











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")


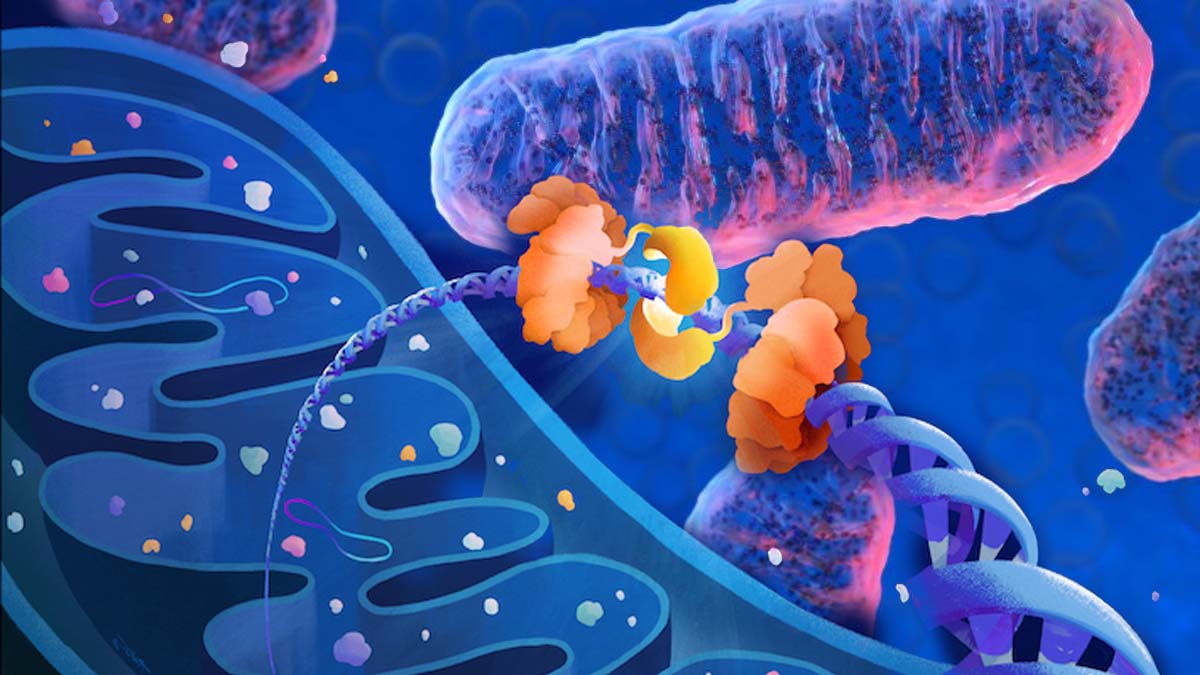
One of the most vexing aspects of the COVID-19 pandemic is doctors’ inability to predict which newly hospitalized patients will go on to develop severe disease, including complications that require the insertion of a breathing tube, kidney dialysis or other intensive care.
Knowledge of a patient’s age and underlying medical conditions can help predict such outcomes, but there are still surprises when younger, seemingly healthier patients suffer severe complications that can lead to death.
Now, scientists at Washington University School of Medicine in St. Louis have shown that a relatively simple and rapid blood test can predict—within a day of a hospital admission – which patients with COVID-19 are at highest risk of severe complications or death.
The study, published Jan. 14 in JCI Insight, involved nearly 100 patients newly admitted to the hospital with COVID-19.
The blood test measures levels of mitochondrial DNA, a unique type of DNA molecule that normally resides inside the energy factories of cells.
Mitochondrial DNA spilling out of cells and into the bloodstream is a sign that a particular type of violent cell death is taking place in the body.
“Doctors need better tools to evaluate the status of COVID-19 patients as early as possible because many of the treatments—such as monoclonal antibodies—are in short supply, and we know that some patients will get better without intensive treatments,” said co-senior author Andrew E. Gelman, Ph.D., the Jacqueline G. and William E. Maritz Endowed Chair in Immunology and Oncology in the Department of Surgery.
“There’s so much we still don’t understand about this disease,” he added. “In particular, we need to understand why some patients, irrespective of their ages or underlying health in some cases, go into this hyperinflammatory death spiral. Our study suggests that tissue damage may be one cause of this spiral, since the mitochondrial DNA that is released is itself an inflammatory molecule.”
