











 Extract (6-MSITC) in Healthy Older Adults")
: An In-Depth Exploration into its Thermogenic Role and Social Significance")



The bacterial gut microbiome is strongly associated with Parkinson’s disease (PD), but no studies had previously investigated the role of fungi in the gut. In this novel study published in the Journal of Parkinson’s Disease, a team of investigators at the University of British Columbia examined whether the fungal constituents of the gut microbiome are associated with PD.
Their research indicated that gut fungi are not a contributing factor, thereby refuting the need for any potential anti-fungal treatments of the gut in PD patients.
“Several studies conducted since 2014 have characterized changes in the gut microbiome,” explained lead investigator Silke Appel-Cresswell, MD, Pacific Parkinson’s Research Centre and Djavad Mowafaghian Centre for Brain Health and Division of Neurology, Faculty of Medicine, University of British Columbia.
“Most existing studies, however, employ bacterial-specific sequencing. To date, a potential role for the fungal constituents of the gut microbiome, also known as the “mycobiome,” has remained unexplored.”
In order to investigate whether the fungal constituents of the gut microbiome are associated with PD researchers enrolled 95 PD patients and 57 controls from the Pacific Parkinson’s Research Centre (PPRC) at the University of British Columbia. Participants provided a single fecal sample and completed a two-hour study visit during which their PD symptoms were assessed.
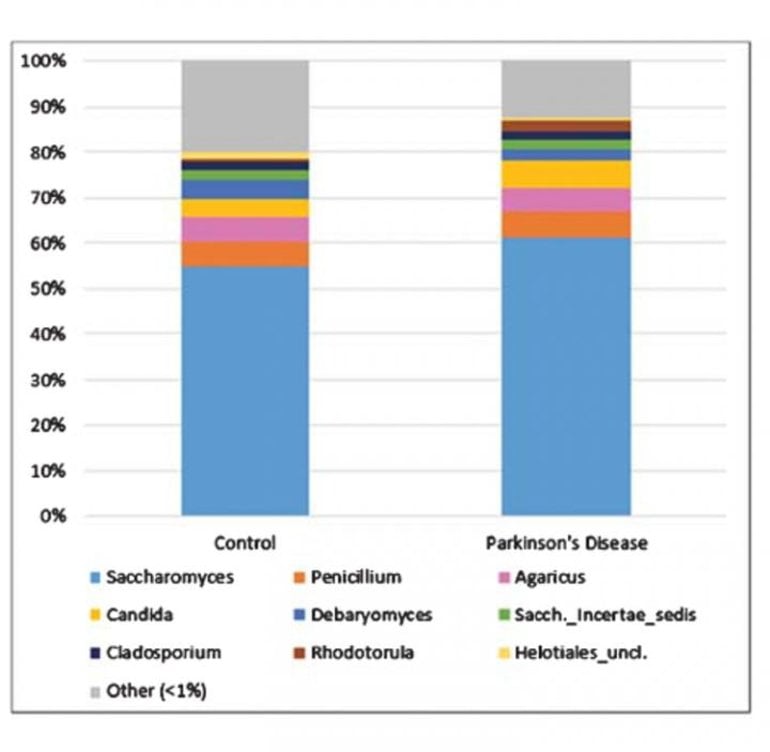
Analysis determined that the fungal microbiome in PD did not essentially differ from that of matched controls, and there were no strong associations between gut fungi and PD symptoms.
Fungi were very sparse among participants’ fecal microbiomes. After filtering, 106 of the 152 participants (64/95 PD and 42/57 control) remained for downstream compositional analysis; the remainder had virtually no detectable fungal genomic content. Most of the genera identified were environmental or dietary in origin.
Saccharomyces was by far the most dominant fungal genus detected. Although these investigations did not reveal any significant role for gut fungi in PD, interestingly, lower overall fungal abundance (relative to bacteria) in the PD gut were observed, which might reflect a less hospitable environment of the gut in PD.
This paper plays an important role by answering the call by the PD research community and funding organizations to publish negative results, crucial to avoid investing precious research funding into likely futile endeavors and providing a more balanced reflection of data in the field.
“The data are an important piece in the puzzle of understanding the overall role of the gut microbiome in PD,” continued Dr. Appel-Cresswell.
“The gut microbiome in PD continues to be an exciting field of research where we are just at the beginning of unraveling potential mechanisms. It will be important to publish negative results as well as positive findings along with detailed methods to have a realistic reflection of the data in the literature to accelerate discovery,” she concluded.
PD is a slowly progressive disorder that affects movement, muscle control, and balance. It is the second most common age-related neurodegenerative disorder affecting about 3% of the population by the age of 65 and up to 5% of individuals over 85 years of age. In recent years, more attention has been given to the gut as a key player in the initiation and progression of PD.
One of the most important challenges in modern medicine is the elucidation of the etiology of neurodegenerative diseases such as Alzheimer’s disease (AD), Parkinson’s disease (PD), amyotrophic lateral sclerosis (ALS) and multiple sclerosis (MS), each of which has remained obscure. PD is a progressive and debilitating disease that induces motor disorders, rigidity and dyskinesia through the functional loss of dopaminergic neurons in the substantia nigra 1.
PD affects over 6 million people worldwide 2-4, with the vast majority of patients diagnosed in their sixth decade of life. In addition to movement disorders, many patients show other clinical symptoms, including psycho emotional and cognitive deficits, dementia, sleep disturbances and olfactory and bladder dysfunction, among others 5-9.
Some of these symptoms can present years before diagnosis, prior to when motor signs are apparent. Progressive cognitive decline has been associated with gray matter atrophy, cortical thinning and considerable alterations to both gray and white matter 10. Moreover, global brain atrophy and modifications in several CNS regions are present in addition to the destruction of the substantia nigra, including the amygdala, thalamus and right caudate hippocampus and nucleus accumbens 11, 12.
In fact, PD represents a very heterogeneous disease and each patient differs regarding their symptoms and clinical progression and, accordingly, several subtypes of PD have been proposed 13. It has been suggested that the initiation of neurodegeneration in PD occurs in the olfactory bulb, which in some instances reduces its volume by about 50% 14-16. Thus far, treatments for PD are directed to alleviate clinical symptoms, for example, by maintaining the levels of dopamine in the striatum 17, 18. There are no treatments to stop or reverse the course of the disease.
