brown adipocytes may have a vasoprotective role that might be mediated by the secretion of hydrogen peroxide (H2O2), which inhibits vessel contractions in nearby vascular cells (Friederich-Persson et al., 2017).
In addition to being highly vascularized, BAT is extensively innervated allowing for its rapid stimulation by the sympathetic nervous system (SNS). The SNS releases catecholamines such as norepinephrine that activate G-protein coupled β3-adrenergic receptors that are highly expressed on mature brown adipocytes, and β1-adrenergic receptors on brown adipocyte precursors (Cannon and Nedergaard, 2004, Morrison et al., 2012, Bukowiecki et al., 1986, Bronnikov et al., 1992).
While β3-adrenergic receptor signaling stimulates mature brown adipocyte lipid catabolic activity and thermogenesis, β1-adrenergic receptor signaling stimulates brown fat adipogenesis upon prolonged cold challenge (Bronnikov et al., 1992). Classic denervation studies reveal the indispensability of the SNS connections for thermogenesis (Silva and Larsen, 1983, Rothwell and Stock, 1984, Takahashi et al., 1992, Labbe et al., 2015). Emerging research also suggests that innervation may also be critical for BAT to communicate directly with other non-SNS tissues, such as the WAT (Schulz et al., 2013, Garretson et al., 2016, Nguyen et al., 2018).
In summary, BAT is a dynamic and heterogeneous tissue, and the extensive networks of vessels and nerves found in BAT suggests that during brown fat development, there is tight coordination between brown adipocyte precursors (discussed below), endothelial lineages, and nerve cell lineages, and likely immune cells too (Lumeng and Saltiel, 2011, Olefsky and Glass, 2010, Villarroya et al., 2018). The signaling and metabolic interactions between different cell lineages during brown fat development has not yet been extensively studied by systems based approaches.
Transcriptional control of brown adipocyte differentiation
Much of the general transcriptional cascade that promotes adipogenesis is shared between brown and white adipocytes, and has been studied at length using in vitro models (e.g. 3T3–L1 cells). The master regulator of adipogenesis, PPARγ, is both necessary and sufficient for adipogenesis (Rosen et al., 1999, Tontonoz et al., 1994, Wang et al., 2013a). Other key components of the general adipogenesis transcriptional cascade also important in brown and brite/beige adipocyte differentiation include the members of the C/EBP family (C/EBPα, C/EBPβ, C/EBPδ)(Farmer, 2006).
While PPARγ is the dominant factor, overexpression of all C/EBP family members induces adipocyte formation. In culture, C/EBPβ and C/EBPδ function in the first wave of adipogenic transcription factors (hours after adipogenic induction) that eventually triggers a second wave (days after adipogenic induction) that includes C/EBPα and PPARγ, which feed-forward activate themselves (Farmer, 2006).
More recently, efforts to identify brown adipocyte lineage specific transcription factors has identified new additional components that may contribute to the brown (or brite/beige) adipocyte fate. PRDM16 (PRD1-BF1-RIZ1 homologous domain containing 16) was originally described as a BAT transcriptional determination factor that induces a robust thermogenic adipocyte phenotype in white adipocytes both in vitro and in vivo, and can direct muscle precursors to differentiate into brown adipocytes in vitro (Seale et al., 2008, Seale et al., 2007, Seale et al., 2011).
In vivo, other PRDM family members can compensate for the loss of PRDM16 in BAT precursors to maintain normal BAT formation (Harms et al., 2014). In addition, the EBF2 (early B-cell factor 2) transcription factor is selectively expressed in both BAT and brite/beige precursors, and it is required for BAT identity and efficient brite/beige cell formation (Rajakumari et al., 2013, Stine et al., 2016, Wang et al., 2014b).
Recent studies also identified zinc finger protein 516 (Zfp516), whose expression in brown fat is markedly increased in response to cold exposure or β-adrenergic stimulation via β-AR-cAMP pathway, and it directly interacts with PRDM16 to promote BAT development and WAT browning while suppressing myogenesis (Dempersmier et al., 2015, Sambeat et al., 2016). Whether there are brite/beige specific transcription factors that do not function in brown adipocyte lineages remains an important area of investigation.
In contrast to pro-thermogenic transcription factors, less is known about the transcriptional machinery that promotes and/or maintains the white adipocyte phenotype. One interesting candidate is Zfp423, which has recently emerged as a critical brake that prevents white adipocytes from converting to thermogenic adipocytes. Zfp423 is expressed in white adipocyte precursor cells and functions to block the brite/beige thermogenic program by inhibiting the EBF2 and PRDM16 (Gupta et al., 2010, Gupta et al., 2012, Shao and Gupta, 2018, Shao et al., 2016).