The researchers said the test could serve as a way to predict disease severity as well as a tool to better design clinical trials, identifying patients who might, for example, benefit from specific investigational treatments.
They also said they would like to evaluate whether the test could serve as a way to monitor the effectiveness of new therapies. Presumably, effective treatments would lower mitochondrial DNA levels.
“We will need larger trials to verify what we found in this study, but if we could determine in the first 24 hours of admission whether a patient is likely to need dialysis or intubation or medication to keep their blood pressure from dropping too low, that would change how we triage the patient, and it might change how we manage them much earlier in the disease course,” said co-senior author Hrishikesh S. Kulkarni, MD, an assistant professor of medicine.
The researchers, including co-first authors Davide Scozzi, MD, Ph.D., a staff scientist, and Marlene Cano, Ph.D., a postdoctoral research scholar, evaluated 97 patients with COVID-19 at Barnes-Jewish Hospital, measuring their mitochondrial DNA levels on the first day of their hospital stays. They found that mitochondrial DNA levels were much higher in patients who eventually were admitted to the ICU, intubated or died. The researchers found this association held independently of a patient’s age, sex and underlying health conditions.
On average, mitochondrial DNA levels were about tenfold higher in patients with COVID-19 who developed severe lung dysfunction or eventually died.
Those with elevated levels were almost six times more likely to be intubated, three times more likely to be admitted to the ICU and almost twice as likely to die compared with those with lower levels.
Further, the test predicted outcomes as well as or better than existing markers of inflammation currently measured in patients hospitalized with COVID-19. Most other markers of inflammation measured in patients with COVID-19, including those still under investigation, are general markers of systemic inflammation, rather than inflammation specific to cell death, according to the researchers.
“Viruses can cause a type of tissue damage called necrosis that is a violent, inflammatory response to the infection,” Gelman said. “The cell breaks open, releasing the contents, including mitochondrial DNA, which itself drives inflammation.
In COVID-19 patients, there has been anecdotal evidence of this type of cell and tissue damage in the lung, heart and kidney. We think it’s possible that measures of mitochondrial DNA in the blood may be an early sign of this type of cell death in vital organs.”
The researchers also emphasized that the test is quick and straightforward to perform in most hospital settings because it uses the same machinery that processes the standard PCR test for COVID-19. The method they developed allows mitochondrial DNA levels to be quantified directly in the blood. Without requiring intermediate steps to extract the DNA from the blood, the technique returned results in less than an hour.
Before they can apply for approval from the Food and Drug Administration (FDA), the scientists will need to verify that the test is accurate in a larger multi-center trial. They have plans to expand the research to more sites.
The study utilized samples obtained from the School of Medicine’s COVID-19 biorepository, which was developed by co-authors Jane O’Halloran, MD, Ph.D., an assistant professor of medicine; Charles Goss, Ph.D., an instructor in biostatistics; and Phillip Mudd, MD, Ph.D., an assistant professor of emergency medicine.
Mitochondrial function in health
Mitochondria is a vital organelle present in all cells except mature erythrocytes, with key cellular functions involving oxidative phosphorylation, ATP synthesis, intracellular calcium regulation, thermoregulation, regulation of ROS, lipid metabolism, iron metabolism, autophagy, apoptosis, and regulation of certain hormones such as cortisol, estrogen, and triiodothyronine [5–7].
Elderly patients with high proportion of senescent tissue and immune cells with suboptimal mitochondria may be especially susceptible and unable to keep up with the sudden hypermetabolic demands associated with sepsis [8]. Further mitochondria also have a role and are associated with cellular senescence, chronic inflammation (inflammaging), decline in innate immunity, and the age-dependent decline in stem cell activity [9] (Fig. 1).