An important neuropathological hallmark of PD is the abnormal deposits of synuclein in some neurons of several brain regions 4, 14. These aggregates were described as Lewy bodies over a century ago, and initially appear in cholinergic and monoaminergic brainstem neurons in the olfactory system, but are also found in limbic and neocortical brain regions as the disease progresses 19.
Other cellular functions are also affected, such as mitochondrial activity, oxidative stress, calcium homeostasis and axonal transport. Another important pathological feature of PD is systemic inflammation 20-22. Neuroinflammation is likely an essential contributor to PD pathogenesis of the CNS, and the elevation in the levels of a number of cytokines and interleukins in peripheral blood of patients point to the stimulation of the immune system 23.
The presence of inflammation has prompted the suggestion that PD is caused by viral or bacterial infection. Thus, it has been speculated that influenza virus or Actinobacteria may be involved in the etiology of PD 24-27. It has been suggested that molecular mimicry between HSV1 and α-synuclein could foster the progression of PD 28, 29.
Also, the possibility that some toxins or fungi can provoke PD has been suggested 30-34. More recently, the possibility that Malassezia might be contributing to PD has been hypothesized 35. However, the direct demonstration that these pathogens are present in brain tissue of PD patients was not provided.
A possible link between the gut microbiota and neurodegeneration has also been developed in recent years 36-38. Thus, the gut microbiota can influence CNS functioning, microglia activation and, in some instances, may lead to the synthesis of metabolites that provoke the pathological communication of gut microbes with microglia in the CNS and induce degeneration by the synthesis of toxic molecules.
In the context of PD, the gut microbiota differs from control individuals and can influence α-synuclein aggregation 37, 39, 40. Accordingly, the gut microbiota in each PD patient may contribute to disease pathogenesis 36, 41. With respect to fungal infection, a connection between the mycobiota and neurodegeneration has been recently reviewed 41.
Similarities between PD and other neurodegenerative diseases have been noted both in clinical symptoms and in cytoplasmic protein aggregates 42-44. Specifically, PD and AD share cognitive decline, the formation of amyloid plaques and phosphorylated tau and α-synuclein aggregates in the cytoplasm of neurons 45, 46.
Hence, it is possible that both diseases may share a similar pathological agent that affects different CNS regions. For some years, we have advanced the concept that these diseases might be caused by polymicrobial infections 47-52, involving a variety of fungal and bacterial species that can progressively colonize the central nervous system (CNS) 47, 51, 53, 54. Some of these microbial cells can be found inside neurons and are located intranuclearly.
In addition, corpora amylacea (CA) from both AD or PD patients contain fungal components, supporting the concept that this infection is located at the CNS, and that these bodies have formed during months or even years, trapping microbial antigens in the brain 55. Against this background, the aim of the present study was to investigate the presence of both fungi and bacteria in brain tissue from patients with PD.
Our results provide direct evidence of the existence of both fungi and bacteria with different morphologies in neural tissue from several CNS regions. The precise fungal and bacterial species found in each PD patient was determined using nested PCR and next-generation sequencing (NGS). Collectively, our present findings provide the first direct evidence of fungal and bacterial infection of PD brains.
Results
Direct visualization of fungal structures in the CNS of PD patients
Previous studies from our laboratory employed a battery of rabbit polyclonal antibodies to examine for fungal structures in human brain sections 47-49, 52. As these antibodies are polyclonal, they cross-react with a variety of fungal species, including the one used to raise the antibody. Accordingly, we can obtain direct support for the existence of mycotic structures (yeast-like and hypha) in the CNS, but we cannot make conclusions about the specific fungal species using this technique.
As a first attempt to explore the presence of fungi in PD CNS, we examined sections from various brain regions, as described in Materials and Methods. CNS sections were analyzed by double immunofluorescence using one of the following antifungal antibodies: anti-C. albicans, anti-C. glabrata, anti-S. racemosum, and anti-P. betae and a second antibody against the human protein α-tubulin.
In neurodegenerative diseases and in aged subjects, α-tubulin can be found not only in the cytoplasm, but also in some nuclei of brain cells 59. Notably, a variety of morphologies were revealed in the CNS of patient PD1 by immunohistochemistry and confocal microscopy (Figure (Figure1).1).
Several fungal structures (green) were clearly evident, such as yeast-like cells (see panels MD S. racemosum and PN C. glabrata), hyphal structures (panels MS P. betae, CB C. glabrata and CLN S. racemosum) and other rounded small bodies (panels MD C. glabrata, PN S. racemosum and CB C. glabrata). DAPI staining (blue) revealed the presence of nuclei in some of these fungal structures. Accordingly, the evidence for fungal infection is based on both the typical mycotic morphologies and their reactivity with anti-fungal antibodies.