While these studies are opening the door to our understanding of adipocyte fate determination at the level of gene expression, there is still much to be learned especially if this information is to be harnessed for therapeutic opportunities. Moreover, other key gene expression factors that contribute to fate decisions, such as epigenetic marks and higher order chromatin regulation, are just beginning to be explored (Roh et al., 2017, Zhao et al., 2016, Carrer et al., 2017, Roh et al., 2018) making this an important area of investigation for many years to come.
Brown Adipocyte Origins
Brown adipocytes are thought to originate from the mesoderm during embryonic development and thus share a very early developmental origin with skeletal muscle, bone, white adipocytes, and connective tissues (Wang et al., 2014b, Atit et al., 2006, Seale et al., 2008, Lepper and Fan, 2010, Sanchez-Gurmaches et al., 2012). However, the pathways that specify the brown adipocyte developmental lineage is not fully clear. In accordance with a mesodermal origin, a population of cells within the central dermomyotome that is labelled at E9.5 by expression of the homeobox transcription factor Engrailed 1 (En1) gives rise to iBAT, dermis, and epaxial muscles (Figure 2A, ,44 and Table 1) (Atit et al., 2006). However, these E9.5 En1+ progenitors do not appear to give rise to sBAT, or any of the major white fat depots (Atit et al., 2006)(Atit personal communication) suggesting that some brown and white adipocyte origins may differ, and that not all brown adipocytes share a common origin (see below). This concept of adipocyte heterogeneity within and between depots, as we will discuss, is now a central tenet of adipocyte biology.

Model of the heterogeneity and complexity in brown and brite/beige adipocyte development.
Several multi-potent cell populations that are mainly mesodermal and express specific transcription factors (e.g. En1, Myf5, Pax3, Prx1) appear to give rise heterogeneously to thermogenic adipocytes in different depots. Note that there is overlap shared with some markers but not with others. For example, Pax3 and Myf5 together may mark a pool of early precursors that give rise to iBAT, but only Pax3 marks a precursor pool that gives rise to some visceral pgWAT adipocytes (discussed in text). The significance of this heterogeneity is not understood. Additionally, there are several populations of brown and brite/beige adipocytes for which potential lineage markers remain unidentified. Also note that the brown and brite/beige adipocytes shown in this figure are depicted in their active state (i.e. upon β-adrenergic stimulation), but in vivo brown and brite/beige adipocytes are not necessarily present at the same time, such as in mild cold conditions (see Figure 2).
Table 1.
Depot-specific developmental origins of BAT
| Tissue Type | Anatomical Locations (Human) | Anatomical Locations (Mouse) | Developmental Origins (Lineage Tracing Study) |
|---|---|---|---|
| Brown adipocyte | Supraclavicular/Paravertebral Perivasicular: (aorta, artery) Periviscus: (heart, lung bronchia) Solid organs: kidney/spleen hilum pancreas, liver | Interscapular | En1+, Myf5+, Pax7+, Pax3+, Prx1− |
| Subscapular | En1−, Myf5+, Pax7+, Pax3+, Prx1− | ||
| Cervical | En1?, Myf5+/−, Pax7+, Pax3+/−, Prx1− | ||
| Peri-renal | En1?, Myf5−, Pax7+, Pax3+/−, Prx1− | ||
| Peri-aortic | En1?, Myf5−, Pax7+, Pax3−, Prx1+/− | ||
| Brite/beige adipocyte | Supraclavicular/Subcutaneous | Posterior-Subcutaneous (Inguinal) | En1?, Myf5−, Pax3−, Prx1+ |
The model tilted toward brown fat and skeletal muscle sharing a common developmental origin with the finding that brown adipocytes in the iBAT and the skeletal muscles, but not certain populations of WAT, share a common cellular origin in the dermomyotome defined by the expression of Myf5-Cre (Seale et al., 2008). Using the constitutively expressing Myf5-Cre knock-in driver with a cytoplasmic reporter, this study found that Cre recombinase activity labels mature brown adipocytes in the iBAT in addition to skeletal muscles (Seale et al., 2008). Myf5 is a classic myogenic determination factor from the basic helix loop helix (bHLH) family, and thus the labeling of brown adipocytes with Myf5-Cre (Tallquist et al., 2000) was predicated to explain the metabolic similarities between brown fat and skeletal muscle with respect to high oxygen consumption and fuel usage, and conversely the metabolic difference between BAT and the less metabolically active and energy storing WAT depots (Harms and Seale, 2013). Notably, at the time most studies used mice that were mildly cold stressed in which the BAT is hyperactive, rather than mice living at thermoneutrality, when brown adipocytes are more similar morphologically and metabolically to white adipocytes. Nevertheless, in support of this model, an inducible Cre driver under control of the Pax7 promoter (the PAX transcription factor family member 7 collaborates with Myf5 and other myogenic factors during skeletal myogenesis) showed that Pax7+ progenitors that arise between E9.5 and E10.5 (but not later in development) also give rise to interscapular brown adipocytes (Lepper and Fan, 2010). This also suggested an early divergence between BAT and muscle lineages.