Covid 19 virus and possible mitochondrial dynamics, complex interplay in immunometabolism. Dysfunctional mitochondrial oxidative phosphorylation leads to accumulation of reactive oxygen species (ROS). There is activation of aerobic glycolysis, pentose phosphate pathway, and lactic acid formation, further increasing the ROS. This leads to mitochondrial membrane damage and release of DAMP factors. Mitochondria may also contribute to the formation of double-membrane vesicle which is critical in corona virus life cycle. Innate immunity is activated with downstream hyper-inflammatory cascade. HIF-α/Sirtuin pathways are also activated by cellular hypoxia and ROS and activate aerobic glycolysis and innate immune system. All these factors lead to multiple organ dysfunction and increase sepsis-related mortality. TLR Toll-like receptor is a PRR, RIG 1 retinoic acid inducible is a cytosolic PRR, IL interleukins, TNF tumor necrosis factor, IFN interferons, NLRP3 NOD like receptor is a PRR
As people age, their cells accumulate replication errors and mutations in the mitochondrial DNA (mtDNA) [8]. Oxidative damage to the cells by the reactive oxygen species (ROS) such as hydrogen peroxide (H2O2), superoxides, and hydroxyl radicals also accumulate with aging. T
he mitochondrial respiratory chain is the source of these ROS and they also have the mechanism for elimination with enzymes that have scavenger functions, for example: superoxide dismutase.
Mitochondrial dysfunction is also associated with toxic exposures and chronic anoxic conditions [7]. With a higher proportion of senescent cells in the elderly, deficiencies in the respiratory chain functions may develop eventually and may be associated with a decline in mitochondrial respiratory efficiency [9].
Mitochondrial dysfunction in sepsis (Fig. 1)
Sepsis is an abnormal pathological state and consists of an initial acute hyper-inflammatory phase, to fight the infection followed by an immune-tolerant, hibernating phase to aid with recovery. Different metabolic processes and reprogramming unique to each phase have been described and play a role in the different, varied clinical manifestations in the two phases. Hyper-inflammatory phase is characterized by increased aerobic glycolysis (Warburg effect).
This phase occurs as the host tissues reacts, with the aim of ramping up energy production, activating the innate immune cells to increase its pathogen killing capacity and control the spread of the infection and defend against the invading microbes [5, 10–12]. The late hypo-inflammatory, immune-tolerant phase is characterized by increased fatty acid oxidation in immune cells [10, 11].
A recent analysis of differential gene expression in the SARS-CoV-2 infected lung cell lines demonstrates upregulation of genes involved in mitochondrial cytokine/inflammatory signaling and downregulation in the mitochondrial organization, respiration, and autophagy genes [13]. These findings provide evidence that mitochondrial disruption hinders an effective immune response, increases inflammation, and severity in the Covid-19-related sepsis.
Metabolic and immune consequences, and release of DAMP (Fig. 1)
A key feature of early sepsis is hypoxia. At the cellular level, hypoxia correlates with a switch to aerobic (glycolytic) respiration with alteration of normal oxidative phosphorylation and mitochondrial function due to reduced activity of complex I–IV and complex V of electron transport chain. There is increased oxygen consumption, elevated ATP production, and hyperglycemia. In addition, due to increased cell turnover, there is increased requirement for nucleotides and increase in pentose phosphate pathway [4, 5, 10, 12]. These altered metabolic pathways result in production of excessive cytoplasmic and mitochondrial ROS. Failure to clear the ROS by senescent mitochondria may impair and damage mitochondrial structure, lipid membranes, biogenesis units, and alter the intact mitochondrial DNA [5, 14]. Other mitochondrial factors exacerbating and causing elevated ROS include altered lipid peroxidation, glutathione (GSH) depletion, and altered iron metabolism [15]. Loss of mitochondrial membrane integrity may release mtDNA in the systemic circulation. These circulating mtDNA fragments are also known as danger-associated molecular patterns (DAMP) and are similar to the pathogen associated molecular patterns (PAMP) which are released by microbes and identified by pathogen recognition receptors (PRR) on the immune cells. Nakahira et al. in a prospective study of 200 patients were first to demonstrate the association of elevated cell-free, plasma mtDNA levels with increased ICU-related mortality. They suggested mtDNA as a DAMP capable of propagating the inflammatory response seen in sepsis and other inflammatory conditions and a predictive biomarker for ICU-related mortality [16]. Other DAMP proteins associated with mitochondrial derangement include mtROS, mt formyl peptides, Cytochrome C, cardiolipin, carbamoyl phosphate synthetase, heat shock proteins (HSP), and high-mobility group box proteins-1 (HMBG-1) [5, 6, 17, 18] (Fig. 1).