Fungal structures in different CNS regions from one PD patient (PD1) analyzed by immunohistochemistry. Six CNS regions (medulla, MD; mesencephalon, MS; hypothalamus, HT; pons, PN; callosal body, CB; and caudate and lenticular nuclei, CLN) of PD1 were processed for immunohistochemistry as described in Materials and Methods. Parafin sections (5 µm) were immunostained with rabbit polyclonal antibodies against C. albicans, C. glabrata, S. racemosum and P. betae (green). Samples were then immunostained with a mouse monoclonal antibody against human α-tubulin (red). Finally, nuclei were stained with DAPI (blue). Scale bar: 5 µm.
To further assess the presence of fungi in these sections, we used two specific rabbit polyclonal antibodies raised against purified fungal enolase and β-tubulin, which do not cross-react with human tissue 56. The anti-enolase antibody (green) was combined with a mouse monoclonal anti-human α-tubulin antibody (red), whereas the anti-fungal β-tubulin antibody (green) was combined with a mouse monoclonal anti-human neurofilament antibody (red). Once again, typical mycotic morphologies could be revealed using these two monospecific antibodies (Figure S1).
To extend this analysis to other patients, three CNS sections (MC, MD and MS) from five PD patients (PD2-PD6) were analyzed by immunohistochemistry and confocal microscopy. Initially, these samples were immunostained with a rabbit polyclonal antibody for C. albicans (green) and a mouse monoclonal antibody for human α-tubulin (red).
In all cases, there was clear evidence for the presence of fungi in the three CNS regions (Figure (Figure2).2). Of interest, some forms that immunoreact with the anti-C. albicans antibodies appeared to be intracellular (see panels PD3 MS and PD4 MC). We then used an anti-P. betae antibody to examine these samples (Figure S2), finding morphologies similar to those observed with patient PD1 with the other antifungal antibodies, which is consistent with the occurrence of fungal infection in the CNS of PD patients. Comparison of these findings with fungi observed in control elderly persons using the same antibodies indicated that the burden of fungal infection was greater in PD as compared with controls 51.