While the Myf5-lineage model of BAT specification was elegant in its simplicity, studies challenging its uniformity soon after revealed that the brown adipocyte developmental landscape is more complicated. Similar fate mapping experiments using the same Myf5-Cre driver, but more broadly examining brown and white fat depots, and using the mTmG reporter, showed that many white adipocytes are also Myf5-Cre lineage positive, and unexpectedly that many brown adipocytes are Myf5-Cre lineage negative (Sanchez-Gurmaches and Guertin, 2014). For example, Myf5-Cre labelled precursors appear to give rise to nearly all brown adipocytes in iBAT and sBAT depots, but only about half of the brown adipocytes in the cervical BAT, and none of the brown adipocytes in prBAT or paBAT. Moreover, Myf5-Cre positive adipocytes populate the asWAT and rWAT depots (Figure 2A), indicating that Myf5-Cre neither uniformly nor specifically labels brown adipocytes. Other studies have replicated these findings confirming the heterogeneous labeling of adipocytes with Myf5-Cre (Sanchez-Gurmaches and Guertin, 2014, Sanchez-Gurmaches et al., 2012, Shan et al., 2013, Wang et al., 2014b)
Interestingly, lineage tracing using a Pax3-Cre driver, (Pax3 is another myogenic Pax family transcription factor that expresses just prior to Myf5) labels similar populations of cells with a few key differences. Notably, Pax3-Cre cells give rise to most of the brown adipocyte in iBAT, sBAT, cBAT and prBAT, but none of the brown adipocytes in the paBAT (Sanchez-Gurmaches and Guertin, 2014, Liu et al., 2013), and also to nearly 50% of the white adipocytes in the large visceral pgWAT depot. For comparison, MyoD-Cre (another classic myogenic transcription factor) does not label any brown or white adipocytes, but importantly does label skeletal muscles (Sanchez-Gurmaches and Guertin, 2014).
Thus, there may be specificity within skeletal muscle lineages in which some precursors (i.e. Pax3/Myf5/Pax7positive) can also become adipocytes while others (i.e. MyoDpositive) cannot, or rather that some adipocyte and muscle precursors can independently express Pax3/Myf5/Pax7-Cre (see discussion above on the challenges of lineage tracing studies)(Sanchez-Gurmaches and Guertin, 2014, Haldar et al., 2008, Gensch et al., 2008).
The most interesting possibility is that there is a temporal or spatial separation between certain lineages and understanding this may help in understanding the commitment phase to brown adipocytes. Regardless, these studies conclusively revealed an unanticipated heterogeneity in both brown and white adipocyte development that suggests brown adipocytes residing in different depots could have different embryonic origins.
The developmental heterogeneity observed between brown adipocyte lineages is not likely due to low efficiency or specificity of the Cre-drivers because independent experiments with Myf5-Cre, Pax3-Cre and Pax7-CreER lines are remarkably similar (Lepper and Fan, 2010, Sanchez-Gurmaches and Guertin, 2014, Liu et al., 2013, Sanchez-Gurmaches et al., 2012, Seale et al., 2008, Shan et al., 2013, Wang et al., 2014b).
Moreover, heterogenous Myf5 labeling is also observed in skeletal muscle lineages in which Myf5 only labels around 50% of the satellite cells in the limb muscles but around 80% in epaxial muscles (Haldar et al., 2008, Gensch et al., 2008). An unanswered question is whether developmentally distinct brown (or white) adipocytes differ only in their anatomical location, or whether they have unique functions (e.g. metabolic efficiency, BATokine production, exosome secretion, etc.) that might be specific by their developmental origins.
Answering these questions will require an improved ability to isolate and study single brown adipocytes, a deeper understanding of the regulatory mechanisms of BAT development, and markers that label the unidentified (Myf5-Cre;Pax3-Crenegative) brown adipocyte lineages.
REFERENCE LINK: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7330484/
More information: Becher, T., Palanisamy, S., Kramer, D.J. et al. Brown adipose tissue is associated with cardiometabolic health. Nat Med (2021). doi.org/10.1038/s41591-020-1126-7



{kind=link}