These endogenously derived DAMP molecules from senescent aged, disrupted mitochondria can activate the innate immune system similar to the PAMP molecules obtained from virus particles. The activation of the PRR (family of Toll-like receptors) results in the activation of immune cells consisting of neutrophils, macrophages, dendritic cells, and natural killer cells. Activated innate immunity initiates the transcription of genes which lead to the production of signal molecules such as prostaglandins, pro-inflammatory cytokines, chemokines, and interleukins, and also activates the complement system, thus evoking the inflammatory cascade [16, 19].
DAMP have also shown to activate the (Nuclear factor kappa-light chain enhancer of B cell) NF-Kβ pathway, activation of inflammasommes, and (stimulator of interferon genes) STING pathways. These mechanisms exacerbate and compound tissue dysfunction involving cardiovascular, immune, metabolic, endocrine, coagulation, and thermoregulatory pathways, and are responsible for the systemic effects and multi-organ failure [5, 16, 19–21]. Previous reports from septic patients and septic animal models indicate decrease in intact mtDNA and an increase in circulating free mtDNA fragments [16, 18].
Role of HIF-α/Sirtuins in sepsis (Fig. 1)
Recent investigations have highlighted mechanisms in acute inflammation and sepsis cascade by HIF-α (hypoxia-inducible factor-Iα)–Sirtuins (silent mating type information regulator) signaling pathway [10, 11, 22]. Tissue hypoxia is a central feature of sepsis. There is upregulation of HIF-α signaling in tissue hypoxia. These signaling molecules and transcription factors are involved in both the hyper-inflammatory phase (elevated HIF-α/low Sirtuins) and the second immune-tolerant, recovery phase of sepsis (elevated Sirtuins/low HIF-α).
Sirtuins (types 1–7) are a highly conserved family of proteins known for their anti-inflammatory and anti-oxidant properties. These proteins belong to the (nicotinamide adenine dinucleotide) NAD+ dependent class III histone deacetylase (HDAC) family of enzymes and different types are present in different compartments of cells such as cytoplasm, mitochondria, and nucleus.
Sirtuins are (NAD+) energy sensors of a cell and are considered as guardians of homeostasis. Sirtuins type 3, 4, 5 are localized to the mitochondria [10, 11, 22] (Fig. 1).
NAD+ levels decline with aging and are further reduced in chronically stressed cells such as patients with diabetes, hypertension, smokers, and obesity thereby reducing the activity of Sirtuin.
These groups of patients have the highest risk for severe sepsis and mortality as currently observed in the Covid-19 pandemic [23]. Recent experiments on SARS-CoV-2 infected murine, human lung cell lines, and autopsy specimens suggest upregulation of poly (ADP) ribose polymerase (PARP) family of genes which are involved in the consumption of NAD+.
Overexpression of these PARP genes consume and depress the levels of NAD+ and are overexpressed in these cells [24]. Reduced activity and depletion of NAD+ further attenuate protective function of Sirtuin [23, 24].
Hypoxic cells upregulate the transcription factor, HIFα [12, 20]. In general, stabilization of HIFα and an increase in its hyper-inflammatory activity occur with loss or dysfunctional protective action of the cellular Sirtuins. This has been demonstrated in the hyper-inflammatory stage of animal models of sepsis [10–12, 22].
During this initial hyper-inflammatory phase with associated tissue hypoxia, the PRR (Toll-like receptors) upregulate a number of proliferative pathways such as NF-ĸB, hyper activation of NLRP3 inflammasommes, mitogen-activated protein kinase (MAPK), and mammalian target of rapamycin (mTOR) that trigger the cytokine storm [23–25].
Along with increased proliferative activity and increased cell turn over, the accompanying aerobic glycolytic pathway and dysfunctional oxidative phosphorylation release ROS which results in downregulation of Sirtuins. Low Sirtuin activity itself promotes elevated ROS due to inefficient clearance. Elevated ROS also downregulates prolyl hydroxylase (PHD).