Immunohistochemistry analysis of CNS regions from five PD patients (PD2-PD6). Three CNS regions (motor cortex, MC; medulla, MD; and mesencephalon, MS) from five PD patients (PD2-PD6) were inmunostained with a rabbit polyclonal antibody against C. albicans (green) and a mouse monoclonal antibody against human α-tubulin (red). Nuclei were stained with DAPI (blue). Scale bar: 5 µm.
Intracellular localization of fungi in neural cells
As mentioned, some of the fungal structures observed presented an intracellular location, and in some instances, appeared to be intranuclear. To further examine this possibility, we analyzed orthogonal projections of these specific cells. As shown in Figure Figure3,3, some yeast cells could be viewed inside the nucleus or close to it, and hyphal structures could also be detected surrounding the nuclei.
Both yeast and hyphae could be stained with DAPI (blue), indicating that they contain nuclei, which was well differentiated from the neural cell nucleus (Figure (Figure3).3). Finally, video images provided compelling evidence for the intracellular location of the fungal cells (Movie S1, S2, S3, S4 and S5, available online). Of note, the intracellular colonization of human cells by yeast requires that the cell is metabolically active 60-64. Therefore, our observations lend support to the concept that human neural cells were alive during the progression of fungal infection.

Orthogonal projections and 3D analyses of CNS sections from PD patients. Immunohistochemistry was carried out as described in Figure Figure1.1. Orthogonal projections (panels A-C) and different stacks from a 3D image (panels D-H) (see also Movies S1-S5) from different sections and PD patients. Panels A and D-F: PD1 and panels B, C and G,H: PD4. Panels A, D-F: CLN region; panels B,C and G: MC region and panel H: MD region. Panels A and D-F: samples were immunostained with an anti-P. betae antibody (green) and panels B,C and G,H: samples were immunostained with an anti-C. albicans antibody (green). Nuclei were stained with DAPI (blue). All scale bars: 5 µm.
Detection of chitin in PD CNS
The polysaccharide chitin is a typical component of the fungal cell wall and can be detected by means of specific antibodies 65, 66. Because the presence of chitin in human tissues provides persuasive evidence for mycotic cells, we examined for the existence of chitin in CNS sections from six PD patients (PD1-PD6) using a rabbit polyclonal anti-chitin antibody (green), and counterstained with a mouse monoclonal anti-human α-tubulin antibody (red).
Strikingly, a number of hyphae could be revealed using the anti-chitin antibody (Figure (Figure4).4). Consistent with the previous results, some hyphae seemed to be intracellular, whereas others were detected in intercellular spaces and, in some instances, DAPI staining (blue) of the nuclei of these hyphae was evident.
In addition to these typical mycotic structures, other morphologies and diffuse material were observed with the anti-chitin antibody (Figure S3). Importantly, some rounded fungal cells were found both intranuclearly and outside of the nucleus (Figure S3, panels PD1 MS, PD3 MC and MS). In conclusion, the evidence for cells containing chitin in the CNS of PD patients supports the existence of fungal infection. The various sizes and morphologies observed likely reflect the existence of different fungal species.

Hyphal structures in CNS sections of PD patients detected by an anti-chitin antibody. Several regions from different PD patients were processed for immunohistochemistry analysis as described in Materials and Methods using anti-chitin (green) and anti-human α-tubulin (red) antibodies. Nuclei appear in blue. Scale bar: 20 µm for panels A-D and 5 μm for panels E-J
As a control for this analysis we used CNS sections from four elderly controls. As shown in Figure S4, there was no sign of chitin immunopositivity in these sections, demonstrating that fungal colonization is not detectable in these subjects with the anti-chitin antibody.
reference link : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7053320/
Original Research: Closed access.
“The Gut Mycobiome in Parkinson’s Disease” by Appel-Cresswell et al. Journal of Parkinson’s Disease



{kind=link}