These enzymes inactivate HIF-α. Taken together, these factors stabilize HIF-α and prevent it from undergoing ubiquitination by E3 ligases (von Hippel Lindau factor, VHL) for proteasomal degradation. Stabilized HIF-α translocates to the nucleus and activates key genes upregulating glycolysis and production of pro-inflammatory cytokines and maintains the vicious cycle of a hyper-inflammatory state [10–12, 22]. Key HIF-α targets include glucose transporter 1 (Glut1), hexokinase (HK), lactic dehydrogenase (LDH), pyruvate dehydrogenase kinase (PDHK), and cyclo-oxygenase (COX-2). These enzymes shift metabolism away from oxidative phosphorylation and promoting glycolytic signature and further fueling anabolic pathways [10–12, 22] (Fig. 1).
During the second phase of sepsis, a hypo-metabolic state will ensue with decreased oxygen consumption, resumption of mitochondrial respiration, and ATP production. Majority of the mortality associated with sepsis occurs during this phase. This hypo-metabolic state allows cells to enter an anti-inflammatory hibernating state allowing for slow recovery of cellular function. However, this cyto-protective, immunosuppressive state may also be detrimental to clear infections [10].
Moreover, secondary nosocomial infections may occur during this phase. For example: clostridium difficile infections, and secondary bacterial and candida infections. Often these infections are multidrug resistant and increase sepsis-related morality in the setting of the hypo-inflammatory phase [10].
Biochemically, this phase is associated with reduced activity of HIF-α and upregulation of Sirtuin activity. Sirtuins with their deacetylase function inactivate NF-ĸB, HIF-α, high-mobility group box proteins-1 (HMBG-1), and other DAMP molecules, thus decreasing the overall cytokine load [23–25]. Sirtuins also coordinate switch from glucose to fatty acid oxidation during this phase.
The molecular mechanisms for this remain unclear [10, 11, 14, 22, 25]. With cell recovery, mitochondrial biogenesis is initiated and there are upregulation of markers of mitochondrial biogenesis such as PAR gamma coactivator-1α (PGC-1α), Transcription factor A for mitochondria (TFAM), and nuclear respiratory factors (NRF-1) [9, 15, 23].
Mitochondria localization may be necessary for SARS-CoV-2 replication
SARS-COV-2 virus uses ACE2 receptor for cell entry and TMPRSS2 for spike protein (S protein) priming [26]. Variations of ACE 2 receptors in different ethnic populations may play a role in the virulence and transmissibility of this virus [27].
SARS-CoV-2 protein and human–protein interactions that connect multiple complex biological processes such as replication, protein trafficking, and ubiquitination have been observed. As a group SARS-CoV viruses produce accessory proteins called open reading frames (ORF) which interact with mitochondrial outer membrane receptors. One particular interaction involves ORF-9 interaction with mitochondrial antiviral signaling systems (MAVS, TOMM.70) [28].
MAVS is a mitochondrial import receptor and also functions as cytoplasmic viral recognition receptor. ORF has shown to suppress MAVS activity, thus limiting the initial host cell, innate immune, interferon, and antiviral response [28–30]. Similar other SARS-CoV-2 virus non-structural protein (NSP4, NSP5, NSP7, and NSP8) interaction with human mitochondrial membranes and matrix functions has been demonstrated.
These viral proteins have been demonstrated to associate with RNA processing, electron transport and mitochondrial signaling, and trafficking proteins. Thus, viral–mitochondrial interactions may disrupt both the membrane integrity and functional aspects of the mitochondria [28].
Once inside the cytoplasm, the SARS-CoV-2 virus has to replicate from a single-stranded RNA (ss RNA) through an intermediate double-stranded RNA (ds RNA). This predisposes the virus to the antagonism through their recognition by Toll-like receptors, RIG-1 receptors, and mitochondrial antiviral signaling systems (MAVS) and, thus, activation of innate immunity as described earlier. SARS-CoV-2 virus evades this detection by forming double-membrane vesicles (DMV) around its dsRNA, thus shielding it from detection. Viruses need intracellular organelles such as mitochondria and endoplasmic reticulum for their replication and dissemination [31, 32].
Based on computational machine learning models, it has recently been demonstrated by Wu et al. that the SARS-CoV-2 RNA genome and all sub-genomic RNAs are enriched in the host mitochondrial matrix and nucleolus [33]. Open reading frame (ORF) from corona viral genome has been identified in the mitochondria.
Based on these models, they predicted that mitochondrial residency may be required to the formation of double-membrane vesicles which is a critical in corona virus being able to replicate un-abated by evading cellular defenses. In other words, the virus simply may hijack the mitochondria and use its machinery for its own replication and sustenance [33].
In doing so, it may damage the mt DNA and this may cause leakage of the mt DNA into the cytoplasm with then acts as a trigger for activation of innate immunity [34]. Previously, an association between high mitochondrial viral RNA and decrease in mitochondrial functional integrity has been demonstrated with HIV RNA virus [35]. This hypothesis needs further validation with SARS-CoV-2.
These findings suggest that mitochondrial involvement by the SARS-CoV-2 RNA virus may actually assist with virulence and transmissibility. Mechanisms by which the virus gains access into the mitochondria and cause dysfunction remain to be investigated.

Venn diagram: depicting the risk for severe sepsis and increased mortality in Covid-19 pandemic. A combination of: (a) elderly patients with comorbidities living in close knit communities and densely populated areas, (b) associated mitochondrial dysfunction, and risk factors such as diabetes, hypertension, lung diseases, and activation of innate immunity, and (c) an increased viral load; all together set a perfect storm for increased viral shedding and transmissibility with risk for severe sepsis and high mortality
Several viral and host-related biomarkers are emerging to be critical in determining the severity of infection in Covid-19 sepsis. The major question remains whether these biomarkers perform consistently, can be tested and replicated uniformly, and can be validated by pooling of the currently available data.
As the current SARS-CoV-2 virus continues to spread in the USA and worldwide, high quantitative viral load emerges as a critical factor in epidemiology, and also to predict the clinical course in Covid-19 infection [36–40]. Key observations with current Covid-19 pandemic demonstrate a high mortality in elderly patients with sepsis. Increased quantitative viral load represents a high level of viral replication and is a surrogate marker for infectivity.
This association may be related to its interaction and mitochondria dysfunction in the elderly. Previously, studies have attempted to ascertain an association between viral loads and increased severity with prolonged hospitalization on patients infected with influenza, rhino, and other respiratory virus [36, 37, 40]. Real-time PCR assays (rRT-PCR) to detect respiratory viruses such as group 2 coronaviruses (HCoV-OC43 and HCoV-HKU1), and SARS associated coronavirus have demonstrated that a high viral load measured on nasopharyngeal samples at the point of hospitalization was associated with longer length of stay in adult patients [36, 40].
Similar to recent literature on SARS-COV-2 virus, Covid-19 pandemic has suggested that higher viral load values from nasopharyngeal swabs remain a significant factor and are associated with prolonged virus shedding period and severe clinical outcomes [38, 39]. Elevated viral loads may be noted in both asymptomatic and symptomatic patients. This remains a cause of concern, as asymptomatic carriers may be silent spreaders in the population. Viral load may be a useful marker for both detecting super spreaders and also disease severity and prognosis in the symptomatic patients [38, 39].
Early determination of quantitative viral load also has other therapeutic benefits, such as early implementation of convalescent plasma, antiviral, and corticosteroids. Convalescent plasma from donors who recovered from Covid-19 infection could potentially improve the clinical outcomes by decreasing the severity in Covid-19 cases [41, 42].
A persistently elevated viral load may thus be associated with worse metabolic and mitochondrial dysfunction with a prolonged hyper-inflammatory phase as described earlier. This hypothesis may explain the clinical variation observed with Covid-19 patients worldwide, i.e., higher mortality in elderly, especially in a close knit population such as nursing homes suggestive of mitochondrial dysfunction associated with high viral load (Fig. 2). Further epidemiological studies and assays on Covid-19 viral loads are needed to confirm this hypothesis.
Other host-related biomarkers which are consistently observed and correlated in various populations across the world from the current pandemic are elevated levels of d-dimer, IL-6 concentration, and lymphopenia. These portend severity of the inflammation and correlate with poor prognosis and increased mortality [41, 43, 44].
reference link : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7410962/
More information: Scozzi D, Cano M, et al. Circulating mitochondrial DNA is an early indicator of severe illness and mortality from COVID-19. JCI Insight. Jan. 14, 2021. www.biorxiv.org/content/10.110 … /2020.07.30.227553v1



{kind=link}